
Abstract
Mycobacterium avium subsp. paratuberculosis (MAP) is the established causative agent of Johne's disease in cattle and other ruminants, and it has also been speculated to be a putative etiological agent of several human autoimmune diseases. It is acknowledged that dairy products deriving from infected animals play a role (could be vehicles) in exposing humans to MAP. MAP could stimulate the human immune system by means of their complex antigen (in the case of lipids, multivalent antigens) and may modulate it, acting as adjuvant molecules such as Freund's complete adjuvant. The immune system might be abnormally stimulated by the constant presence of MAP antigens (for example, in the dairy products), and this might be particularly relevant in genetically predisposed individuals. However, there is limited understanding about the current human exposure to MAP. The present study analyzed the antibody recognition profile of MAP lipophilic antigens in a cohort of 126 healthy Japanese. We measured the serum levels of total immunoglobulin G (IgG) and subclasses targeting MAP surface antigens through ethanol vortex indirect enzyme-linked immunosorbent assay (EVELISA) by using serum absorbed with Mycobacterium phlei. Elevated IgG (especially IgG1 and IgG4) responses were observed in 14% of the sera. To assess the specificity of EVELISA, the same samples were analyzed by means of a commercially available Johnelisa II kit. It was noteworthy that a high degree of correlation was observed when comparing the two methodologies (r s=0.7, p<0.0001). Moreover, in order to investigate the specificity of the binding, inhibition assay experiments were carried out also searching for antibodies against Bacillus Calmette-Guérin antigens, but no cross-reaction was observed. The result obtained represents the first evidence implying that the Japanese population is exposed to MAP, and additionally the existence of a foodborne chain of exposure that transmits MAP antigens to humans.
Introduction
M
Constant exposure to MAP in humans can in turn modulate their immune system. This process is essentially confirmed by the following: the high infection rate and the elevated prevalence of paratuberculosis worldwide (Nielsen and Toft, 2009; Fernandez-Silva et al., 2014; Wolf et al., 2014), the shedding of MAP bacilli into the milk (Patel and Shah, 2011; Stabel et al., 2014), and the consumption of contaminated dairy products by a large number of people (Grant, 2003).
According to previous evidence of human exposure to MAP, we hypothesize that the consumption of food contaminated with MAP could also alter the adaptive and innate immune responses of the host.
In spite of the fact that the prevalence of bovine paratuberculosis in Japan is 0.2% at the farm level in contrast to 70% in the United States (USDA-APHIS, 2008), Japanese people are constantly exposed to MAP since they consume a substantial quantity of dairy products imported from Western countries (Momotani, 2012a) such as Europe (Kohl and Baumgarter, 2012), Australia (Ridge et al., 2010), and the United States (Carter, 2012).
For the aforementioned reasons, and in order to determine the seroprevalence of anti-MAP antibodies (Abs) in the healthy Japanese population, we performed an enzyme-linked immunosorbent assay (ELISA) using a lipophilic antigen that was previously used for the diagnosis of bovine paratuberculosis (Eda et al., 2006; Scott et al., 2010). In addition, the human immunoglobulin G (IgG) subclass profile against the MAP antigens was evaluated. To improve specificity of the ELISA reaction, test sera were pre-adsorbed with M. phlei antigen and heat-killed Bacillus Calmette-Guérin (BCG). This is the first article reporting anti-MAP Abs seroprevalence in the Japanese population.
Materials and Methods
Serum samples
Human serum samples were collected from 126 noncompensated volunteer donors, recruited from Sangenjyaya hospital staff between March and April 2014 during a routine annual health check. Subjects were ethnically all Japanese from at least three generations; 99 were female and 27 were male (F:M ratio=3.7:1). The mean age at the inclusion was 41.9±11.7 years. There was no previous history of tuberculosis infection; however, according to the Tuberculosis Prevention Act law, all Japanese receive the BCG vaccination when they are negative for the tuberculin test in early childhood. Five milliliters of venous blood were collected in K2-EDTA, and after separation by centrifugation sera were stored at −20°C until serological assays. This study, including informed consents, was evaluated and permitted by the research ethics committee of the hospital (Approval No. 2014.3.7) and Tohto College of Health Sciences (Approval No. H2511).
Antigens
Preparation of antigens was performed as described elsewhere (Eda et al., 2006). ATCC 19698 strain of MAP and the BCG strain Tokyo 172 were cultured in Middlebrook 7H9 medium (Difco Laboratories, Detroit, MI) enriched with 10% Middlebrook OADC (Becton Dickinson, Tokyo, Japan) and, in the case of MAP, the medium was complemented with 2 mg/L of mycobactin J (Allied Laboratory, Fayette, MO). The cultures were maintained at 37°C without shaking until they reached an optical density (OD) of approximately 0.7 at 600 nm. MAP and BCG were harvested from liquid culture at stationary phase, which contained approximately 5–10 mg/mL of bacteria, and centrifuged at 2600×g for 10 min. The pellet (1.7 mg/mL) was suspended in 80% ethanol, mixed by vortexing at room temperature for 60 s, and centrifuged at 10,621×g for 10 min. Finally, surface lipophilic antigen was dissolved with absolute ethanol and used in ELISA at the concentration of 0.68 μg/μL.
Absorption of serum sample
Since mycobacteria share a variety of antigens, serum samples have been pre-absorbed with various antigens that are known to be cross-reactive with other mycobacteria before performing ELISA examination (Yokomizo et al., 1983; Bannantine et al., 2008).
M. phlei
M. phlei antigen has been most routinely used to improve the specificity in diagnosis of paratuberculosis as an international standard method (Yokomizo et al., 1983). For our purpose, we used the absorption reagent and M. phlei soluble antigen found in a commercially available ELISA kit, Johnelisa II kit (Kyoritsu Seiyaku Corporation, Tokyo, Japan). This kit has been previously used for the National paratuberculosis control program in Japan (Momotani, 2012a). The absorption was carried out according to the instruction manual of the kit, for 2 h at room temperature.
BCG
All the healthy donors enrolled in the study were BCG vaccinated, and this could cause false-positive reactions (Buddle et al., 2003). To discount the existence of cross-reactivity due to BCG vaccination, all IgG-positive serum samples were also absorbed with BCG antigen overnight at 4°C.
In order to confirm the specificity of MAP antigens, inhibition experiments were performed as follows: all sera and one negative serum were pre-incubated overnight at 4°C with saturating concentration (titrated for each serum) of BCG antigens, then subjected to ELISA on plates coated with MAP antigens. MAP antigen was also added as positive control.
Ethanol vortex indirect ELISA (EVELISA)
EVELISA was carried out similarly to methods reported in previous articles (Eda et al., 2006; Scott et al., 2010). The optimal dilutions of antigens, serum sample, and secondary Ab were determined by check board titration. Then, each well of a 96-well plate (SUMILON ELISA Plate MS-8496F; Sumitomo Bakelite Co., Ltd., Tokyo, Japan) was coated with 50 μL of diluted antigens at room temperature overnight to immobilize ethanol-extracted materials onto the surface of the wells by evaporating the solution. The day after, 300 μL of blocking solution (Dulbecco's phosphate-buffered saline [PBS] pH 7.0 containing 20 vol/vol% Blocking One (Nakalai tesque, Kyoto, Japan) was added to each well and the plate was further incubated at room temperature for 1 h. After rinsing with 100 μL of PBST (10 mM PBS, pH 7.0, containing 0.5% Tween 80), serum samples were added at 1:100 dilutions in PBST for 2 h at room temperature. After the wells were washed 4 times with PBST, horseradish peroxidase (HRP)–labeled-goat anti-human IgG polyclonal antibody (1:2000 dilution), anti-human IgG1-HRP (1:2000), anti-human IgG2-HRP (1:2000), anti-human IgG3-HRP (1:4000), or anti-human IgG4-HRP (1:2000) (all purchased from Southern Biotech, Birmingham, AL) was added for 1 h at room temperature. After washing four times with PBST, 100 mL of a solution of ABTS Peroxidase System (SeraCare Life Sciences, KPL, Gaithersburg, MD) was added to each well and the plates were incubated at room temperature to develop color reactions according to the manufacturer's instructions. The OD was read at a 415-nm wavelength for 20 min using a Bio-Rad iMark microplate absorbance reader (Bio-Rad, Tokyo, Japan).
Conventional ELISA
To compare the results obtained by our in-house EVELISA, we performed an ELISA control by means of a commercially available Johnelisa II kit (Kyoritsu Seiyaku Corporation). The system uses a whole MAP antigen. Since the kit is made for bovine paratuberculosis, instead of the secondary Ab provided by the kit, we used an HRP-labeled goat anti-human IgG polyclonal antibody (Southern Biotech) (1:2000 dilution). Each serum sample was pre-absorbed with soluble antigens of M. phlei, with the purpose of eliminating much of the cross-reacting Ab due to environmental bacteria (Yokomizo et al., 1985). Indeed, this pre-absorption step was shown to be efficient for reducing such false-positive reactions and therefore improving the test's specificity without reducing the sensitivity.
Statistical analysis
The data obtained was analyzed by GraphPad Prism 6.0 software (La Jolla, CA). Data are presented as median with interquartile range, with p values below 0.05 indicating a statistically significant result. The cut-off for positivity (mean plus two times the standard deviation of OD of 415 nm) in each assay was set at 95% specificity and the sensitivity was chosen accordingly. In order to validate the information produced, a correlation analysis between the titer of anti-MAP Abs detected by our EVELISAs and commercial ELISA kit was performed by the Spearman correlation test. Furthermore, agreement between assays was assessed using Bland–Altman analyses. ELISA results were normalized to a strongly positive control serum included in all experiments. Negative control wells were obtained by incubation of immobilized antigens with secondary Ab alone, and their mean values were subtracted from all samples.
Results
EVELISA was able to reveal total IgG and subclass levels IgGs (1–4). At the established cut-off of 0.53, MAP antigens gave strong ELISA titers values in 18 out of 126 (14%; 95% confidence interval [CI]: 9–21%) sera (Fig. 1A). Among the IgG-positive subjects, high levels of IgG1 and IgG4 were observed in 8 out of 18 (44.5%, 95% CI: 24–66%) sera and 10 out of 18 (55.5%, 95% CI: 34–75%) sera, respectively. In contrast, no substantial levels of IgG2 and IgG3 were detected in sera (Fig. 1B).
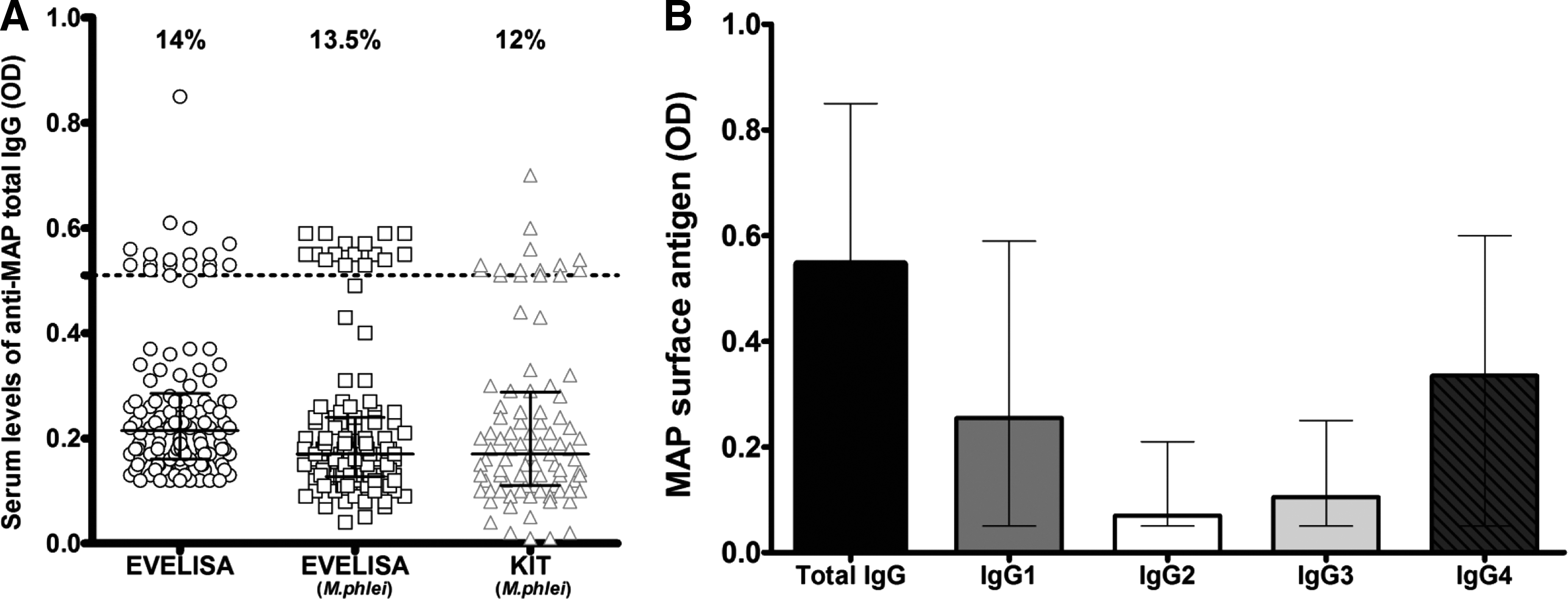
Evaluation of the humoral response to Mycobacterium avium subsp. paratuberculosis (MAP) antigens.
The EVELISA assay displayed good reproducibility. Indeed, a MAP IgG-positive serum with Ab reactivity near the cut-off value was tested in three separate experiments giving an interassay coefficient of variation (CV) of 6.6%. The same serum was tested 20 times in a single experiment, producing an intra-assay CV of 2.3%.
We also performed EVELISA against total IgG but with a pre-absorption step with M. phlei, and the results were almost the same (Fig. 1A). Indeed, Abs were detected in the sera of 17 out of 126 (13.5%; 95% CI: 8–20%) subjects, with a correlation r s=0.7 and p<0.0001 (Fig. 2B). Bland–Altman analysis showed that the 2 methods were in agreement within 95% CI of limits –0.09 to 1.17, with a bias and precision of 0.04±0.06 (Fig. 2D).
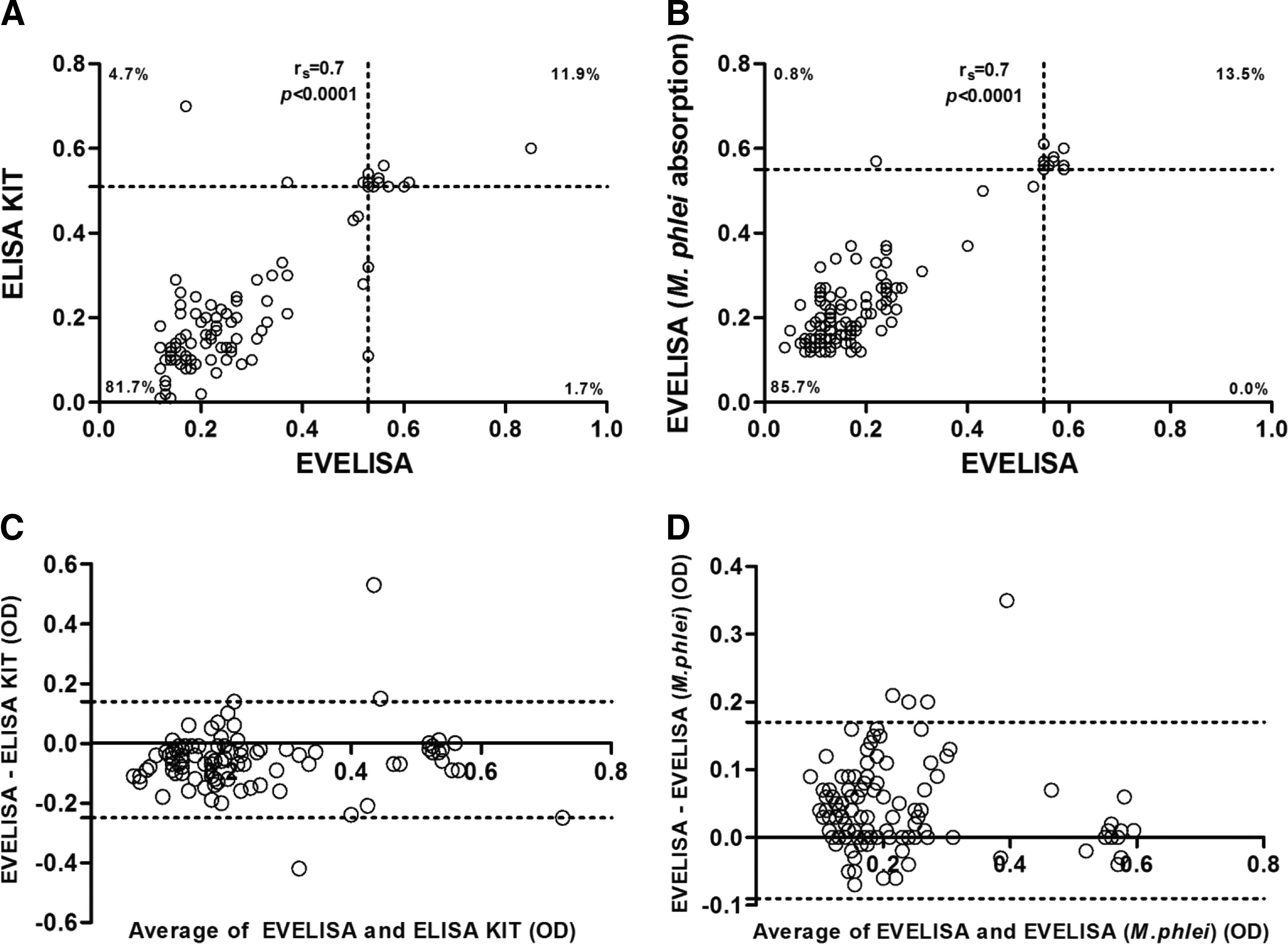
Agreement between enzyme-linked immunoassay (ELISA) methods.
Concerning the commercial ELISA kit, high levels of total IgG were detected in 15 out of 126 (12%; 95% CI: 7–18%) sera (Fig. 1A). There was high degree of correlation when comparing the two methodologies (r s=0.7 and p<0.0001) (Fig. 2A), which suggests that EVELISA is a good technique for MAP detection in humans. The Bland–Altman method showed good agreement between the techniques, with a mean bias of −0.05±0.1 and 95% limits of agreement of −0.25 to 0.14 (Fig. 2C).
Unfortunately, since the diagnostic sensitivity and specificity of the tests using known positive and negative human sera have not been determined, there is no way to know what proportion of the positive tests are true/false positives or how many true positives were missed.
In the inhibition experiments, we confirmed both the data obtained by EVELISA and that anti-MAP and anti-BCG Abs targeting surface lipophilic antigen are not cross-reactive. When pre-adsorbing the sera with BCG antigen, the latter was not capable of blocking the binding of the Abs to the targeted MAP antigens. In fact, BCG antigen caused only a little decline in signal (5–6%), while positive control (MAP antigen) inhibited the MAP seroreactivity in all Ab-positive sera (40–75%) (Fig. 3).
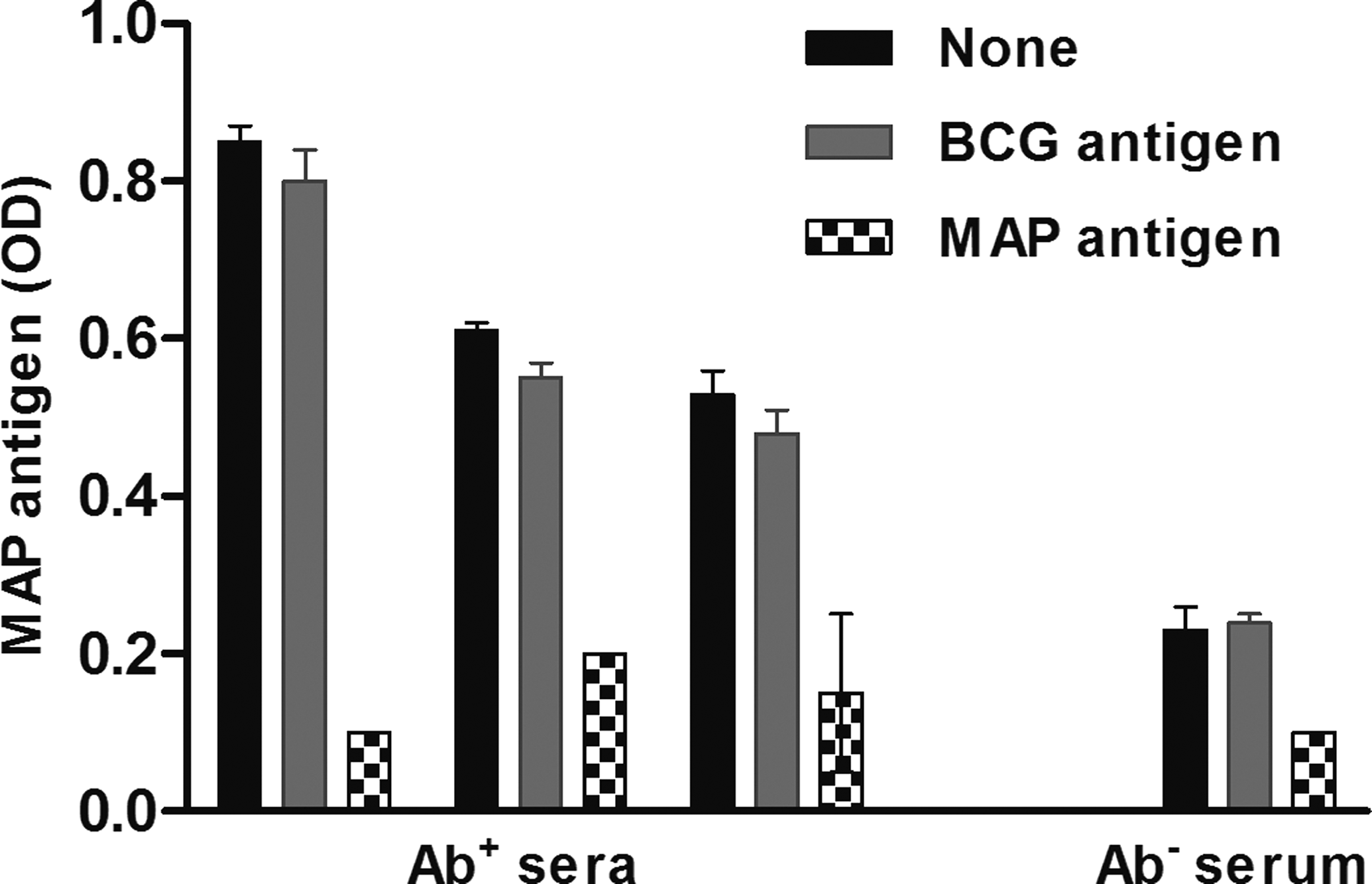
Inhibition assay. Three antibody-positive (Ab+) sera and one antibody-negative (Ab−) serum were pre-incubated overnight with saturating concentrations of Bacillus Calmette-Guérin (BCG) antigen and Mycobacterium avium subsp. paratuberculosis (MAP) antigen. The first bar (none) represents a regularly performed enzyme-linked immunoassay (at 1/100 serum dilution). Results are representative of two separate experiments. OD, optical density.
Discussion
The present study revealed the seroprevalence of anti-MAP Abs in a healthy cohort of potentially MAP-exposed Japanese individuals, which will allow us to understand whether the Japanese population could be considered at risk of becoming exposed to MAP. Indeed, our results showed that 14% of sera were IgG positive. These findings prompt us to investigate the profile of IgG subclasses in serum. MAP antigen was capable of eliciting a strong IgG1 Abs response, detected in the sera of 44.5% of the subjects under scrutiny, confirming the capability of MAP antigens of raising a high Ab level of IgG1 subclass in bovine paratuberculosis (Yokomizo et al., 1983).
EVELISA showed that IgG4 was the most abundant subclass, corresponding to 55.5% of the detected IgG. This is very remarkable new data, since this molecule accounts for less than 5% of the total IgG in healthy persons (Stone et al., 2012). The presence of IgG4 in the sera suggests the occurrence of a prolonged antigenic stimulation, as IgG4 are usually present only when antigenic exposure is chronic (Stone et al., 2012). The concept of “IgG4-related disease” has often been used to understand the pathogenesis of various malignant, infectious, autoimmune, and inflammatory disorders (Kamisawa and Okamoto, 2006), however, there is no previous study documenting a relationship between antigen exposure and elevated levels of IgG4 against the mycobacteria. Our new hypothesis is stated on the assumption that the continuous exposure of human to MAP, mainly due to the continuous consumption of MAP-contaminated dairy products, may be a relevant environmental factor involved in the onset of IgG4-related diseases (can be seen as a background). It is noteworthy that among the genetic susceptibility factors to consider in Japan, both the HLA serotypes DRB1*0405 and DQB1*0401 are known to increase the susceptibility to IgG4-related disease in Japanese populations (Kawa et al., 2002).
The prevalence of paratuberculosis is very high worldwide, with the exception of a few well-controlled countries such as Japan or Sweden (Momotani, 2012a). The shedding of the bacilli into milk (Favila-Humara et al., 2010) and contaminated meat is considered to be the result of limited control measures, such as leaving infected animals in the same herd even after a positive diagnosis of paratuberculosis. Subsequently, shedding levels of MAP into feces and milk increase as the disease progresses (Sharp, 1987). Paratuberculosis incidence is low in Japan, mainly because of the successful national control measures and quarantine under law (Momotani, 2012a), but large amount of dairy products have been imported from Western countries (Momotani, 2012a), with the latter exposing the Japanese population to MAP antigens as reported in the present study.
The present study also demonstrates the value of EVELISA performed with MAP lipophilic antigens (Scott et al., 2010) for detection of human Ab targeting MAP. Of note, this test was proven to be highly sensitive and subspecies-specific for the detection of MAP-infected cattle, indeed the reported diagnostic specificity and sensitivity of the EVELISA were 97.4% and 100%, respectively (Eda et al., 2006). The diagnostic specificity was improved by absorption with other mycobacterial antigen derived from BCG and M. phlei. Indeed, the specificity was not influenced by the preabsorption. The fact that we obtained comparable results when analyzing human samples with both EVELISAs and the commercial ELISA kit seems to confirm what was previously observed when analyzing infected animals (Eda et al., 2006).
Given that until now there are no specific antigenic epitopes for practical MAP detection in humans, EVELISA could be definitely used together with the molecular detection by real-time PCR to detect MAP-infected individuals or to perform a serological screening (Scott et al., 2010).
Conclusions
Our findings suggest the existence of a contamination of MAP in dairy products, and that the Japanese are a population at risk of becoming infected and/or exposed with MAP. Additional in-depth investigation of the mechanism through which MAP might interact with the human immune system by continuous exposure via food is required to prevent the onset or reduce the progress of MAP-related diseases.
Footnotes
Acknowledgments
We gratefully acknowledge Dr. Y. Mori and Y. Shimoji of National Institute of Animal Health for providing heat-killed mycobacterial antigens. This work was supported by Japan Society for the Promotion of Science KAKENHI Grant Number 23240061.
Disclosure Statement
No competing financial interests exist.