
Abstract
Salmonella enterica subspecies enterica serovar 4,[5],12:i:-, a monophasic variant of Salmonella Typhimurium, has emerged as one of the most common serotypes related to human salmonellosis. In this study, the 22 isolates of S. 4,[5],12:i:- from food animals were identified by a specific multiplex polymerase chain reaction between 2009 and 2012. The isolation rate of S. 4,[5],12:i:- accounted for 1.7% (22/1271) of Salmonella spp. isolates from food animal origins: more specifically, 7.6% (18/235) from pigs and 0.6% (4/686) from chickens. The predominant S. 4,[5],12:i:- isolates in Korea belonged to phage type DT193 (12/22) with ampicillin–streptomycin–sulfonamide–tetracycline (ASSuT) resistance pattern (9/22). The XbaI-pulsed-field gel electrophoresis (PFGE) analysis revealed 11 different pulsotypes, and the major X-1 pattern was shared by 8 isolates. The isolates belonging to pattern X-1 were further subdivided into three BlnI-PFGE patterns and four variable-number tandem-repeat analysis (MLVA) allele combinations. The combining of MLVA and PFGE data could be valuable in characterizing highly clonal strains and discriminating their epidemiological relationship.
Introduction
S
Various studies have been performed in other countries on this serotype. Despite the diversity of phage types, resistance profiles, and molecular types found in the S. 4,[5],12:i:- strains, many of the isolates were reported as having several analogous characteristics. Most dominant clones of S. 4,[5],12:i:- belonging to phage type DT193, DT120, and U302, express resistances to ampicillin, streptomycin, sulfonamides, and tetracycline (ASSuT), and exhibit a few predominant pulsed-field gel electrophoresis (PFGE) patterns and multilocus variable-number tandem-repeats analysis (MLVA) profiles (Gallati et al., 2013; Arguello et al., 2014).
The present study was undertaken to investigate the rate of occurrence of S. 4,[5],12:i:- in Korean food animals. Furthermore, we present the phenotypic and molecular characterization of S. 1,4,[5],12:i:- isolates with respect to their phage types, antimicrobial resistance patterns, and DNA fingerprints using PFGE and MLVA.
Materials and Methods
The Salmonella spp. isolates were obtained for research in Animal and Plant Quarantine Agency (QIA, Anyang, Korea), or submitted for diagnostic investigation from regional veterinary laboratories, or received from Korean Veterinary Antimicrobial Resistance Monitoring System participating centers. We identified the serotypes of Salmonella isolates by agglutination with diagnostic antisera (Difco Laboratories, BD, Sparks, MD) according to the White–Kauffmann–Le Minor scheme, and then a multiplex PCR assay was performed to differentiate among Salmonella Typhimurium and S. 4,[5],12:i:-. A multiplex polymerase chain reaction (PCR) protocol that allows simultaneous amplification of a fragment between the genes fljB and fljA and the phase-2 flagellar gene (fljB) was used (Tennant et al., 2010). No S. 4,[5],12:i:- isolates harbored any detectable fljB PCR fragment, which means that they were true S. 4,[5],12:i:- according to the European Food Safety Authority (EFSA) guidelines (EFSA, 2010). Phage typings were performed using the protocol reported by Anderson et al. (1977), and following the guidelines defined for Salmonella Typhimurium by the Health Protection Agency (Colindale, UK).
The isolates were tested for antimicrobial susceptibility by the disk-diffusion method according to the Clinical and Laboratory Standards Institute document M31-A3. The antimicrobial disks (BBL Sensi-Disk; BD) were used in this study as follows: ampicillin (10 μg), amoxicillin/clavulanic acid (20/10 μg), cefotaxime (30 μg), cefepime (30 μg), streptomycin (10 μg), gentamicin (10 μg), neomycin (30 μg), tetracycline (30 μg), colistin (10 μg), sulfisoxazole (250 μm), sulfamethoxazole/trimethoprim (23.75 μg/1.25 μg), chloramphenicol (30 μg), nalidixic acid (30 μg), and ciprofloxacin (5 μg). Escherichia coli ATCC 25922 was used as quality-control strains.
Genetic relatedness was determined by PFGE analysis after digestion of DNA with XbaI and BlnI restriction enzymes (Takara Bio Inc., Shiga, Japan) according to the PulseNet standardized protocol (Ribot et al., 2006). The fragment restriction patterns were analyzed using Bionumerics, version 5.1 (Applied Maths, Sint-Martens-Latem, Belgium). The band patterns in the range of 28.8–1135.0 kb were compared.
MLVA was used to amplify five loci by using previously designed primers, as described in an earlier study (Lindstedt et al., 2004). The allelic combinations were recorded in the following order: STTR9-STTR5-STTR6-STTR10-STTR3. The PCR products were analyzed by ABI 3130xl (Applied Biosystems, Foster City, CA) and VNTR allele number assignment was performed as described by Larsson et al. (2009).
Results and Discussion
The 1271 Salmonella spp. isolates recovered from food-animal feces and organ samples in farms and slaughterhouses between 2009 and 2012 included 108 isolates from cows, 235 isolates from pigs, 686 isolates from chickens, and 232 isolates from ducks. The 22 S. 4,[5],12:i:- isolates were identified from 1271 Salmonella spp. isolates including 18 isolates from pigs and 4 isolates from chickens. The isolation rate of S. 4,[5],12:i:- accounted for 1.7% (22/1271) of all Salmonella spp. isolates from food animal origin, including 7.6% (18/235) in pigs and 0.6% (4/686) in chickens. Similarly, pigs were assumed to be the main reservoir of S. 4,[5],12:i:- in several European countries (Hauser et al., 2010; Hopkins et al., 2010). The variant of Salmonella Typhimurium antigenic formula was S. 4,5,12:i:- in 15 (68.2%) of the 22 isolates, while the other 7 (31.8%) were serotyped as S. 4,12:i:-. In agreement with data published by Hauser et al. (2010), the majority of the isolates presented the factor O:5 in their antigenic formula.
Phage typing showed 5 different types with the following distribution: 12 isolates belonged to DT193, 4 isolates to DT208, 3 isolates to DT104b, 2 isolates to U291, and 1 isolate was RDNC (reaction pattern did not conform to the phage scheme). Ampicillin and tetracycline resistance were most frequently detected in 16 (72.7% of the total isolated isolate) of S. 4,[5],12:i:- isolates, followed by sulfisoxazole (50%) and streptomycin (45.5%). A low level of resistance was observed toward nalidixic acid (13.6%), gentamicin (9.1%), and chloramphenicol (4.5%). All the isolates were susceptible to amoxicillin/clavulanic acid, cefotaxime, cefepime, neomycin, colistin, sulfamethoxazole/trimethoprim, and ciprofloxacin. Nine S. 4,[5],12:i:- isolates had the antimicrobial resistance pattern ASSuT. Especially, this ASSuT resistance profile was present only in the DT193 isolates. The phage type DT193 and resistance profile ASSuT of S. 4,[5],12:i:- isolates were the most representative in this study.
As in European countries, the strains of Salmonella 4,[5],12:i:- DT193 ASSuT also circulated in Korea (Barco et al., 2014; Hopkins et al., 2010). It seems that DT193 isolates are associated with this ASSuT resistance profile more than other phage types in S. 4,[5],12:i:- and Salmonella Typhimurium strains (Arguello et al., 2014). Also, S. 4,[5],12:i:- DT208, DT104b, and U291 have previously been reported in Europe and the United States (Hopkins et al., 2010; Trupschuch et al., 2010; Arguello et al., 2014).
The S. 4,[5],12:i:- isolates were molecularly characterized by PFGE and MLVA. Eleven unique XbaI-PFGE profiles were identified among the 22 isolates. The major XbaI-PFGE pattern, X-1, was shared by eight isolates. All X-1 isolates were phage type DT193 and from pigs. Twelve S. 4,[5],12:i:- DT193 isolates were clustered together by XbaI-PFGE patterns (X-1, X-2, X-3, and X-4) with 85.8% similarity. There are also 12 BlnI-PFGE patterns that were identified among the 22 isolates.
MLVA divided the isolates into 17 different profiles (Fig. 1). The most variable loci were STTR5 and STTR6 with six and eight different profiles, respectively. The STTR3 and STTR9 loci had few variations, resulting in four different profiles. The locus STTR10 was consistently absent in 81.8% (18/22) of the isolates, indicating the lack of the typical Salmonella Typhimurium virulence plasmid.
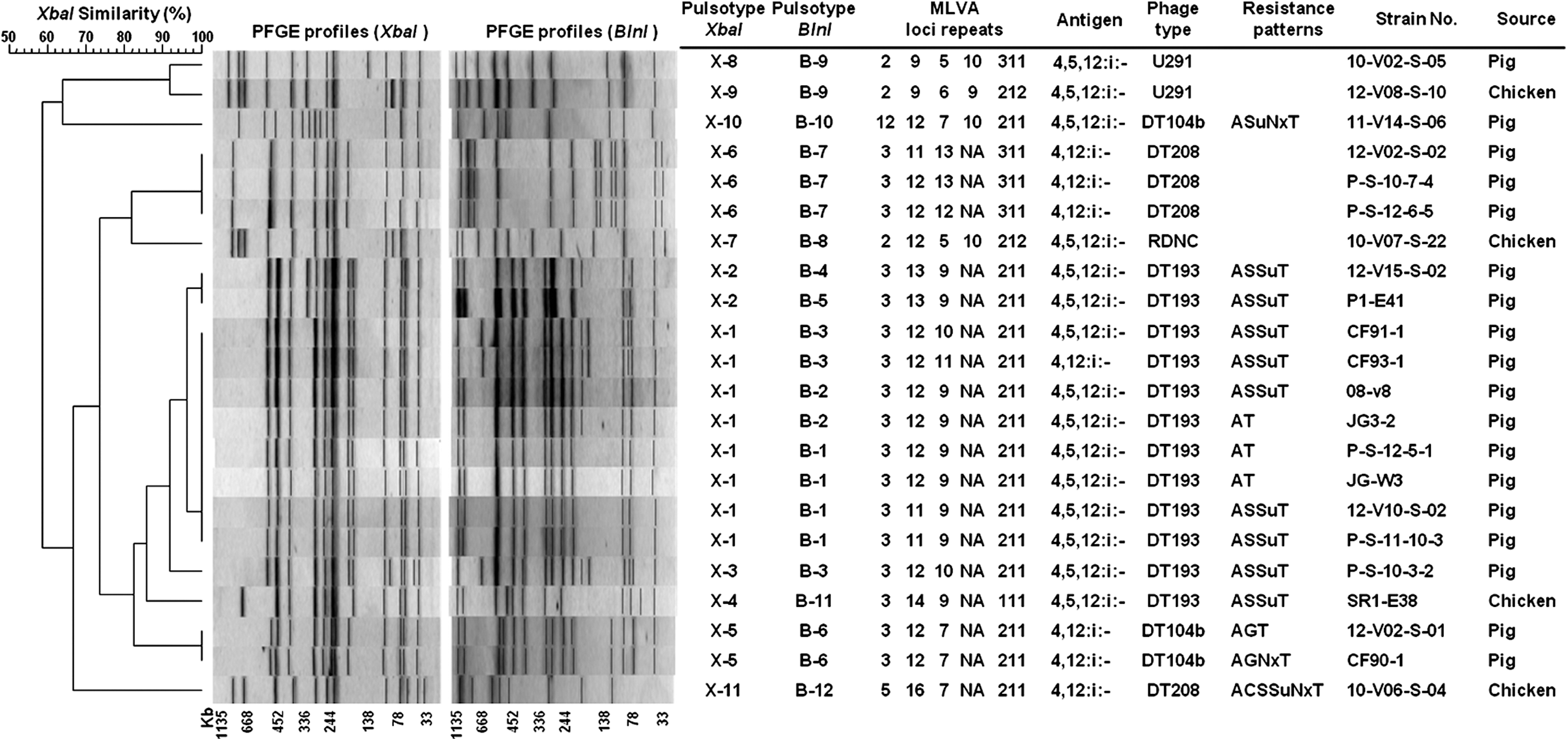
Dendrogram of Korean monophasic Salmonella 4,[5],12:i:- isolates based on pulsed-field gel electrophoresis (PFGE), characteristics, and type assignment by different typing methods. PFGE dendrogram, types, and profiles obtained with XbaI and BlnI restriction enzymes using Dice coefficient and unweighted-pair group method with arithmetic mean (optimization, 1.0%; tolerance, 1.0%) are shown. Multilocus variable-number tandem-repeat analysis (MLVA) loci repeats were composed of five numbers indicating repeat unit for each locus, with the following order: STTR9-STTR5-STTR6-STTR10-STTR3. Resistance patterns: A, ampicillin; C, chloramphenicol; G, gentamicin; S, streptomycin; Su, sulfisoxazole; Nx, nalidixic acid; T, tetracycline. Antigen composition, Phage type, Source, and Strain No. are indicated.
All isolates with MLVA profiles 3-12-9-NA-211 and 3-11-9-NA-211 belonged to XbaI-PFGE cluster X-1. The BlnI-PFGE and MLVA profiles were more scattered among the obtained XbaI-PFGE profiles. The isolates belonging to pattern X-1 were further subdivided into three BlnI-PFGE patterns and four MLVA allele combinations.
In conclusion, this study shows that S. 4,[5],12:i:- was present in Korean food animals, as in other countries, and it seemed that they are most related to pigs. The strains of S. 4,[5],12:i:- DT193 ASSuT were predominant in Korean food animals. Moreover, the combination of PFGE and MLVA could be valuable to characterize highly clonal strains and to discriminate their epidemiological relationship.
Footnotes
Acknowledgment
This work was supported by a grant from the Animal and Plant Quarantine Agency, Ministry of Agriculture, Food and Rural Affairs, Republic of Korea.
Disclosure Statement
No competing financial interests exist.