
Abstract
The purposes of this study were to investigate the antimicrobial resistance of Aeromonas veronii biovar sobria isolated from gilthead sea bream and to characterize the virulence-implicated genes. Fish samples (n=365) were collected from wholesale and retail markets in Aljouf, Saudi Arabia between 2013 and 2014. A total of 45 A. veronii biovar sobria isolates (12.3%) from those samples were tested for resistance to a range of antimicrobial agents. All strains exhibited 100% resistances to nalidixic acid, carbenicillin, cephalothin, erythromycin, kanamycin, tetracycline, and trimethoprim–sulfamethoxazole. Additionally, the highest susceptibility encountered was to ciprofloxacin (100%). In the present study, we examined the presence of several genes, including aerolysin, elastase, lipase, flagellin, enterotoxin, and DNases, that code for putative virulence factors that may play important roles in bacterial infection. It was found that all of these genes were common in these strains. Several strains isolated from diseased gilthead sea bream were tested for virulence in gilthead sea bream by intraperitoneal injections. The median lethal dose values ranged from 5×103 to 5.2×109 colony-forming units per fish. These data suggest that commercial gilthead sea bream fish may act as the reservoir for multiresistant A. veronii biovar sobria and facilitate the dissemination of virulence genes.
Introduction
A
The gilthead sea bream (Sparus aurata L.) is a marine fish with high economic value for the aquaculture industry. Several pathogenic microorganisms are involved in outbreaks in these fish (Zorrilla et al., 2003). Little information about the microbiological contamination of gilthead sea bream fish is available. Therefore, studies on fish microbiology are necessary for the management of aquaculture and public health.
Moreover, because little is known about the putative virulence genes involved in A. veronii biovar sobria infection, we decided to evaluate the public health significance of representative A. veronii biovar sobria isolates that were recovered from gilthead sea bream fish and investigate whether these isolates contained putative virulence (aerolysin, elastase, lipase, hydrolipase, flagellin, enterotoxin, and DNases genes) factors associated with antimicrobial resistance.
Materials and Methods
Fish preparation and conditions for the isolation of Aeromonas
Gilthead sea bream were purchased from fish markets in the Aljouf province of Saudi Arabia. Gilthead sea bream were maintained at room temperature (28°C) in sterile water. Twenty-five grams of fish flesh were weighed aseptically and homogenized for 2 min in stomacher bags containing 225 mL of alkaline peptone water (CM1028; Oxoid, UK). After 18 h of incubation at 37°C, an aliquot of the enrichment was inoculated into blood agar containing 30 mg/L ampicillin and incubated at 28°C for 24 h (Yano et al., 2015). Forty-five presumptive A. veronii biovar sobria colonies were selected for further identification. All isolates were stored in Luria-Bertani (61748 LB Broth; Sigma-Aldrich, US) broth containing 20% glycerol at −70°C. Consents and all ethical relevant procedures were designed according to Aljouf University bioethics committee guidelines.
Phenotypic identification
Before each test, all cultures were grown on tryptose soya agar (CM0131; Oxoid) at 37°C for 18 h. All Aeromonas spp. were identified biochemically to the species level using the following 14 tests that were chosen from those described by Altwegg (1999): the indole, gas from glucose, arginine dihydrolase, lysine decarboxylase, and ornithine decarboxylase by the Moeller's method, esculin hydrolysis, Voges–Proskauer, acid production from
Molecular identification
All strains were re-identified based on restriction fragment length polymorphism patterns that were obtained from the 16S rDNA following the method described by Borrell et al. (1997).
Antimicrobial susceptibility
The resistances of all strains to different antimicrobial agents were determined with the disk-diffusion method as previously described (CLSI, 2012). The antibiotics and concentration ranges tested were as follows: amikacin (30 μg), nalidixic acid (30 μg), carbenicillin (100 μg), cephalothin (30 μg), cefotaxime (30 μg), ciprofloxacin (5 μg), chloramphenicol (30 μg), erythromycin (15 μg), gentamicin (10 μg), kanamycin (30 μg), rifampicin (5 μg), tetracycline (30 μg), and trimethoprim–sulfamethoxazole (25 μg). The resistance breakpoints defined by the Clinical and Laboratory Standards Institute (CLSI, 2012) for A. veronii (ATCC 35626) were used as controls.
Genetic detection of virulence genes
Nucleic acid isolation
Nucleic acids were isolated via lysis of the bacteria in a solution containing 0.5% sodium dodecyl sulfate and 5 mg lysozyme per milliliter followed by extraction with phenol-chloroform. Nucleic acid samples were precipitated with ethanol and dissolved in TE buffer (10 mM Tris chloride, 1 mM EDTA [pH 8.0]). The nucleic acid content was quantified by determining the optical density at 260 nm and adjusted to produce a final concentration of 200 μg/mL in TE buffer. Template DNA for polymerase chain reaction (PCR) was prepared by further dilution in distilled H2O to a concentration of 2 μg/mL (Sambrook and Russel, 2009).
Detection of virulence genes
The identified A. veronii isolates were streaked onto nutrient agar plates and incubated at 36°C for 24 h. Single distinct colonies were suspended in nuclease-free water, vortexed to create a suspension, and boiled at 100°C for 10 min. Thereafter, the suspension was centrifuged at 12,000×g for 5 min, and the supernatant was transferred to a fresh sterile 2-mL Eppendorf tube and immediately placed on ice and used for template DNA. A. veronii ATCC 35626 was used as a positive control, and nuclease-free water was used as a negative control for all PCR reactions. The oligonucleotide primers used for the amplification of the virulence genes are shown in Table 1. These primers were selected based on their specificities as confirmed by the previous studies indicated in Table 1. PCRs were carried out in total volumes of 25 μL. The PCR conditions for the detection of the virulence genes are presented in Table 1 (Nawaz et al., 2010). The subsequent electrophoresis was performed using 1% (wt/vol) agarose gel in 0.5 X Tris-borate-EDTA buffer and a voltage of 100 V that was applied for 45 min and ethidium bromide (E8751; Sigma-Aldrich, USA) staining. A molecular weight marker of 100 bp was used for the gene DNA ladder. After electrophoresis, gel imaging was performed using the Molecular Imager® Gel Doc™ XR+System with Image Lab™ Software 170-8195 (BioRad, USA).
Infectivity tests
To confirm the pathogenicity of A. veronii biovar sobria, experimental challenges were conducted using healthy gilthead sea bream and sea bass (mean weight: 10 g). The 50% lethal dose (LD50) tests were performed in batches of 6 fish per dose via intraperitoneal injections. Five strains (the molecular types that were the most predominant) were selected, and the virulences of these types for gilthead sea bream and sea bass were tested (Table 2). Overnight cultures of the bacterial strains to be tested were washed by centrifugation and suspended in phosphate-buffered saline (PBS, pH 7). The gilthead sea bream and sea bass were injected with 0.1 mL of serial 10-fold dilutions of the culture (103 to 108 colony-forming units (CFU)/mL). The control fish were inoculated with PBS only. The experimental infections were performed in aquaria at 35% salinity and 22°C. Mortalities were recorded daily for 7 days, and were attributed to the inoculated bacterium when that bacterium was recovered in pure cultures from the internal organs (i.e., the liver and kidney). The LD50 was calculated with the highest-numbers method of Reed and Müench (1938).
Injected by phosphate-buffered saline (pH 7) only.
CFU, colony-forming units; LS50, median lethal dose.
Results and Discussion
Incidence of A. veronii biovar sobria in the fish samples
A total of 45 (12.3%) A. veronii biovar sobria isolates were recovered from 365 fish and seafood samples. The susceptibilities of the 45 isolates of A. veronii biovar sobria to 13 antimicrobial agents were evaluated. All strains exhibited 100% resistance to nalidixic acid, carbenicillin, cephalothin, erythromycin, kanamycin, tetracycline, and trimethoprim–sulfamethoxazole. Additionally, the highest susceptibility encountered was that to ciprofloxacin (100%) followed by amikacin (89%), chloramphenicol (80%), rifampicin (33), gentamicin (29%) and cefotaxime (27%) (Fig. 1).
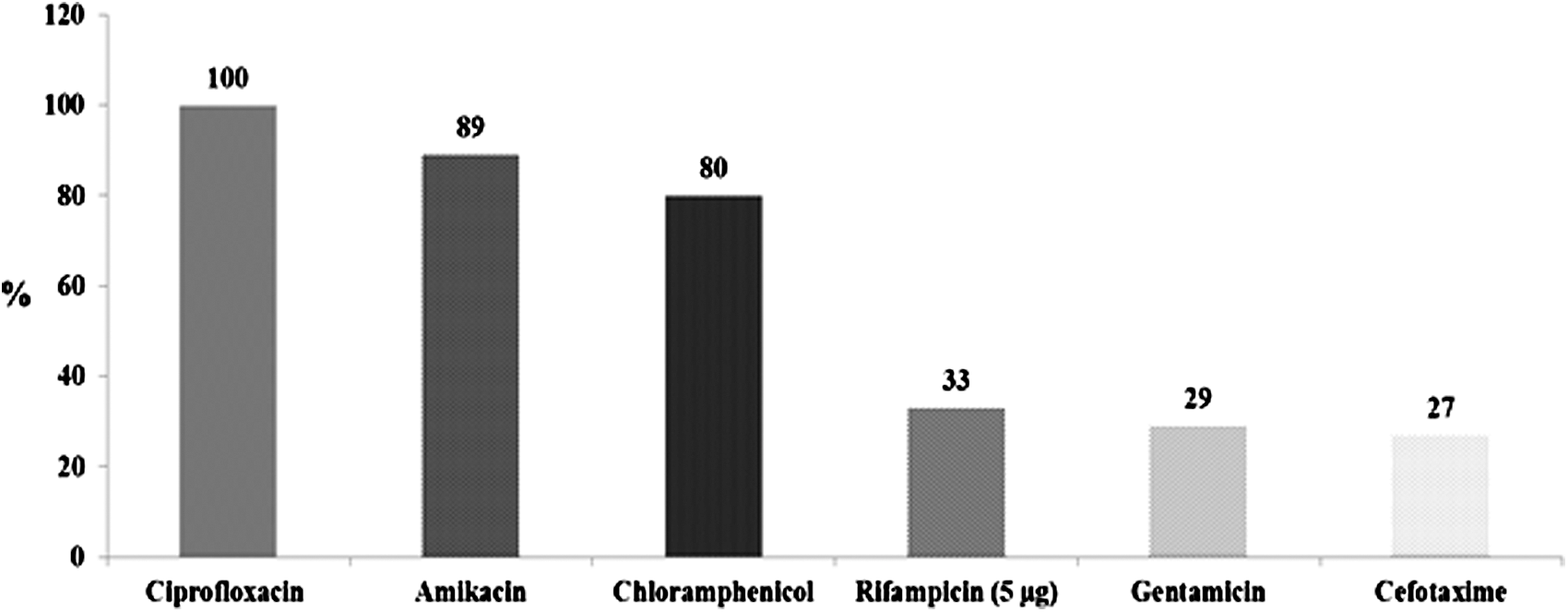
Percentage of antimicrobial sensitivity patterns of Aeromonas veronii biovar sobria strains that were isolated from the gilthead sea bream (Sparus aurata L.)
Gilt-head sea bream (S. aurata L.) are the most important marine fish that are cultured in Mediterranean countries, but description of the pathologies that affect this species remain scarce. In the present work, we performed a survey study of the A. veronii biovar sobria pathologies that affect this fish species. A previous study of marine fish (Zorrilla et al., 2003) found that the most prevalent infectious pathologies that affected the farmed gilthead sea bream in the southwest of Spain were of bacterial origin and that Gram-negative bacteria were the most frequently isolated (>93%). In agreement with this study, we found that the 45 strains of A. veronii biovar sobria that were isolated were the predominant bacteria in the gilthead sea bream.
In the present study, multiresistance to antimicrobials agents was examined. We found that all of the tested strains were resistant to nalidixic acid, carbenicillin, cephalothin, erythromycin, kanamycin, tetracycline, and trimethoprim–sulfamethoxazole. However, tetracyclines were among the therapeutic agents and are most commonly used in human and veterinary treatments. The widespread use of tetracycline has provoked the dissemination of resistance to tetracycline to many marine bacteria (Furushita et al., 2003). Because the products of aquaculture are consumed by humans and because many determinants of antibiotic resistance are encoded by transferable plasmids, cultured fish may serve as a vehicle for the transmission of antibiotic resistance to bacteria that are commensal or pathogenic to humans (Rhodes et al., 2000).
Prevalence of virulence genes in A. veronii biovar sobria
The frequencies of the virulence factors investigated in the 45 A. veronii biovar sobria isolates are shown in Table 3. The gene that was most frequently found in the isolates was the elastase gene (87%), and the least frequently found genes were those for DNases (20%). Of the 45 isolates that were analyzed, 39 (87%) had at least 1 of the genes for the studied virulence factors. In total, 12 distinct patterns of genes for virulence factors were found in the A. veronii biovar sobria isolates. The combination of virulence factors that was most frequently observed was aerolysin/elastase, which was identified in 14 isolates. The aerolysin/elastase/lipase/flagellin/enterotoxin/DNases combination was found in 2 isolates. Notably, 29% of the isolates (13/45) had at least 3 of the evaluated virulence factors.
Aeromonas are considered potential foodborne pathogens when they carry the aerolysin and hemolysin genes (Cumberbatch et al., 1993; Gosling, 1996). Gonzalez-Serrano et al. (2002) and Pollard et al. (1990) failed to detect the aero gene in any of their A. veronii isolates. In contrast, our results indicated that 31% of the A. veronii biovar sobria isolates from the gilthead sea bream harbored the aero gene and the enterotoxin act gene. Aerolysin and enterotoxin play crucial roles in the establishment of infections (Chopra et al., 1996; Soler et al., 2002; Sha et al., 2002). These toxins trigger fluid accumulation in animal intestinal loops, the elongation of CHO cells, the rounding of Y1 adrenal tumor cells, and increases in intracellular cAMP levels (Chopra and Houston, 1999; Gosling, 1996). These toxins exhibit hemolytic, cytotoxic, and enterotoxic activities. Our results indicate that the A. veronii biovar sobria strains isolated from the gilt-head sea bream were virulent strains because they contained both the aero and act genes (4%).
The three genes involved in the regulation and secretion of extracellular serine protease and lipases play a coherent, integrated role in the establishment of the pathogenicity of the Aeromonas spp. (Pemberton et al., 1997). These extracellular enzymes are secreted into the environment via the well-characterized type II or general secretory pathways (Pemberton et al., 1997). Additionally, serine protease has been associated with the activation of aerolysin (aerA) and other extracellular enzymes and thus affects the overall virulence of aeromonads (Nawaz et al., 2010). Aeromonads secrete four different types of extracellular lipases (i.e., lip, lipH3, pla, and plc) that actively participate in the alteration of the host plasma membrane and thus increase the severity of infection (Pemberton et al., 1997). Swarming motility is a flagellum-dependent behavior and is crucial for biofilm formation and bacterial virulence. The polar, monotrichous flagella of the Aeromonas spp. aid in motility and the colonization of surfaces after initial attachment, and aeromonads with lateral flagella are frequently associated with severe dysenteric infections (Kirov et al., 2002; Nawaz et al., 2010).
Infectivity of A. veronii biovar sobria in gilthead sea bream
The results of the virulence assays of the five selected A. veronii biovar sobria strains are shown in Table 2. These assays demonstrated that all of the tested strains were pathogenic for gilthead sea bream. The LD50s ranged from 5×103 to 5.2×109 CFU/fish. Experimentally, the infected fish exhibited external signs similar to those that are observed in outbreaks, including hemorrhagic fins and ulcers. Mortalities were initially observed on the first day and continued to the seventh day postchallenge, and no mortalities were detected in the control fish. Pure cultures of the inoculated strains were reisolated from the livers and kidneys of the moribund fish.
A. veronii biovar sobria was isolated in pure cultures from the internal organs of the diseased fish. Thus, the LD50 for this species was calculated. All of the isolates tested were virulent for the gilthead sea bream. Based on the criteria of Santos et al. (1988), the examined strains can be considered virulent. The LD50 values we observed agree with those reported for this fish pathogen by other authors (Zorrilla et al., 2003). Therefore, specific virulence tests for gilthead sea bream are needed to unequivocally confirm the pathogenic nature of this species. Experimental infections using gilthead sea bream have demonstrated the presence of cytotoxic and neurotoxic effects in fish and in cell lines (Balebona et al., 1998b). The research groups that published the latter study also demonstrated that this strain can obtain iron from high-affinity compounds via siderophores. Moreover, these authors showed that this strain has the ability to adhere to the skin, gills, and intestinal mucosa of sea bream. The extracellular products of this strain are able to degrade sea bream mucus and are cytotoxic for fish cell lines.
Conclusions
In conclusion, in the present work, we characterized the A. veronii biovar sobria strains isolated from gilthead sea bream that were purchased in Saudi Arabia. We found general increases in the resistances to different antimicrobials among the different microorganisms that were isolated in this study, and these findings should be considered to ensure the more rational use of antimicrobials and to execute programs of vaccination against these pathogenic microorganisms. Virulence factors were widely distributed among A. veronii biovar sobria isolates, which indicates the pathogenic potential of these bacteria for aquatic organisms. Aerolysin was the most frequently found virulence factor in the isolates that were analyzed. The present work is the first description of these pathogens affecting cultured gilthead sea bream fish. To assess the virulence of the isolates, the lethal doses for gilthead sea bream were identified, and the results indicate that the isolates were virulent to the fish. Based on the results of this study, we suggest that this bacterial species should be considered an opportunist pathogen for gilthead sea bream and included in future strategies for the treatment and prophylaxis of farmed gilthead sea bream. However, gilthead sea bream serve as a reservoir of A. veronii containing putative multiple virulence genes and multiple antibiotic resistance genes. Thus, the potential health risk due to the consumption of improperly cooked gilthead sea bream harboring A. veronii should not be underestimated. Although adequate cooking should eliminate pathogenic bacteria, undercooking or cross-contamination during preparation could be of concern.
Footnotes
Acknowledgment
This work was funded by the Deanship of Scientific Research (DSR), King Abdulaziz University, Jeddah under grant no. (130-012-D1434). Therefore, the authors acknowledge and thank the DSR for technical and financial support.
Disclosure Statement
No competing financial interests exist.