
Abstract
A total of 221 Salmonella enterica from raw pork (n=64), raw chicken (n=80), and humans (n=77) were characterized for antimicrobial resistance phenotypes and genotypes and virulence plasmid–associated genes. Most Salmonella isolates (95.9%) were multidrug resistant and exhibited high resistance to sulfamethoxazole (96.4%), streptomycin (93.2%), spectinomycin (76.5%), tetracycline (73.3%), ampicillin (70.1%), and trimethoprim (60.2%). Forty-one percent of all isolates were intI1-positive, of which 60% carried class 1 integrons with variable region ranging in size from 0.2 to 2.0 kb. Six integron profiles (IP-I to IP-VI) were defined. The dfrA12-aadA2 cassette was most prevalent (66.7%). Class 1 integrons with the dfrA12-aadA2 cassette in five pork isolates could be horizontally transferred. Three pork isolates carried Salmonella genomic island 1 (SGI1), of which a serovar Anatum harbored SGI1 gene cluster located between thdF and int2. Two single-point mutations (i.e., G-259-T and C-248-T) in gyrA leading to Asp-87-Tyr and Ser-83-Phe substitutions in GyrA, respectively, were detected. Of all plasmid-mediated quinolone resistance genes tested, only qnrS (4.1%) and qnrB (1.8%) were found. The virulence plasmid–associated genes including spvC, pefA, and rck were identified in 8.1%, 1.8%, and 1.4% of all Salmonella isolates, respectively.
Introduction
M
Northeastern Thailand, the largest region with the most population of the country, shares a common border with Laos PDR to the north and east and Cambodia to the southeast, where millions of people and animals travel across each year (Murshid et al., 2005). Thailand's cross-border trade with neighboring countries has boomed and contributed to a significant amount of income. Livestock (including pigs, poultry, and cattle) and their products are the predominant cross-border merchandise that are traded legally and illegally (Murshid et al., 2005). Such relocation at the border undoubtedly has contributed to emergence and distribution of various bacterial pathogens including MDR Salmonella and their resistance determinants. This would affect not only the border area but also other parts of the two countries and eventually other parts of the world. While Salmonella is the second most common cause of diarrhea in Thailand, chicken and pig products are important exporting goods (Bodhidatta et al., 2002). AMR in Salmonella has been widely studied in Thailand, but this is not always the case in neighboring countries. Reports showed that diarrhea and food poisoning are the most common diseases among Cambodian patients who came for treatment in Thai hospitals along the border (Lay et al., 2011). Septicemic salmonellosis caused by MDR Salmonella, particularly Salmonella Choleraesuis, has increased in Cambodia as well (Lay et al., 2011). In Laos PDR, Salmonella was commonly found in pigs and pork carcasses in slaughterhouses (Boonmar et al., 2008). However, data on AMR is still limited.
To date, the prevalence, characteristics, and transfer of AMR in Salmonella has been extensively studied but mostly limited to developed countries. Therefore, this study aimed to (1) determine antimicrobial susceptibility; (2) characterize class 1 integrons and the SGI1 variants; and (3) examine the presence of virulence plasmid–associated genes in the Salmonella isolates from pork, chicken meat, and humans in Northeastern Thailand.
Materials and Methods
Bacterial strains
A total of 221 Salmonella isolates from raw pork (n=64), raw chicken meat (n=80), and humans (n=77) were obtained from samples collected in 5 provinces in Northeastern Thailand, including Kalasin, Khon Kaen, Loei, NongKhai, and Roi Et during 2010–2013 (Table 1). Sample collection was performed at the largest municipal slaughterhouses with the highest number of slaughtered pigs or broilers per day and at local markets receiving the carcasses from the selected slaughterhouses. Pig carcass swabs and broiler carcasses were taken at the end of the slaughter line before transporting to retail markets. Retail pork cuts and broiler carcasses were obtained from the selected local markets. All the human isolates were originated from rectal swabs of diarrhea patients at Khon Kaen Hospital. However, dietary history (including chicken or pork consumption prior to illness onset) and information on food-animal rearing was not available. The Salmonella strains were isolated using the standard methods as described in ISO6579:2002 (E) (ISO, 2002) at Department of Veterinary Public Health, Faculty of Veterinary Medicine, Khon Kaen University and were subjected to serotyping at Center of Antimicrobial Resistance in Foodborne Pathogens (in cooperation with the World Health Organization), Faculty of Veterinary Science, Chulalongkorn University. Only 1 colony of each serotype was collected from each positive sample and stored as 20% glycerol stocks at −80°C. Three Escherichia coli strains carrying qnrA, qnrB, or qnrS were used as positive controls (Wu et al., 2008).
Antimicrobial susceptibility test
Minimum inhibitory concentrations (MICs) were determined using a twofold agar dilution (CLSI, 2008). Ten antimicrobials and their breakpoints were as follows: ampicillin (AMP, 32 μg/mL), cefoperazone (CEF, 64 μg/mL), ciprofloxacin (CIP, 4 μg/mL), chloramphenicol (CHL, 32 μg/mL), gentamicin (GEN, 8 μg/mL), streptomycin (STR, 32 μg/mL), spectinomycin (SPC, 128 μg/mL), sulfamethoxazole (SUL, 512 μg/mL), tetracycline (TET, 16 μg/mL), and trimethoprim (TRI, 16 μg/mL) were tested. All antibiotics were purchased from Sigma-Aldrich (St. Louis, MO). Pseudomonas aeruginosa ATCC 27853 and E. coli ATCC 25922 served as control strains.
General DNA manipulations
Template DNA for polymerase chain reaction (PCR) were whole-cell DNA prepared using the boiling lysate procedure (Levesque et al., 1995) except when indicated. Chromosomal DNA was isolated using the GF-1 Nucleic Acid extraction kit (Vivantis, Selangor Darul Ehsan, Malaysia). Plasmid DNA was obtained by using QIAprep Mini-spin kit (Qiagen, Hilden, Germany). PCR products were purified using Nucleospin® Gel and PCR cleanup (Mccherey-Nagel, Düren, Germany).
Identification of class 1 integrons, SGI, and virulence plasmid–associated genes
All PCR reactions were performed by using Red dye PCR Master Mix Genei (Merck, Darmstadt, Germany).
Class 1 integrons were characterized in all the Salmonella isolates using conserved segment (CS)-PCR as previously described (Levesque et al., 1995; Chuanchuen et al., 2007). CS-PCR amplicons were gel purified and submitted for nucleotide sequencing at First Base Laboratories (Selangor Darul Ehsan, Malaysia). The DNA sequences obtained were compared with the published data in the GenBank database, available at:
Association of class 1 integrons with SGI1 was investigated by PCR in all the Salmonella isolates carrying class 1 integrons with resistance gene cassettes (Doublet et al., 2003; Khemtong and Chuanchuen, 2008). The left (thdF) and right (S044–yidY) junction of SGI1 were first determined and the organization of antibiotic-resistance gene clusters for SGI1-like element was examined using different combinations of primers.
The presence of Salmonella-virulence plasmids including spvC, pefA, and rck was screened in all isolates (Chiu and Ou, 1996; Guerra et al., 2002; Skyberg et al., 2006). Plasmid DNA was extracted from the spvC, pefA, or rck–positive Salmonella strains and used as templates for PCR amplification to confirm plasmid localization of the genes.
Detection of mutation(s) in the quinolone resistance-determining region (QRDR) of the gyrA and parC
Mutation(s) in the QRDR of the gyrA and parC were examined in all the ciprofloxacin-resistant Salmonella strains, including the chicken (n=2) and the human (n=4) isolates by nucleotide sequencing analysis using the forward and reverse primers as previously described (Chuanchuen and Padungtod, 2009). The QRDRs of gyrA and parC obtained from two ciprofloxacin-susceptible strains were included as controls. All the Salmonella isolates were screened for the presence of qnr genes (qnrA, qnrB, and qnrS), aac(6′)-Ib-cr and qepA (Park et al., 2006; Yamane et al., 2008; Stephenson et al., 2010).
Conjugation experiments
Filter mating assay was performed as previously described (Khemtong and Chuanchuen, 2008). All Salmonella isolates with class 1 integrons carrying resistance gene cassettes served as donors (n=45), and the spontaneous rifampicin-resistant derivatives of E. coli K-12 MG1655 (MG1655 rifr, MIC=256 μg/mL) were recipients. In E. coli transconjugants, transfer of class 1 integrons and resistance gene cassettes was confirmed by PCR using plasmid DNA as described above.
Results
Antimicrobial resistance of the Salmonella isolates
The overall resistance rates to sulfamethoxazole, streptomycin, spectinomycin, tetracycline, ampicillin, trimethoprim, chloramphenicol, gentamicin, cefoperazone, and ciprofloxacin were 96.4%, 93.2%, 76.5%, 73.3%, 70.1%, 60.2%, 36.7%, 36.7%, 10.4%, and 3.2%, respectively. All the Salmonella isolates were resistant to at least one antibiotic (Fig. 1). Most (95.9%) exhibited multidrug resistance (being resistant to at least 3 different classes of antimicrobial agents). AMR patterns commonly found are shown in Table 2.
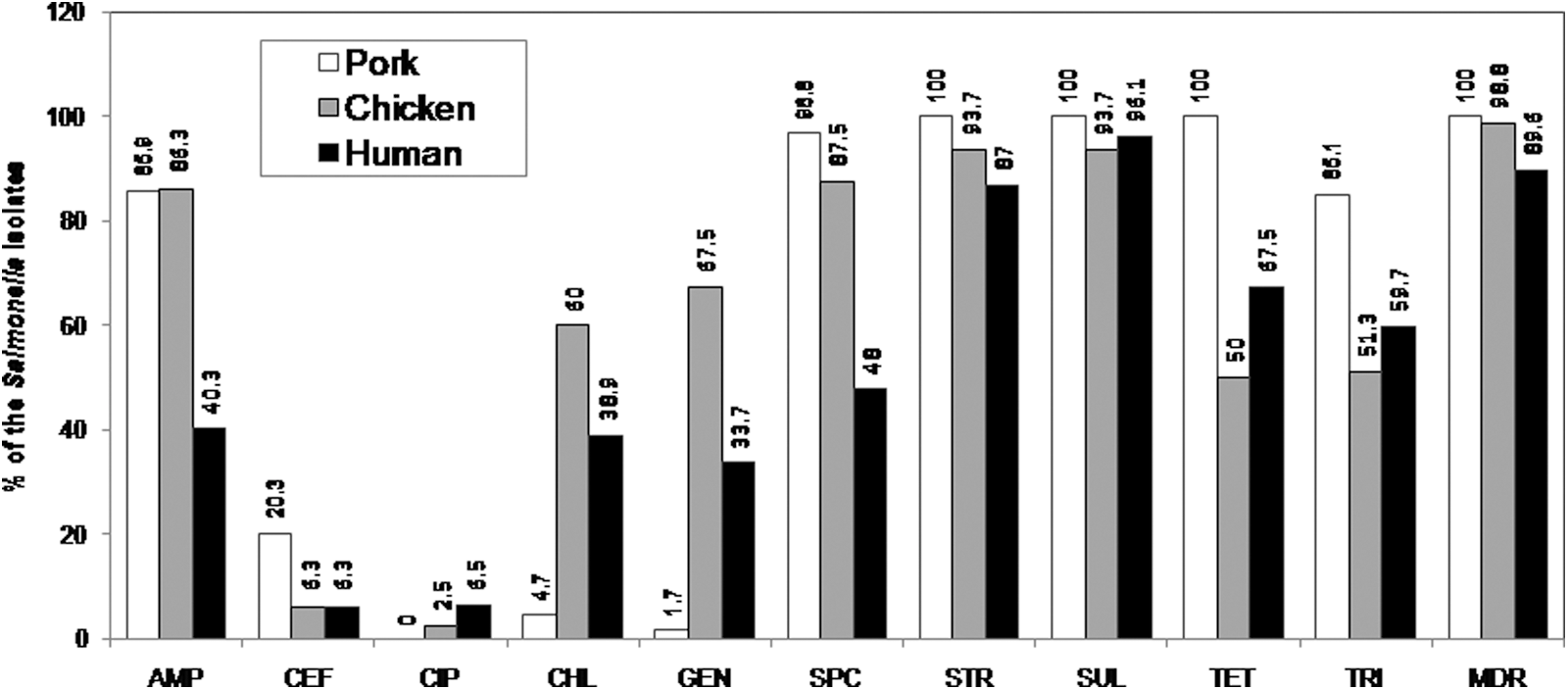
Frequency of resistance to 10 antimicrobial agents in Salmonella (n=221) from retail pork (n=64), retail chicken (n=80), and humans (n=77). AMP, ampicillin; CEF, cefoperazone; CIP, ciprofloxacin; CHL, chloramphenicol; GEN, gentamicin; SPC, spectinomycin; STR, streptomycin; SUL, sulfamethoxazole; TET, tetracycline; TRI, trimethoprim; MDR, multidrug resistant.
Only the antimicrobial resistance patterns represented by at least five isolates are shown.
The superscript numbers in parentheses represent the numbers of isolates with corresponding chracteristics indicated by the adjacent superscript.
The most common resistance pattern in serovars Rissen from pork (n=10).
The most common resistance pattern in serovars Anatum from pork (n=12).
The most common resistance pattern in serovars Schwarzengrund from chicken (n=5).
The most common resistance pattern in serovars Rissen from chicken (n=6).
The most common resistance pattern in serovars Weltevreden from human (n=3).
AMP, ampicillin; SPC, spectinomycin; STR, streptomycin; SUL, sulfamethoxazole; TET, tetracycline; TRI, trimethoprim; CEF, cefoperazone; CHL, chloramphenicol; GEN, gentamicin.
Class 1 integrons, gene cassettes, and their horizontal transfer
Forty-one percent of the Salmonella isolates were positive to intI1 (i.e., pork, 26/64; chicken, 49/80; and human, 15/77), of which 60% carried class 1 integrons with variable regions ranging in size from 0.2 to 2.0 kb. Six integron profiles (IP-I to IP-VI) were defined (Table 3). The most prevalent resistance genes were dfrA12 (66.7%) and aadA2 (75.9%), which were combined to the dfrA12-aadA2 cassette array (66.7%). Coexistence of two distinct integrons carrying different gene cassettes (i.e., aadA2 and bla PSE-1) was identified in four isolates (IP-II).
The integron profiles (IPs) were defined by the number and the size of the polymerase chain reaction amplicons and resistance gene cassettes identified.
bla PSE-1 encodes β-lactam resistance; dfrA12 encodes trimethoprim resistance; aadA2 encodes streptomycin and spectomycin resistance; aadA4 encodes streptomycin and spectomycin resistance.
The superscript numbers in parentheses represent the numbers of isolates with corresponding characteristics indicated by the adjacent superscript.
Number of strains is indicated in parentheses.
Successful class 1 integrons transfer.
Carry both spvC and class 1 integrons.
Positive to thdF and int2 and termed SGI1-like elements.
Positive to thdF and S044–yidY of SGI1.
Nine isolates from pork and humans (4.1%) carried class 1 integrons with the 200-bp fragment of an internal segment of purG encoding enzyme phosphoribosylformylglycinamide synthetase (IP-VI). All the isolates additionally yielded a 750-bp PCR fragment of a partial codB gene mediating cytosine permease that was not located within class 1 integrons, as determined by nucleotide sequencing.
Only class 1 integrons with the dfrA12-aadA2 cassette array in five isolates from retail pork (i.e., three serovars Anatum, a serovar Virchow, and a serovar Seftenberg) could be horizontally transferred to E. coli.
Presence of SGI1-like gene cluster in Salmonella enterica Anatum
Only three pork isolates carrying class 1 integrons with resistance gene cassettes were positive to thdF. Two isolates (i.e., serovars Anatum and Albany) carried thdF and S044–yidY (IP-IV and IP-V). The others (i.e., a serovar Anatum) were positive to thdF but not S044–yidY (IP-II). The latter harbored int2 and all SGI1 antibiotic-resistance gene clusters located between thdF and int2.
Mutations within gyrA and parC QRDRs and the presence of plasmid-mediated quinolone resistance (PMQR) genes
Two point mutations in gyrA (i.e., G-259-T and C-248-T, leading to Asp-87-Tyr and Ser-83-Phe substitutions in GyrA, respectively) were detected. Four ciprofloxacin-resistant Salmonella strains (MIC=4 μg/mL) carried mutations in gyrA, including a serovar Virchow from chicken and two serovars Enteritidis and a serovar Rissen from humans. A G-283-C change in parC leading to a Val-95-Leu mutation in ParC was additionally found. This amino acid substitution was also identified in two ciprofloxacin-susceptible Salmonella control strains and therefore, could be a result of sequence variation.
Nine Salmonella isolates harbored qnrS, including a serovar Stanley (CIP MIC=4 μg/mL) from humans; a serovar Anatum (CIP MIC=4 μg/mL), five serovars Covallis (CIP MIC=2 μg/mL) from poultry, and two serovars Covallis (CIP MIC=2 μg/mL) from pork. The qnrB gene was found in two pork isolates, a serovar Rissen (CIP MIC=0.125 μg/mL), and a serovar Stanley (CIP MIC=0.125 μg/mL). None carried both PMQR genes and mutation in gyrA QRDRs.
Occurrence of Salmonella virulence plasmids
Overall, the prevalence of Salmonella plasmid virulence genes was low (i.e., spvC, 8.1%; pefA, 1.8%; and rck,1.4%) (Table 4). All the isolates yielded PCR products of the corresponding genes when using plasmid DNA as templates. Three Salmonella isolates (i.e., two serovars Rissen from pork [IP-1] and a serovar Panama from human [IP-III]) carried both class 1 integrons and spvC (Table 3) but none of class 1 integrons was located on conjugative plasmid. The latter was confirmed by PCR using the E. coli transconjugant plasmid DNA.
Discussion
In general, the majority of Salmonella serotypes can affect different hosts, but some are host specific. In this study, some serotypes were predominantly found in particular hosts such as serovars Rissen in pork and chicken, serovars Schwarzengrund in chicken, serovars Anatum in pork, and serovars Weltevreden in humans (Table 1). These findings suggest potential host-specific population dynamics of Salmonella.
The data in this study highlighted a high rate of AMR among the Salmonella isolates from pork, chicken meat, and diarrhea patients in Northeastern Thailand. The frequency of MDR isolates was higher than that in a previous report in the Mekong Delta, Vietnam (Ogasawara et al., 2008). The observation of ciprofloxacin resistance in the chicken and human isolates, even at a low rate, raises particular concern because fluoroquinolone is the recommended drug for invasive salmonellosis. These results may suggest the wide use of the antimicrobials in either animals or humans in Thailand. However, it should be noted that spread of AMR bacteria may be a result of nonantibiotic selective pressure (e.g., disinfectants and heavy metals) (Langsrud et al., 2004).
Overall, the prevalence of class 1 integrons was high (40.7%). It is comparable to our previous study in the pork and human isolates from Northern Thailand (Wannaprasat et al., 2011) but higher than that reported in other countries (Thong and Modarressi, 2011). It is interesting to observe that class 1 integrons were more common among the chicken isolates than the pork and human isolates. The reason for this observation, however, remains unclear. The pork and chicken isolates originated from five provinces, while the human isolates were from one province. This source difference together with the possible sampling bias may limit the ability to compare the resistance profile among the isolates.
A particular concern was the presence of the same class of integrons (i.e., IP-I, II, and VI) in the pork, chicken, and human isolates, suggesting circulation and horizontal transfer of these resistance determinants in the food chain. Therefore, the food chain serves as a potential route for the introduction of animal- and environment-associated antibiotic resistant bacteria into the human. However, transmission of AMR from animals to humans can take through other routes (e.g., direct handling, close contact, and the spreading of slurry).
Class 1 integrons with incomplete purG (IP-VI) were previously identified as a PCR artifact (Lindstedt et al., 2003) or as a small integron specific to serovar Typhimurium (Daly et al., 2000). In this study, the partial purG gene was part of class 1 integron but not serovar specific. All the isolates in IP-VI had an additional-incomplete codB fragment that was found to be a PCR artifact by DNA sequencing. In contrast, this defective codB gene was part of class 1 integrons in our previous study (Khemtong and Chuanchuen, 2008).
Almost half of the intI1-positive isolates (40%) carried empty integrons. These are likely due to excision of the previously inserted gene cassettes when the antimicrobial selective pressure is diluted (Rosser and Young, 1999) and are available for new-emerging resistance genes. Therefore, minimized antimicrobial use may significantly reduce the integrons with resistance gene cassettes and their horizontal transfer among bacteria, leading to reduction of resistant bacteria.
It is of particular interest to observe a serovar Anatum from pork carrying SGI1-like elements with SGI1-gene cluster located between thdf and int2. This is in agreement with a previous study reporting a serovar Meleagridis containing SGI1-like gene cluster with the right junction int2 retron phage (Ebner et al., 2004). The int2 gene is part of a retron sequence that was found only in Salmonella Typhimurium (Doublet et al., 2003). Therefore, identification of SGI1-like elements on a non-Typhimurium isolate confirmed that SGI1 genes are (partly) mobile (Ebner et al., 2004), and this could be a possible route for spreading of multidrug resistance. Horizontal transfer of class 1 integrons and the presence of SGI-1 were limited to the pork isolates in this study. A possible explanation may be the different selective pressure created by different antibiotics used in different food animals and humans.
In this study, mutations were observed only in gyrA QRDRs. Two point mutations in GyrA (i.e., Asp-87-Tyr and Ser-83-Phe) were previously identified in Salmonella (Wang et al., 2015). It was shown that those with mutation at position 87 exhibited higher ciprofloxacin resistance than those with mutation at position 83 (Wang et al., 2015). In contrast, all four isolates with mutation in gyrA QRDRs had the same ciprofloxacin MIC value (i.e., 4 μg/mL). Still, it cannot be concluded that the single mutation in gyrA QRDRs was a sole mechanism responsible to the MIC value observed, and other uncharacterized mechanisms may exist.
In this study, ciprofloxacin MIC of the qnr-carrying isolates varied and two pig isolates carrying qnrB had low ciprofloxacin MIC (0.125 μg/mL). This supports that the impact of PMQR genes on ciprofloxacin MIC level varies, and their presence does not always cause a high level of fluoroquinolone resistance (Robicsek et al., 2006). The PMQR determinants are likely to provide coselection of resistance genes for other antibiotics. Two ciprofloxacin-resistant isolates had neither gyrA mutations nor PMQR determinants, indicating the existence of other resistance mechanisms (e.g., the efflux pumps and decreased membrane permeability).
The spvC gene was most frequently found (8.1%). This is not surprising because the spv operon is common to all Salmonella virulence plasmids (Zou et al., 2012). The pefA and rck genes were rare and limited to serovars Enteritidis and Typhimurium. This is most likely due to the serovar specificity of these two genes (Skyberg et al., 2006). It should be also noted that virulence plasmid–associated genes can be presented on a chromosome (Boyd and Hartl, 1998). The isolate with the genes on genomic DNA (if any) would also yield a positive result. Some Salmonella carried both class 1 integrons and spvC but not on the same conjugative plasmid, suggesting the presence of multiple plasmids or chromosomal integration of the virulence plasmid.
In summary, the results in this study highlight the high rates of MDR phenotype and class 1 integrons among the Salmonella isolates from pork, chicken, and humans. It suggests that antimicrobials should be appropriately used in food animals and humans to preserve the efficacy of the existing drugs and to minimize the risk of transfer of resistant foodborne pathogens to humans.
Footnotes
Acknowledgments
We thank Genetic Strains Research Center, National Institute of Genetics, Japan for providing E. coli K12 MG1655. This work was financially supported by the Research Grant for Mid-Career University Faculty RMU5680051 cofunded by Thailand Research Fund (TRF), the Faculty of Veterinary Science, and Chulalongkorn University. It was partially supported by the 90th anniversary of Chulalongkorn University fund. NS is a recipient of the Royal Golden Jubilee PhD program PHD/006/2553, cofunded by TRF and Chulalongkorn University.
Disclosure Statement
No competing financial interests exist.