
Abstract
The impact of extended-spectrum β-lactamases (ESBLs) and plasmid-mediated AmpC β-lactamases (PMAβs) of animal origin constitutes a public health concern. In this study, 179 Escherichia coli from food animals and products were analyzed, among which, 15 cephalosporin-resistant isolates harboring ESBL (CTX-M-1 [n = 8], CTX-M-14 [n = 1], SHV-12 [n = 2]) or PMAβ [CMY-2, n = 5]) were identified in poultry and swine, from different farms of distinct regions of Portugal. The multiple sequence-type IncI1-driven spread of ESBLs and PMAβs, flanked by widely disseminated mobile elements, was guaranteed by ST26/IncI1-harboring bla SHV-12, ST12/IncI1-harboring bla CMY-2, ST3 and ST38/IncI1-harboring bla CTX-M-1, and ST1/IncI1-harboring bla CTX-M-14. An IS10-disrupted In2-4, presenting a new attI2 recombination site, was also detected in a SHV-12/CTX-M-1-harboring isolate. This study highlights the fact that animals may act as persistent sources of ESBL- and PMAβ-harboring plasmids genes that might be transferred to humans through direct contact or via the food chain.
Introduction
T
The aim of the present study was to investigate the importance of ESBL and PMAβ within a group of E. coli from food animals, and to determine the extent of their association with MGE; in addition, we expect to contribute to the architectural knowledge of a new type of class 2 integron.
Materials and Methods
A collection of 179 E. coli isolates was gathered in a food safety laboratory covering four regions of Portugal (North, Center, Lisbon and Tagus Valley, and South). All E. coli isolates were recovered from routine diagnostic samples (macerated organs [n = 174] and fecal samples [n = 5]), during 1 year, identified by automated methods and sent to the NRL-AR/HAI at the National Institute of Health in Lisbon, without any prior selection criteria. The samples were recovered from three distinct animal origins, as follows: poultry (n = 164), pigs (n = 13), and rabbits (n = 2), and collected as reported elsewhere (Jones-Dias et al., 2013). Antimicrobial susceptibility of all isolates was performed by disk-diffusion method, as previously reported (Bonnet et al., 2013). Isolates nonsusceptible to third-generation cephalosporins were investigated for the presence of ESBL- and PMAβ-encoding genes, transferability of resistance, multilocus sequence typing (MLST), plasmid MLST, integrons, and genetic environment of interest genes, as reported elsewhere (Clemente et al., 2015). Isolates were considered multidrug-resistant (MDR) if they presented nonsusceptibility against three or more structurally unrelated antibiotics (Magiorakos et al., 2012).The content of the class 2 integron from isolate INSLA289 was also assessed through polymerase chain reaction mapping, using a targeting sequencing approach with primers described elsewhere (Ramírez et al., 2005; Sáenz et al., 2010). The new version of integron In2-4 was submitted to the European Nucleotide Archive with accession number LN827615.
Results and Discussion
Overall, among the 179 E. coli isolates recovered, we detected 15 isolates nonsusceptible to third-generation cephalosporins, all of them harboring either ESBL or PMAβ (Table 1). bla CTX-M-1 (n = 8) was the most frequent ESBL-encoding gene, followed by bla SHV-12 (n = 2) and bla CTX-M-14 (n = 1); bla CMY-2 (n = 5) was the only PMAβ identified. More than half of the isolates were also identified as MDR, as shown in Table 1. The presence of the same β-lactamases (TEM-1, CTX-M-1, CMY-2) in animals from different regions points to different sources of these β-lactamases, spread across three important regions of the country (Table 1). This is in agreement with the high genetic diversity noticed: one clonal complex (ST88 and ST23) and eight additional singletons (Table 1). Thus, the spread of ESBL and PMAβ in E. coli from food animals in the country strongly suggests horizontal transfer driven by MGE, rather than a clonal dissemination.
Features that were transferred by conjugation to recipient strains, along with the bla SHV-12, bla CTX-M-1, -14 and bla CMY-2 genes, are underlined.
Farms I, J, and K belonged to the same company and are located in the same county; farms B, C, and F belong to the same company, but are located in different counties.
ND, not determined due to the presence of two ardA genes within the same IncI1 plasmid, which prevent us from assigning a single allele; MLST, multilocus sequence typing; PBRT, polymerase chain reaction–based replicon typing; pMLST, plasmid multilocus sequence typing; LTV, Lisbon and Tagus Valley region; NA, not applicable because no transconjugant was obtained from INSLA290; P, penicillins (amoxicillin); I, β-lactam/β-lactamase inhibitor combinations (amoxicillin plus clavulanic acid, piperacillin plus tazobactam, cefotaxime plus clavulanic acid); C1G, first-generation cephalosporins (cephalotin); C2G, second-generation cephalosporins (cefuroxime); C3G, third-generation cephalosporins (ceftiofur, ceftriaxone, cefotaxime, ceftazidime); C4G, fourth-generation cephalosporins (cefepime); M, monobactam (aztreonam); F, cephamycines (cefoxitin); C, carbapenems (imipenem, meropenem); Q, quinolones (nalidixic acid, norfloxacin, pefloxacin, ciprofloxacin, flumequine, marbofloxacin, enrofloxacin); TE, tetracyclines (tetracycline); A, aminoglycosides (kanamycin, gentamicin); FE, phenicols (chloramphenicol); S, sulphonamides (trimethoprim, trimethoprim plus sulfamethoxazole); FT, nitrofurans (nitrofurantoin).
All the transconjugants carrying ESBL- or PMAβ-encoding genes harbored IncI1 (14/15), and three of them also carried IncFIB (3/13) plasmids (Table 1). However, E. coli isolates harbored plasmids belonging to different replicon types: IncI1 (n = 15), IncFIA (n = 1), IncFIB (n = 13), and IncFIC (n = 7) (Table 1). In fact, the mobilization of the detected bla genes was driven by five different IncI1 plasmids: ST26/IncI1-harboring bla SHV-12, ST12/IncI1-harboring bla CMY-2, ST3 and ST38/IncI1-harboring bla CTX-M-1, and ST1/IncI1-harboring bla CTX-M-14. bla CTX-M-1 was associated with two different plasmids from clonal complex 3, suggesting the occurrence of intraplasmid evolution. ST12/IncI1 and ST3/IncI1 have been associated with the spread of CTX-M-1 and CMY-2 β-lactamases in animals, which represents a concern, because they might be easily disseminated to humans, water, and soil (Ben Sallem et al., 2014). Globally, although the isolates carried other plasmids, IncI1 plasmids seem to be responsible for this epidemic-like spread, which has already been cause for concern in other countries (de Been et al., 2014).
The characterization of the genetic context of ESBL- and PMAβ-encoding genes showed the presence of ISEcp1 upstream of the nine bla CTX-M-1 and of the five bla CMY-2 genes, while the two bla SHV-12 genes were flanked upstream by IS26. The downstream genetic context of the genes consisted of orf477 in bla CTX-M-1 genes (n = 8), IS903 in bla SHV-12 and a blc-sugE structure in bla CMY-2 (Table 1). The detection of those IS elements, despite the plasmid background or ST, indicates that they might play an important part in the capture, expression, and mobilization of bla ESBL and bla PMAβ genes (Stokes et al., 2011; de Been et al., 2014).
Furthermore, five isolates were positive for class 1 integrons, and the isolate INSLA289 for a class 2 integron; the array for the latter revealed a new arrangement (Table 1). The integron sequence resembled the classic In2-4, except within the recombination site where the array containing dfrA1, sat2, and aadA1y was preceded by a complete IS10 that resulted in the disruption of the original attI2 and reconstruction of a second attI2 immediately after the inserted IS10 (Fig. 1) (Ramírez et al., 2005, 2010). Consequently, integrating abilities of In2 remained intact, and the new construct benefits from an additional capability to move using the recently acquired IS10 transposase, which may have important consequences on resistance dissemination. Moreover, an amino acid alteration (H379Y) was detected within TnsD protein.
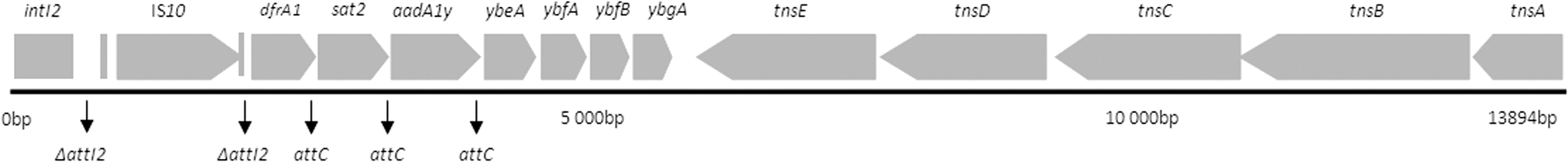
Scheme of the In2-4 harbored by isolate INSRA289 (LN827615) where the attI2 recombination site (793-809bp and 2148-2193bp) is disrupted by the insertion of a complete IS10 (1329bp, at position 810-2138bp).
This study reports multiple animal sources of ST3/IncI1-harboring bla CTX-M-1, and other ESBL/PMAβ-encoding genes associated with the spread of epidemic plasmids within and among different farms. These genes have been increasingly identified in humans and animals across Europe, highlighting the widespread distribution of those MGE along the food chain.
Footnotes
Acknowledgments
This work was partially presented at the 23rd European Congress of Clinical Microbiology and Infectious Diseases in Berlin, Germany. This work was supported by Fundação para a Ciência e a Tecnologia (FCT) for grant number PTDC/CVT/65713. We also thank the FCT for Project grant PEst-OE/AGR/UI0211/2011-2014, Strategic Project UI211-2011-2014. D. Jones-Dias and V. Manageiro have received research funding from FCT (grant numbers SFRH/BD/80001/2011 and SFRH/BPD/77486/2011).
Disclosure Statement
No competing financial interests exist.