
Abstract
Background:
Globally, antimicrobial drug–resistant Escherichia coli is among the most common etiological agents of invasive disease in humans. In Europe, increasing proportions of infections due to third-generation cephalosporins and/or fluoroquinolone-resistant extraintestinal pathogenic E. coli (ExPEC) strains are reported. E. coli from poultry are those more closely linked to human E. coli, but lack of reliable data makes it difficult to assess the attributable risk of different food sources. In the present study, our objective was to investigate the antimicrobial resistance profile, phylogenetic background, and virulence factors of E. coli isolates from broiler chicken meat sold at retail in Palermo, Italy.
Materials and Methods:
Isolation of multidrug resistant (MDR) E. coli was performed during April–December 2013 on a total of 163 chicken meat samples. Susceptibility to a panel of nine antimicrobial agents was determined. PCR assays were carried out to detect extended-spectrum β-lactamase (ESBL), plasmid-mediated AmpC β-lactamase, and plasmid-mediated quinolone resistance (PMQR) genes, phylogenetic group, and ExPEC-associated traits. A single nucleotide polymorphism (SNP) PCR was done to detect E. coli sequence type (ST)131.
Results:
One hundred thirty-four isolates from 109 meat samples were MDR. B1 was the most prevalent phylogenetic group (47.8%), followed by groups D (25.4%), A (22.3%), and B2 (4.5%). ESBLs and AmpC β-lactamases were detected by PCR in 132 (98.5%) and 15 (11.2%) isolates. PMQR determinants were detected in 122 (91%) isolates. Twenty-two MDR isolates met the molecular definition of ExPEC. SNP-PCR results confirmed that four B2 isolates were ST131. Enterobacterial Repetitive Intergenic Consensus sequence–PCR analysis showed a large heterogeneity with 55 unique profiles and 31 clusters including 2–4 isolates.
Conclusions:
An alarmingly high prevalence of MDR E. coli from broiler chicken meat is evident in our geographic area. The ongoing use of antimicrobial drugs in livestock should be urgently restricted, particularly in the poultry sector.
Introduction
E
Antibiotics in livestock production are used for a variety of reasons, such as infectious disease treatment, prevention, control, and growth promotion/feed efficiency (Anderson, 2003; DANMAP, 2010). However, any use of antimicrobial drugs, mainly low-dose, prolonged courses among food animals, will select for antibiotic-resistant bacteria, which in turn will spread by direct contact or through food, water, and animal waste application to farm fields (Finley et al., 2013; Millman et al., 2013). Methods of livestock production differ in antibiotic use, influencing the prevalence of antibiotic-resistant bacteria on retail meats (Millman et al., 2013).
Extended-spectrum β-lactamase (ESBL) and AmpC-producing E. coli are associated with localized or systemic disease in humans (Reich et al., 2013). Resistance genes coding ESBLs are mostly plasmid associated and therefore can easily horizontally spread among E. coli strains. Additionally, although chromosomal AmpC genes exist in several enterobacteria including E. coli, plasmid-bound types can be also present (Nikaido, 2009). Emergence of resistance to fluoroquinolones that, because of their low toxicity and relatively broad-spectrum coverage, are widely employed tools for treating human infections, is of particular concern (Angulo et al., 2000; Livermore et al., 2002; Literak et al., 2013).
There are many reports in the literature describing the molecular characteristics of E. coli isolates from human clinical samples and animals, and several studies have examined the degree of molecular similarities between pathogenic isolates of either origin (Johnson et al., 2005a, 2007; Jakobsen et al., 2010; Kluytmans et al., 2013a). The role of E. coli isolates from foods of animal origin as a link between the livestock niche and humans is of special interest.
In this study, our objective was to investigate phylogenetic background, antimicrobial resistance profile, virulence factors, and their association in E. coli isolates recovered from retail broiler chicken meat on sale in Sicily, Italy.
Materials and Methods
Bacterial isolates
A total of 163 chicken broiler meat samples were bought from a range of retail grocery stores in Palermo, Italy, during April–September 2013. According to labeling, chicken had been conventionally raised in Italy. A 5-g portion of each specimen was added to 45 mL of modified trypticase soy broth, containing 20 mg/L of novobiocin (Sigma Chemical Co., St. Louis, MO). The enrichment cultures were incubated at 37°C for 18–24 h and then subcultured as a continuous lawn onto MacConkey agar plates. Antibiotic disks containing ceftazidime (30 μg), ciprofloxacin (5 μg), and cefoxitin (30 μg) were placed on each plate before incubation. One lactose-positive colony of each morphologically different type, growing into the antibiotic inhibition haloes, was picked up, subcultured, and presumptively identified as E. coli by indole, citrate, methyl red, and Voges-Proskauer tests. To confirm identification, polymerase chain reaction (PCR) amplifications of trpA gene, as a species-specific gene, for E. coli was performed (Clermont et al., 2009).
Antimicrobial susceptibility and ESBL/AmpC status
Susceptibility to 10 antimicrobial agents including amoxicillin–clavulanic acid (20–10 μg), ciprofloxacin (5 μg), cefotaxime (30 μg), cefoxitin (30 μg), ceftazidime (30 μg), cefepime (30 μg), gentamicin (10 μg), imipenem (10 μg), sulfamethoxazole–trimethoprim (25 μg), and tetracycline (30 μg) was determined by disk diffusion and interpreted according with the EUCAST (European Committee on Antimicrobial Susceptibility Testing) guidelines (
Phylogenetic grouping by multiplex PCR
Phylogenetic group was determined using a triplex PCR according with Clermont et al. (2000).
Multiplex PCR assay for ESBLs and AmpC detection
Three multiplex PCRs were used: (1) bla TEM/bla SHV/bla OXA-1-like PCR; (2) bla CTX-M PCR including groups 1, 2, and 9; and (3) a plasmid-mediated AmpC β-lactamase gene targeting PCR including six groups (i.e., ACC, FOX, MOX, DHA, CIT, and EBC) (Dallenne et al., 2010). As positive controls, we used previously characterized isolates from our collection carrying all types of tested β-lactamases.
Detection of plasmid-mediated quinolone resistance (PMQR) determinants
All isolates were investigated for the presence of qnrA, qnrB, qnrS, and aac(6′)-Ib genes according with Cavaco et al. (2008). As positive controls, previously characterized isolates carrying the PMQR gene sequences under study were used.
Virulence genotyping
All isolates belonging to phylogenetic groups B2 and D were investigated for the following eight ExPEC-related virulence genes by two quadruplex PCRs (Johnson et al., 2005b). Four strains were used as positive controls in the PCR screening: E. coli RS218 (kpsMT II, papA, papC, sfaS, hlyD), E. coli V27 (kpsMT II, papA, papC, iutA, focG), E. coli 2H16 (papC, iutA, afa, hlyD), and E. coli J96 (papA, focG). E. coli MG1655 was used as a negative control in the reactions. All the reference E. coli strains were kindly provided by James R. Johnson (VA Medical Center, Minneapolis, MN). E. coli isolates were defined as ExPEC if they tested positive for at least two of papA and/or papC, sfa/foc, afa/dra, kpsMT II, and iutA genes (Johnson et al., 2005b).
Detection of ST131
All group B2 isolates were screened by PCR for the ST131-associated single nucleotide polymorphisms (SNPs) in mdh and gyrB (Johnson et al., 2010). E. coli strains MVAST131, jj2663, and BUTI 1-2-1 (H17) were used as positive control strains.
ERIC-PCR analysis
Enterobacterial Repetitive Intergenic Consensus sequence polymerase chain reaction (ERIC-PCR) was performed on all isolates (Wilson et al., 2006). BioNumerics software, version 6.1, (Applied Maths, Sint-Martens-Laten, Belgium) was used for analyzing the ERIC-PCR fingerprints. The similarities in amplicon profiles were compared using a Dice coefficient at 1% tolerance and 0.5% optimization, and a dendrogram was constructed with the unweighted-pair group method with arithmetic mean clustering method with a cut-off of 80% similarity (McLellan et al., 2003).
Results
All 163 chicken meat samples under investigation grew E. coli and 109 (66.9%) yielded isolates resistant to ceftazidime, ciprofloxacin, or both. In particular, E. coli isolates resistant to ceftazidime were found in 95 samples of 109 (87.1%), to cefoxitin in 24 samples (22.0%), and to ciprofloxacin in 109 samples (100.0%), respectively. One hundred thirty-four E. coli were isolated from the 109 meat samples and further analyzed. Their resistance pattern can be summarized as follows: 92 (68.7%) were resistant to amoxicillin–clavulanic acid, 123 (91.7%) to cefotaxime, 123 (91.7%) to ceftazidime, 23 (17.1%) to cefepime, 24 (17.9%) to cefoxitin, 119 isolates (88.8%) to ciprofloxacin, 19 (14.1%) to gentamicin, 106 (79.1%) to sulfamethoxazole–trimethoprim, and 123 (91.8%) to tetracycline. None was resistant to imipenem (Table 1).
Tested by the disk-diffusion method.
AMC, amoxicillin–clavulanic acid; CAZ, ceftazidime; CTX, cefotaxime; FEP, cefepime; FOX, cefoxitin; GEN, gentamicin; CIP, ciprofloxacin; SXT, sulfamethoxazole–trimethoprim; TET, tetracycline.
All of the 134 isolates tested positive for ESBL production by DDST. A putative AmpC expression by the 3-dimensional test with cefoxitin disk was detected in 24 cefoxitin-resistant isolates. All of the 134 isolates met the criteria for the definition of MDR.
Phylogenetic group B1 was the most prevalent (64 isolates, 47.8%) followed by groups D (34 isolates, 25.4%), A (30 isolates, 22.3%), and B2 (six isolates, 4.5%). Table 1 describes frequency of resistance to the antibacterial drugs tested according to phylogroups.
ESBLs and AmpC β-lactamases were detected by PCR in 132 (98.5%) and 15 (11.2%) isolates, respectively, while PMQR determinants were detected in 122 (91%) of isolates. Moreover, all isolates testing positive for the PMQR genes carried also ESBL gene determinants. In particular, 90 isolates (67.2%) tested positive for SHV, 104 (77.6%) for TEM, and 65 (48.5%) for both TEM and SHV. None of the isolates was positive for the OXA-1-like gene. Twelve isolates (8.9%) were positive for CTX-M-1 group, 23 isolates (17.2%) for CTX-M-2 group and 10 isolates (7.5%) for CTX-M-9 group, respectively. Fifteen isolates (11.2%) carried AmpC genes which all proved to be CIT-like. Two isolates of D phylogroup were positive for SHV, TEM, CTX-M, and plasmidic AmpC β-lactamases. Among the PMQR determinants, only qnrA was detected in all fluoroquinolone-resistant isolates. The differential distribution of the ESBL, AmpC, and PMQR genes under study according with the phylogroup is shown in Figure 1. Phylogroup D isolates carried CTX-M more frequently than phylogroups A and B1 isolates (D, 16 of 34 isolates [47.0%] versus A, 9 of 30 isolates [30.0%] and B1, 15 of 64 [23.4%], respectively) and Amp-C (D, 11 of 34 isolates [32.3%] versus A, no isolate and B1, 3 of 64 [4.7%], respectively) genes. Two and one B2 isolates, respectively, tested positive for CTX-M and AmpC.
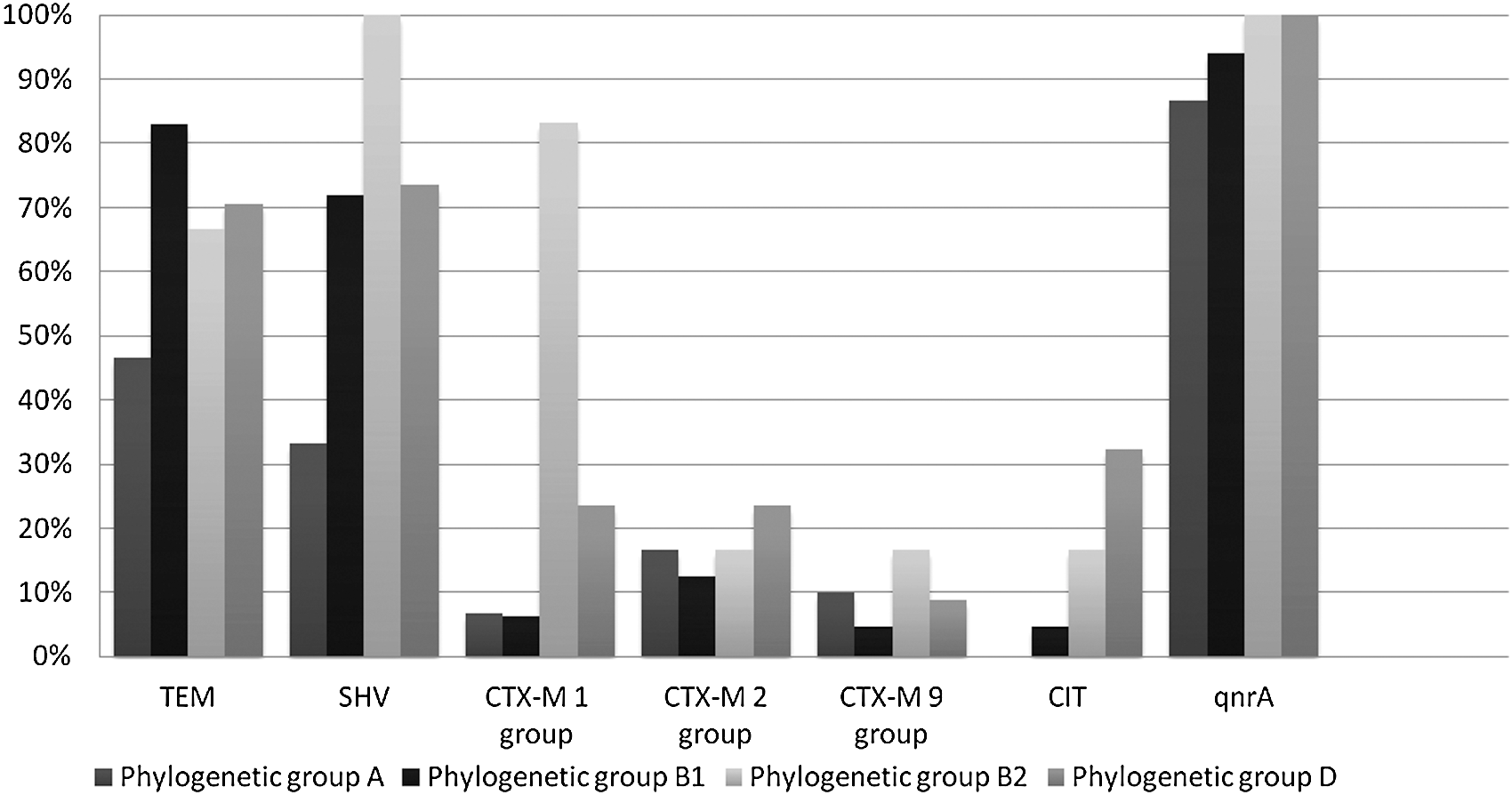
Distribution of the resistance genetic determinants under study by phylogroup.
Based on the molecular definition of ExPEC, 22 isolates of 40 belonging to phylogroups B2 and D were attributed with the status of ExPEC (all the B2 isolates and 16 of 34 [47.1%] D isolates). The most prevalent virulence factors were kpsMT II and iutA. SNP-PCR results confirmed that 4 isolates of phylogroup B2 were ST131. These four isolates were positive for both ESBL and PMQR determinants.
The ERIC-PCR analysis showed that 79 of the 134 E. coli isolates belonged to 31 different clusters including between 2 and 6 isolates each, whereas the remaining 55 were singletons (Fig. 2). Isolates belonging to phylogroup D were more likely to be clustered (26 of 34 isolates [76.5%] included into 10 clusters) than those belonging to phylogroups B1 (40 of 64 isolates [62.5%] included into 15 clusters) and A (11 of 30 isolates [36.7%] included into 5 clusters) (Fig. 2).
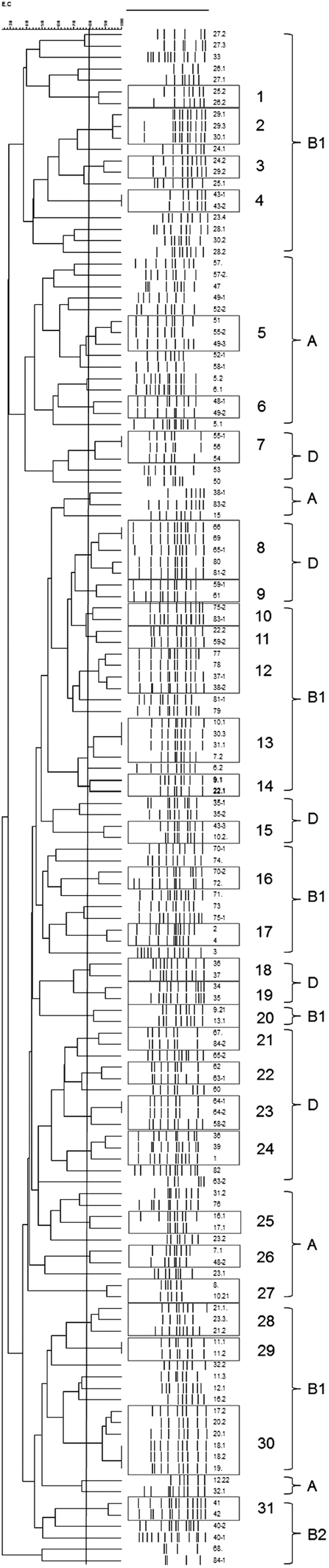
Dendrogram of enterobacterial repetitive intergenic consensus sequence–polymerase chain reaction fingerprints of Escherichia coli strains isolated from retail chicken meat, Italy.
Discussion
Antimicrobial resistance is an issue of great concern because of the serious limitation and even the risk for loss of effective antimicrobial treatment of infections. There are undeniable evidences that foods from many different animal sources and in all stages of processing contain resistant bacteria and their resistance genes (Cabello et al., 2006; Dierikx et al., 2013). It has also been documented that they can be transferred to humans (Witte, 2000; Perreten, 2005; Cabello et al., 2006). According to the literature, antimicrobial-resistant E. coli strains carried by healthy animals seem to link directly with human infection cases (Marshall and Levy, 2011). In particular, studies about bloodstream infections caused by E. coli in Europe suggest that poultry might be an important source of resistant isolates (Johnson et al., 2006; Johnson et al., 2007; Vieira et al., 2011).
In this study, MDR E. coli were recovered from 66.9% of poultry meat samples on sale in Palermo, Italy. These results appear to be consistent with findings from other European and non-European countries (EFSA Panel on Biological Hazards, 2011). In three similar studies in Portugal, Spain, and the Netherlands, indeed, MDR E. coli strains were isolated in 60%, 90%, and 94% of chicken carcasses at the retail level, respectively (Costa et al., 2009; Cohen Stuart et al., 2012; Silva et al., 2012). However, results from different studies are difficult to compare because of different settings and sample-processing methods. Our findings also can be cautiously compared with those of previous studies because of our MDR E. coli detection approach.
All the MDR E. coli isolates tested phenotypically positive for ESBLs and in a lower proportion for AmpC β-lactamases. In 3 similar studies in Germany, Spain, and the Netherlands, ESBLs were found in 88.6%, 79.7%, and 79.8%, respectively, of chicken carcasses, fecal samples, and meat, respectively (Costa et al., 2009; Cohen Stuart et al., 2012; Reich et al., 2013). Again, our detection method makes it difficult to meaningfully compare our ESBL-producing E. coli prevalence with those previously reported.
The predominant ESBL gene families among our E. coli isolates were bla SHV, bla TEM, and bla CTX-M-1 group, which is comparable with previous data from European countries. TEM, SHV, and CTX-M-1 group are reported as the most prevalent β-lactam resistance determinants in the food animal reservoir (Costa et al., 2009; Cohen Stuart et al., 2012; Reich et al., 2013). AmpC β-lactamases are less frequently reported among E. coli isolates, with the family CMY-2 being the most prevalent worldwide. Likewise, in our study a prevalence of about 10% plasmid-mediated AmpC-producing E. coli was detected (Cohen Stuart et al., 2012). However, plasmidic AmpC was always attributed with the CIT-like type. The absence of AmpC genes in some phenotypically positive isolates might indicate a different mechanism of resistance, such as overexpression of chromosomal AmpC (Peter-Getzlaff et al., 2011).
The prevalence of plasmid-mediated quinolone resistance was very high among our isolates, which highlights a serious clinical and public health issue of concern. qnrA was the only PMQR determinant detected in all ciprofloxacin resistant isolates accounting for a prevalence of about 90%, substantially higher than any other previous report. For instance, in two studies from Czech Republic and Portugal, PMQR genes were detected only in 4% and 5.5% of strains, respectively (Literak et al., 2013; Jones-Dias et al., 2013). This finding could be tentatively related to the extensive use of fluoroquinolones in the poultry industry in the southern European countries, including Italy (Cerquetti et al., 2009). Transmission of qnr-positive E. coli isolates to humans through the food chain has been previously hypothesized (Fàbrega et al., 2008).
The majority of the isolates in our study belonged to the phylogenetic group B1, followed by groups D, A and B2. These results are consistent with previously reports from both European and non-European countries (Hannah et al., 2009; Lyhs et al., 2012; Obeng et al., 2012; Hiki et al., 2014). Interestingly, according with Walk et al. (2007), the majority of the E. coli strains able to persist in the environment belong to the B1 phylogenetic group.
Special attention has been focused on retail meat as a vehicle for transmission of ExPEC from animals to humans (Johnson et al., 2005b). Within our collection, 22 (16.4%) E. coli isolates could be classified as ExPEC. Johnson et al. (2005b) found 21% of 110 E. coli isolates from chicken products to meet the criteria for ExPEC identification. In a further study by Lyhs et al. (2012), 22% of E. coli strains from chicken samples could be classified as ExPEC. Xia et al. (2011) found the highest rate of ExPEC strains among different retail meats in ground turkey (23.5%) and chicken breasts (20.2%). Of interest, our D isolates were more frequently antibiotic-resistant, ESBL and AmpC carrying compared with those belonging to phylogroups A and B1, which is consistent with literature reporting a close combination of antibiotic resistance and virulence traits (Pitout, 2012). B2 phylogroup isolates behaved similarly but their limited number does not allow meaningful conclusions to be drawn.
Genotyping by the ERIC-PCR method showed 55 unique DNA banding profiles and a proportion of ≈60% of isolates grouped into 31 small clusters. These findings support a high genetic heterogeneity of E. coli isolates, despite their isolation in a restricted geographic area. Such a finding is a further source of concern, emphasizing that emergence of MDR E. coli in chicken is not resulting from the clonal expansion of a few successful bacterial strains, but more likely from a complex interaction between several different clones and laterally transferred resistance genes (Olsen et al., 2014).
Our study has some limitations. First, a relatively small number of samples were analyzed from a restricted geographic area. This, because of the presumable homogeneity of the husbandry techniques, could lead to an overestimate of the prevalence of MDR E. coli. On the other hand, our microbiological approach could have determined an underestimate of the heterogeneity of the cephalosporin- and/or ciprofloxacin-resistant E. coli isolates and biased our evaluations. Moreover, selection of only lactose-positive colonies might have further contributed to underdetecting samples containing MDR E. coli. Characterization of resistance genetic determinants was not further defined because the wide heterogeneity of their distribution made this approach unaffordable.
In summary, our results show that in Sicily a large proportion of retail chicken meat samples carry cephalosporin- and ciprofloxacin-resistant E. coli. A high prevalence of ESBL and PMQR determinants was also found in these isolates, which raises a serious public health concern. A prudent use of antimicrobial agents is urgent in veterinary as well as in human medicine to minimize the selection and spread of antibiotic-resistant strains and their resistance genetic determinants.
Footnotes
Acknowledgments
We are indebted to Dr. James R. Johnson and his team (VA Medical Center, Minneapolis, MN) for sending us all reference E. coli strains used in this work and for giving us many useful suggestions.
Disclosure Statement
No competing financial interests exist.