
Abstract
This study evaluated antimicrobial resistance and virulence factors in Salmonella enterica isolated from a turkey flock in which the birds were raised in an environment where antimicrobials were not administered to the birds, either through feed or water. Salmonella was isolated from turkeys and various environmental samples in the facility using conventional microbiological procedures. Isolates were serotyped and analyzed phenotypically by antimicrobial resistance profiling and genotypically by pulsed-field gel electrophoresis (PFGE) fingerprinting, integron analysis, plasmid profiling, replicon-based incompatibility (Inc) group typing, and virulence gene profiling. Ninety-five S. enterica isolates were isolated from cecal contents (n = 29), feed (n = 22), leftover feed (n = 13), litter (n = 12), drinkers (n = 10), environment (n = 8), and an insect. The following serotypes were identified: Montevideo (24%), Anatum (22%), Agona (17%), Kentucky and Worthington (12%), Senftenberg (11%), and rough phenotypes (3%). The majority of isolates (61/95; 64%) were susceptible to 12 antimicrobials tested; however, despite the absence of antimicrobials in the facility, approximately 36% of the isolates were resistant to two to five antimicrobials. Class 1 integrons were detected in 8% of the isolates. The integron sequence analysis revealed dihydrofolate reductase (dhfr) and aminoglycoside adenylyl transferase (aadA2) genes, which encode trimethoprim and streptomycin resistance, respectively. Furthermore, 71% of the isolates had at least one plasmid. There were five plasmid replicon types identified among the isolates, including IncI1, IncHI2, IncFIIA, IncB/O, and IncP, with variable prevalence among the serotypes. All 95 isolates tested polymerase chain reaction–positive for 19 virulence genes and negative for virD4 and virB4. The virulence gene profiles were similar within the isolates from the same serotype. Within particular serotypes, PFGE patterns revealed 100% similarity, even when the bacterial strains were isolated from different sources, indicating cross-colonization of sources within the turkey facility. On this antibiotic-free turkey farm, turkeys and feed appeared to be the major reservoirs of multidrug-resistant Salmonella, which harbored multiple virulence genes.
Introduction
N
Although considerable research has been conducted to describe the molecular diversity of Salmonella in chickens (Cormican et al., 2002; Foley et al., 2013), limited information is available on the colonization and the mechanisms of Salmonella transmission in turkeys (Nayak and Kenney, 2002; Nayak et al., 2004; Nayak and Stewart-King, 2008). The turkey production cycle ranges from 20 to 22 weeks compared to 6 weeks for chickens. The longer production cycle likely results in different Salmonella dynamics in turkey compared to broiler facilities (Scupham, 2007). There is a continued need to evaluate how on-farm practices affect the level of colonization in turkeys before slaughter.
Antibiotic use in poultry production has been implicated in the emergence and spread of antibiotic-resistant Salmonella (Marshall and Levy, 2011). Multidrug-resistant Salmonella has become a major public health threat worldwide (CDC, 2013; Lai et al., 2013). The percentage of multidrug-resistant Salmonella isolates recovered from organic poultry facilities is significantly lower than that from conventional facilities (Alali et al., 2010; Sapkota et al., 2014). In order to limit risks associated with antibiotic-resistant Salmonella as well as foodborne salmonellosis, efforts should also be in place to implement effective controls on the farms that can reduce and/or eliminate the incidence of Salmonella transmission in the production environment before the birds enter the processing plants. This study examines the antimicrobial, virulence, and plasmid characteristics of Salmonella serovars isolated from a turkey flock. Since the turkeys were raised without antimicrobials, it is valuable to determine whether antimicrobial resistance remains a concern among Salmonella isolated from this production scenario.
Materials and Methods
Bacterial isolates
This study was part of a collaborative effort between West Virginia University and British United Turkeys of America (BUTA), in which seven consecutive flocks were studied in a production facility. The facility was not populated with poultry for approximately 20 years prior to flock placements. Data from six flocks have been published (Hoover et al., 1997; Nayak et al., 2002, 2004; Nayak and Stewart-King, 2008). This article highlights the genotypic and phenotypic characterization of 95 Salmonella strains, representing multiple serovars that were isolated from the previously unreported flock. Prior to flock placement, day-old poults were sampled and environmental swabs were collected from random locations in the facility to obtain baseline data on initial Salmonella contamination. Appropriate farm biosecurity measures and turkey-house cleaning and disinfecting methods were used between flock placements (Nayak, 2000). During the study, samples for Salmonella isolation were taken from turkey cecal contents, litter, drinkers, feed, leftover feed, air samples, and environmental swabs at weeks 7, 13, and 18 of the production cycle. Turkeys in this study received two types of proprietary diets administered by BUTA reported to be free of antibacterial agents. A general description of sampling, Salmonella isolation, and serotyping procedures has been previously described (Hoover et al., 1997; Nayak et al., 2000).
Evaluation of antimicrobial resistance profiles in Salmonella serovars
Antibiotic susceptibility testing was performed using a NARMS CMV1AGNF plate (Trek Diagnostics, Cleveland, OH). Twelve antimicrobials were tested: amoxicillin/clavulanic acid (AUG; 1/0.5–32/16 μg/mL), ceftriaxone (AXO; 0.25–64 μg/mL), ceftiofur (TIO; 0.12–8 μg/mL), chloramphenicol (CML; 2–32 μg/mL), ciprofloxacin (CIP; 0.015–4 μg/mL), gentamicin (GEN; 0.25–16 μg/mL), kanamycin (KAN; 8–64 μg/mL), nalidixic acid (NAL; 0.5–32 μg/mL), streptomycin (STR; 32–64 μg/mL), sulfisoxazole (SUL; 16–256 μg/mL), tetracycline (TET 4–32 μg/mL), and trimethoprim/sulfamethoxazole (SMX; 0.12/2.38–4/76 μg/mL). Pseudomonas aeruginosa ATCC 27853, Escherichia coli ATCC 25922, and Staphylococcus aureus ATCC 29213 were used as quality-control strains. The results were interpreted according to Clinical and Laboratory Standards Institute (CLSI) guidelines (CLSI, 2002a, b).
Integron analysis
Class 1 integrons were detected by polymerase chain reaction (PCR) (Melendez et al., 2010) and sequenced to determine the antimicrobial resistance genes carried in the integrons (Lynne et al., 2009). Sequences were compared, using BLAST, to National Center for Biotechnology Information's GenBank to identify genes present in the positive isolates.
Pulsed-field gel electrophoresis (PFGE) analysis
XbaI-PFGE was used for fingerprinting Salmonella strains using the PulseNet protocol (Ribot et al., 2006). PFGE patterns for Salmonella were analyzed using BioNumerics software, version 6.0 (Applied Maths, Sint-Martens-Latem, Belgium). Banding pattern similarity was calculated using Dice coefficients and a dendrogram was generated using the unweighted-pair group method arithmetic averages algorithm. Positional tolerance and optimization shift of 2% were allowed for analyses.
Plasmid analysis
Bacterial cells were grown overnight at 37°C in 2–5 mL of Luria–Bertani broth. Cells were harvested by centrifugation and lysed; plasmid DNA was extracted using a commercial kit (Qiagen, Valencia, CA). Plasmid DNA was electrophoresed in 0.8% agarose gels with 1X Tris–borate–EDTA buffer. The BAC-Tracker™ Supercoiled DNA Ladder (Epicentre, Madison, WI) was used as a molecular weight marker.
Incompatibility (Inc) typing
DNA was isolated from each isolate using the Wizard Genomic DNA Purification System (Promega, Madison, WI). PCR-based replicon typing was conducted as described by Johnson et al. (2007), using appropriate positive control strains (Carattoli et al., 2005).
Detection of virulence genes
Thirty-five virulence genes were detected by PCR (Han et al., 2013). PCR products were separated in 96-well 2% E-gels (Invitrogen, Carlsbad, CA), visualized and analyzed using Principal Components Analysis (PCA) (David et al., 2010).
Results
Distribution of Salmonella isolated from different sources in turkey facility
Ninety-five isolates were collected from turkey ceca (n = 29), feed (n = 22), leftover feed (n = 13), litter (n = 12), drinkers (n = 10), environmental samples (n = 8), and an insect. Salmonella were undetected from air samples. Salmonella serotypes Kentucky, Agona, Montevideo, Worthington, Senftenberg, Anatum, and a rough phenotype were identified from the samples. Salmonella Montevideo was the most commonly detected serotype (24%; 23/95), followed by Salmonella Anatum (22%; 21/95), and Salmonella Agona (17%; 16/95) (Table 1). Salmonella was undetected in any of the baseline samples, including day-old poults, incoming feed, water, bedding, environment swabs, and air samples that were collected prior to placement of this flock.
Antibiotic susceptibility
Sixty-one isolates (64%) were susceptible to all the tested antimicrobials. Antimicrobial resistance was observed only in serotypes Kentucky, Worthington, and Anatum isolates (Fig. 1). Thirty-four isolates (36%) were resistant to two or more antimicrobials that included SUL, TET, SMX, GEN, and/or STR (Fig. 1). Resistance to SUL was observed at the highest rate among the tested isolates (36%; 34/95), followed by GEN (26%; 25/95), STR (18%; 17/95), TET (12%; 11/95), and SMX (11%; 10/95).

Dendrogram showing the pulsed-field gel electrophoresis (PFGE) patterns for XbaI restricted turkey-associated Salmonella isolates from different sources in a turkey facility. Similarity was calculated using the Dice coefficient, and clustering analysis was performed by the unweighted pair-group method with arithmetic averages (optimization, 2% and position tolerance, 2%). Clustering cut-off was 85% similarity. I–VI indicates the clusters numbers. TET, tetracycline; GEN, gentamicin; STR, streptomycin; SUL, sulfisoxazole; SMX, trimethoprim and sulfamethoxazole.

Integron analysis
Class 1 integrons were detected in only 8% (8/95) of the isolates, including seven from Salmonella Kentucky. All Salmonella Kentucky integron amplicons were 1.6 kb, whereas the single Salmonella Anatum integron was 1 kb. Sequence analysis of all integrons from the Salmonella Kentucky isolates identified dihydrofolate reductase (dhfr) and aminoglycoside adenylyl transferase (aadA2) genes that encode trimethoprim and streptomycin resistance, respectively, whereas only aadA2 was identified in Salmonella Anatum.
Genotyping (PFGE)
Six primary PFGE clusters were identified in which isolates shared >80% similarity (Fig. 1). Each individual cluster contained isolates largely from the same serotype, regardless of source. All Salmonella Kentucky and two rough isolates were located in cluster I that possessed four different profile patterns. Cluster II contained all Salmonella Agona isolates represented by two different patterns. All Salmonella Montevideo isolates and one rough isolate formed cluster III that included four different patterns. Clusters IV and V contained Salmonella Worthington and Senftenberg isolates, respectively. There were two unique genotypic profiles identified in clusters IV and V. Finally, all Salmonella Anatum isolates formed cluster VI that contained six different profiles. Isolates from different sources in the production facility often possessed the same fingerprints. For example, one fingerprint in cluster II included isolates from ceca, litter, and drinkers. Fingerprints in cluster IV included isolates from turkey ceca, feed, litter, drinkers, and leftover feed, while in cluster VI, isolates from ceca, feed, drinkers, and litter shared a common fingerprint (Fig. 1).
Plasmid analysis and Inc typing
Sixty-seven (71%) isolates had at least one plasmid identified (Fig. 1). Only 13% (3/23) of Salmonella Montevideo and 2 of the 3 rough isolates were positive for plasmids, whereas in each of the other serotypes, at least 81% of the isolates carried at least 1 plasmid. There were five Inc-types identified among the isolates. IncI1 plasmids predominated among the Salmonella Anatum isolates; all but 1 of these isolates (n = 20/21) carried the plasmid. Conversely, among Salmonella Kentucky isolates, 8 of 11 contained at least an IncHI2 plasmid, with 2 also carrying IncI1 plasmids. Other Inc-groups identified among isolates included IncFIIA, IncB/O, and IncP.
Virulence genes
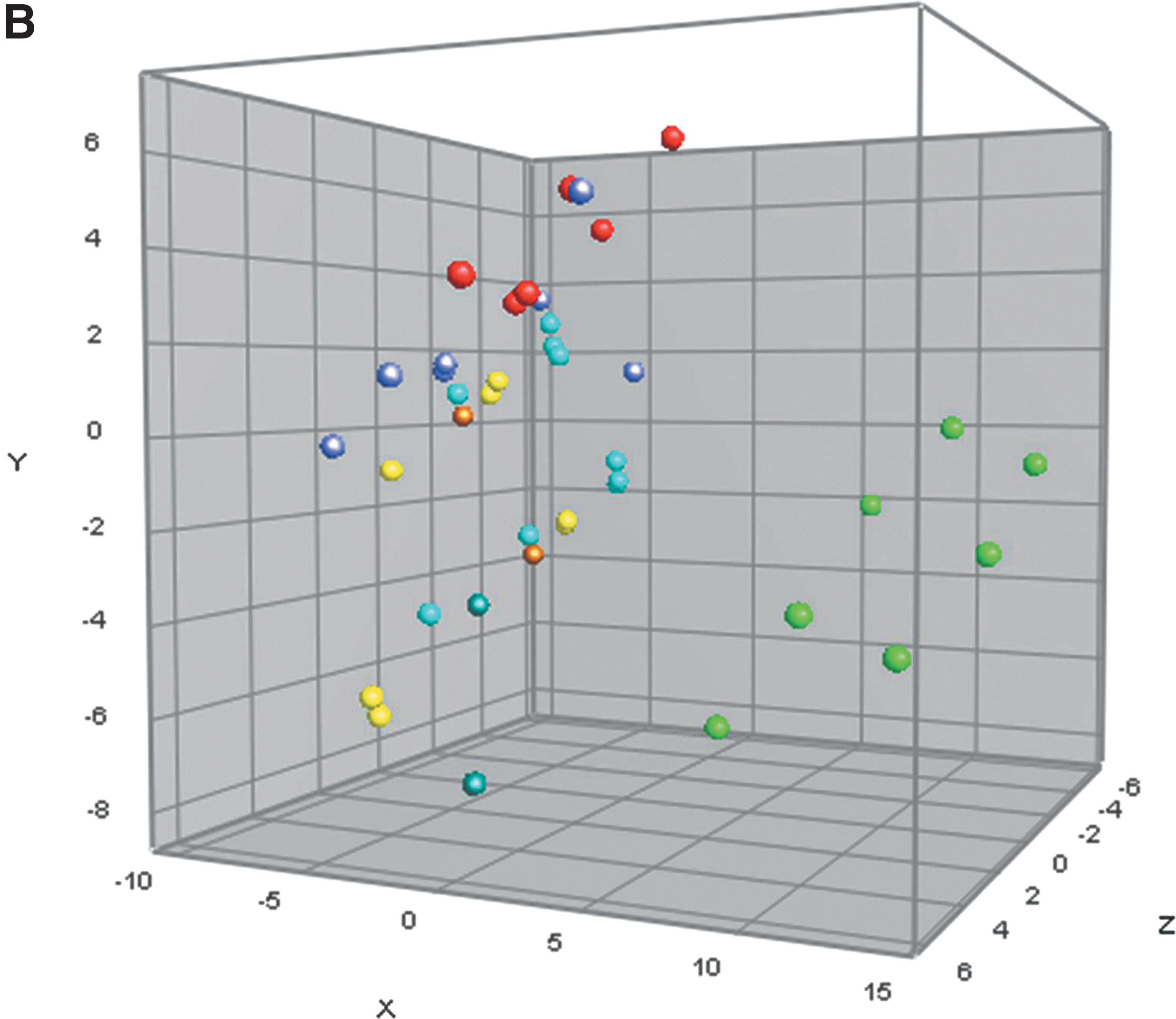
All isolates tested PCR positive for spaN, sipB, pagC, spiA, msgA, ttrB, spi4H, purR, iroN, fimH, rmbA, sopE, aceK, iacP, tolC, orgA, hilA, and invA, and negative for virD4 and virB4. The prevalence of other genes varied among the isolates; 77/95 (81%) were positive for avrA and 10, 6, 3, and 3 isolates were positive for intA, spvB, rhuM, and pefB, respectively. The PCA revealed an association between certain virulence genes and Salmonella serovars (Fig. 2A). Furthermore, PCA results indicated that different serotype clusters were associated with different virulence factor coordinates (Fig. 2B).
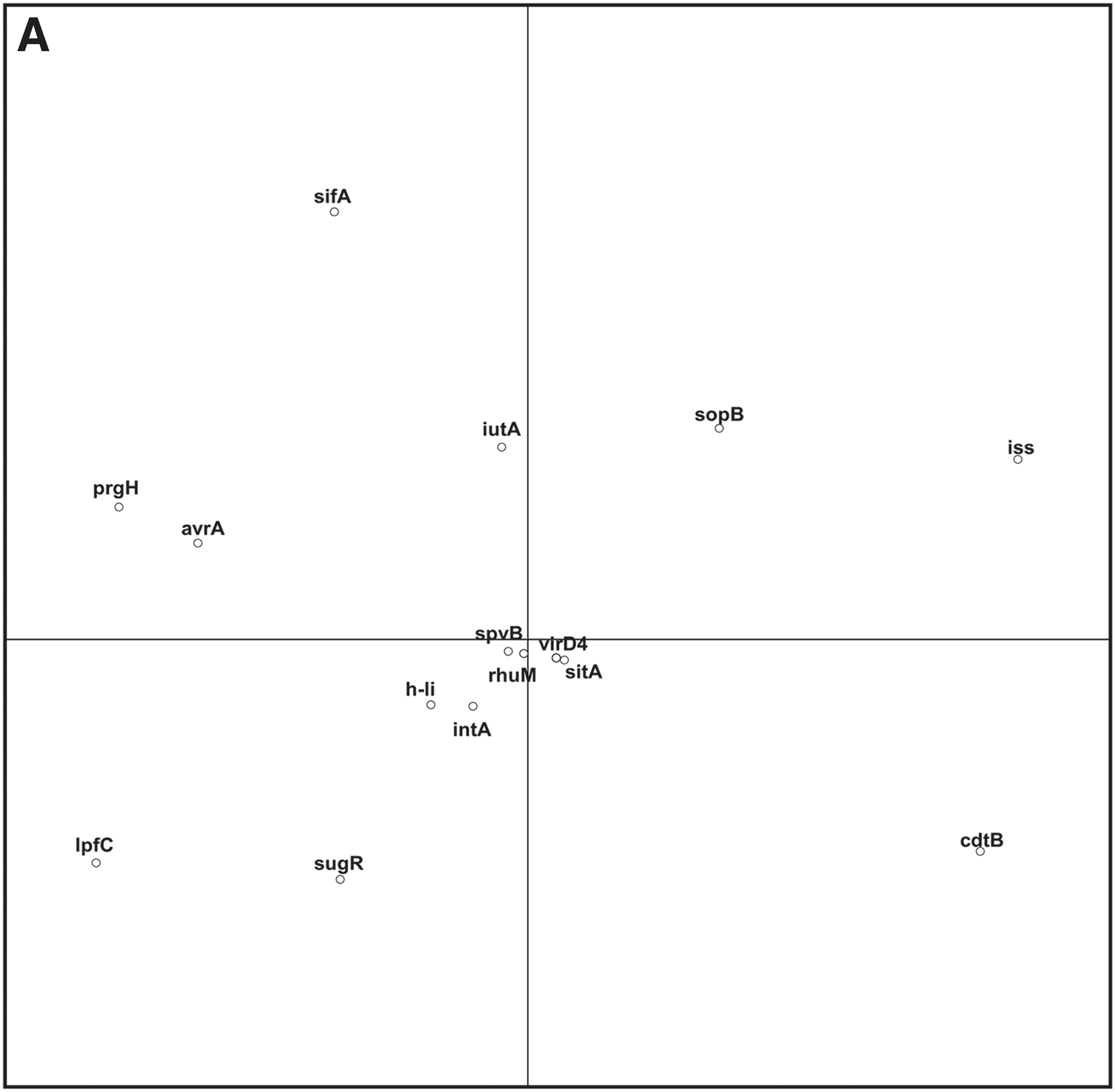
Results of the Principal Component Analysis of virulence factors versus serotypes. (
Discussion
Salmonella is commonly associated with contaminating raw and processed poultry in the United States (Nayak and Stewart-King, 2008; Jackson et al., 2013). The focus of this study was to characterize Salmonella strains that were isolated from a turkey flock in which the birds were raised in the absence of antimicrobial pressure. This approach included analysis of genotypic diversity, virulence factors, antimicrobial resistance, and plasmid characteristics of these Salmonella isolates. A potential limitation of the study was the inability to ascertain in detail the prevalence or frequency for Salmonella colonization in this flock due to limitations in the sample collection data available to the investigators.
In this study, Salmonella Montevideo was the most common serotype (24%) identified. In 2012, a multistate outbreak of Salmonella Montevideo infections in humans was linked to chicks, ducklings, and other backyard flocks (CDC, 2012). Salmonella Anatum was the second most identified serotype 21/95 (22%), which is a lower proportion than a study conducted by Rodriguez et al. (2006), who reported that Salmonella Anatum was the most common serovar isolated (48%) from different food animal farms, including chicken and turkey farms. Conversely, Salmonella Kentucky was the most prevalent serotype in other studies examining conventional and nonconventional (free-range, pasture, and organic) poultry production environments (Parveen et al., 2007; Melendez et al., 2010) and in commercial flocks (Santos et al., 2007). Turkey-associated human salmonellosis was most commonly associated with serovars other than Salmonella Enteritidis and Salmonella Typhimurium (Hald et al., 2012). Turkeys have been identified as reservoirs for human infections by Salmonella Kentucky, Salmonella Agona, and Salmonella Senftenberg (Hald et al., 2012; Jackson et al., 2013).
Antimicrobial resistance has been linked to the use of antimicrobials in production systems (Jacob et al., 2008). Multidrug-resistant pathogens can create a major problem for treatment of human and animal illnesses (Zhao et al., 2006; Maripandi, 2007; Miranda et al., 2008). In our study, the majority of the isolates (64%) were susceptible to all tested antimicrobials, which may be associated with the nonuse of therapeutic or subtherapeutic antimicrobials on the farm. Thirty-four of 95 (36%) isolates were resistant to at least 2 antimicrobials, which is lower than rates typically reported for isolates from traditional systems of raising turkeys. Nayak and Stewart-King (2008) reported that 77–100% of Salmonella strains from conventional turkey flocks were resistant to one or more antimicrobials. In another study, 86% of Salmonella isolates collected from commercial turkey farms were resistant to at least one antimicrobial (Santos et al., 2007). Class 1 integrons play a major role in antimicrobial resistance development in Salmonella; they typically encode sulfonamide resistance in their more conserved regions and can accumulate cassette-borne resistance genes in their variable region (Michael et al., 2006). Integrons located on plasmids can contribute to the dissemination of antimicrobial resistance among Salmonella (Cambray et al., 2010; Lai et al., 2013). In our study, class 1 integrons were detected in a relatively low proportion of isolates (8/95), compared to 68% in Salmonella from pasture-reared poultry (Melendez et al., 2010). This finding may help explain the overall low rate of antimicrobial resistance in the study population. A relatively low percentage of isolates also harbor larger plasmids that often carry antimicrobial resistance genes (Rodrigue et al., 1992; Melendez et al., 2010). The lack of antimicrobial pressure could select for isolates without integrons and plasmids to predominate or led to the loss of unneeded resistance plasmids by strains.
With this study, it was important to investigate the genetic diversity of Salmonella from turkeys in order to assess potential routes of transfer in the turkey production facility. Our PFGE data revealed that all isolates were clustered mostly based on their serotype, rather than source (Fig. 1). In some instances, Salmonella isolated from different sources in the facility possessed genotypes that were 100% similar. For example, in clusters III, IV, V, and VI, isolates from turkey ceca, feed, drinkers, and leftover feed were identical genotypically and phenotypically, which suggests possible cross-colonization. Conversely, 11 Salmonella Kentucky strains (cluster I) were isolated from feed and the environment but not from the turkey ceca, while 12 Salmonella Agona strains (cluster II) were identified in turkey ceca but not from feed, suggesting that colonization potential of either those serotypes may be source specific. Thus, the preharvest environment, including feed, is likely an important source of Salmonella contamination (Meerburg et al., 2006; Rodriguez et al., 2006; CDC, 2013; Ricke et al., 2013).
The link between genotyping, replicon typing, plasmid analysis, source, and the antimicrobial resistance profiles of Salmonella serovars was also evaluated. In cluster I, 10 of 13 isolates (8 Salmonella Kentucky isolates and 2 rough phenotypes) were mainly from feed and possessed the HI2 replicon (Fig. 1). They were resistant to at least three different classes of antimicrobials, including TET, SUL, and SMX. In previous studies, Salmonella isolates carrying IncHI2 plasmids were often resistant to multiple antimicrobials, including TET (Lai et al., 2013). In cluster IV, 4 of 11 isolates were resistant to at least 3 antimicrobials including SUL, STR, and GEN (Fig. 1). These four isolates were 100% identical in their PFGE profiles, had variable plasmids profiles, and all contained the IncI1 replicon. Conversely, in clusters II, III, and V that included Salmonella Agona, Montevideo, and Worthington isolates, respectively, no Inc types were detected in any of the isolates, whether or not plasmids were identified. Salmonella Worthington isolates 1149 and 1165 were positive for IncB/O and IncFIC replicons; however, they were susceptible to all the antimicrobials. All of the Salmonella Anatum (n = 20) and Salmonella Worthington (n = 4) isolates that possessed the IncI1 replicon were resistant to SUL and GEN, whereas 15 of the isolates were resistant to STR. All isolates but one possessed a 115-kb plasmid (Fig. 1). The IncI1-resistance plasmid encodes antimicrobial resistance to GEN, STR, and SUL (Johnson et al., 2011; Han et al., 2012). Our data indicated a strong association between possession of IncI1 replicon and resistance to SUL and GEN.
The PCA revealed an association between certain virulence genes and Salmonella serovars (Fig. 2A). All isolates tested PCR positive for 54% (19/35) of virulence genes tested, while presence of the remaining genes was variable among isolates. This was comparable to another study from our lab in which Salmonella isolates were positive for 58% of tested virulence genes (Zou et al., 2011). Our findings showed a distinct association between the presence of iss and sifA, as well as the absence of sugR and lptC in Salmonella Anatum (red dots in Fig. 2B). Conversely, the absence of iss and presence of sugR and lptC was associated with serotypes Worthington and Senftenberg (yellow and turquoise dots in Fig. 2B). Additionally, the association between the detection of iss and cdtB along with the absence of prgH and avrA was observed for Salmonella Montevideo (green dots in Fig. 2B), while the absence of iss and cdtB in conjunction with the presence of prgH, avrA, and lptC was common in serotypes Kentucky and Agona (blue and purple dots in Fig. 2B). Overall, the virulence gene profiles were similar within the isolates from the same serotype. In contrast, Zou et al. (2011) reported that the gene variability was independent of Salmonella serotype.
Conclusions
This study investigated the characteristics of Salmonella isolated from a turkey flock in which the birds were raised in the absence of selective antimicrobial pressure. Our findings highlight the importance of managing preharvest sources for control of Salmonella colonization in turkey flocks. The applied methods in this study provide a valuable tool to potentially trace the source and spread of Salmonella during turkey production cycles. Salmonella isolated in our turkey flock, while at lower rates than traditional turkey flocks, can still carry antimicrobial and virulence genes and harbor plasmids that may contribute to their pathogenicity and potential dissemination in the production environment. Nonetheless, farm biosecurity measures, such as all-in all-out production, cleaning and disinfection of the farm before, during, and between flock placements could reduce Salmonella that may persist on poultry farms. Poultry breeding programs that can render Salmonella-free poults to the production facility can also play a vital role in overall Salmonella colonization dynamics. Hence, it is critical to assess and identify preharvest sources (incoming birds, feed, water, etc.) and vectors (insects, fomites, etc.) of Salmonella contamination in a turkey production facility so that intervention strategies can be implemented to reduce or eliminate these bacteria before the birds are shipped to the processing units.
Footnotes
Acknowledgments
We thank Dr. Alessandra Carattoli for providing the positive controls for Incompatibility replicon typing and Dr. Bashar Shaheen for his technical expertise. We also thank Drs. Carl Cerniglia, Sangeeta Khare, and John Sutherland for reviewing the manuscript. Authors Yasser M. Sanad, Kelly Johnson, and Jing Han were supported through the Oak Ridge Institute for Science and Education. The views presented in this article do not necessarily reflect those of the U.S. Food and Drug Administration.
Disclosures
No competing financial interests exist.