
Abstract
Kudoa septempunctata, a myxosporean parasite infecting the trunk muscles of olive flounder (Paralichthys olivaceus), has been recently reported to be the causative agent of a type of food poisoning in humans. Patients exhibited acute diarrhea and vomiting after ingestion of the raw flesh of infected flounder. A recent increase in the number of food-poisoning cases has prompted us to develop a control strategy of this parasite. In this study, we evaluated the efficacy of several temperature and chemical treatments for inactivating K. septempunctata spores in vitro using the vital staining assay with the fluorescent dyes Hoechst 33342 and propidium iodide (PI). Screening tests of treatment methods against K. septempunctata suggested that 25% ethanol for 5 min, 80°C for 10 s, limonene at 10 μL/mL for 5 min, and salinities at 0‰ and 160‰ for 5 min were effective for killing spores. To verify toxicity loss in K. septempunctata spores after the treatments, tight junction barrier integrity assays with Caco-2 cells were conducted. The results of the Caco-2 assays corresponded well with those of the Hoechst 33342-PI staining assay. Further studies are required to determine a practical treatment procedure for inactivating spores considering the treatment application in the production process of cultured olive flounder.
Introduction
K
To date, several methods for determining the viability of myxosporeans have been developed, such as methylene-blue staining (Hoffman and Markiw, 1977), a double staining with fluorescein diacetate (FDA) and propidium iodide (PI) (Markiw, 1992; Yokoyama et al., 1997), and a fluorescent staining with Hoechst 33342 and PI (Yokoyama et al., 2009). Preliminary tests using K. septempunctata spores showed that methylene-blue and FDA-PI staining methods often yielded ambiguous results. Low stainability of K. septempunctata spores with FDA was observed as previously reported in Myxobolus artus (Yokoyama et al., 1997). This might be explained by the low permeability of the dye through the rigid spore valves of myxosporean spores. Alternatively, it might be caused by fading of the fluorescent dye. In any case, judgment of spore viability based on the FDA-PI staining was difficult. In contrast, a dual staining with Hoechst 33342-PI allows clear discrimination of live and dead cells. Hoechst 33342 penetrates the membranes of both live and dead cells, specifically binding nucleic acids, and PI is permeable only through the membrane of dead cells, rendering the nuclei of nonviable cells red and the nuclei of viable cells blue (Yokoyama et al., 2009).
The toxicity of Kudoa myxosporeans has been investigated using experimental animals, including house musk shrews and suckling mice (Kawai et al., 2012). House musk shrews vomited 20–30 min after feeding on flounder meat containing K. septempunctata spores, whereas suckling mice exhibited fluid accumulation in the gut within 1.5 h of oral administration of purified K. septempunctata spores. However, in vivo animal experiments are not suitable for screening of a wide range of treatment conditions inactivating spores. By contrast, an in vitro tight junction (TJ) barrier integrity assay with human intestinal epithelial cells (Caco-2) is useful for evaluating the toxicity of K. septempunctata (Ohnishi et al., 2013; Suzuki et al., 2015).
In the present study, we evaluated the efficacy of several temperature and chemical treatments for inactivating K. septempunctata spores in vitro using the vital staining assay with Hoechst 33342 and PI. Subsequently, toxicity loss of spores was verified by a measurement of TJ barrier integrity in intestinal Caco-2 cells.
Materials and Methods
Parasite samples
Juvenile olive flounder naturally infected with K. septempunctata were transported on ice to the Laboratory of Fish Diseases in the University of Tokyo from the National Research Institute of Aquaculture (Kamiura Branch, Oita Prefecture, Japan), where infected fish were stocked in a land-based tank. Somatic muscle was minced, passed through a 380-μm steel mesh (S-0770, Cell Dissociation Sieve Tissue Grinder Kit; Sigma, St. Louis, MO) and then through a 100-μm nylon mesh (Cell Strainer; BD Falcon, Corning, NY) with phosphate-buffered saline (PBS). The spore suspensions were centrifuged at 430 × g for 15 min, the supernatant was discarded, and the pellet was resuspended in 2.0 mL PBS. The spore suspension was placed into a 15%/30% Percoll gradient solution (GE Healthcare, Tokyo, Japan) in a 15-mL plastic tube. After centrifugation of the tube at 2330 × g for 30 min, the pellet was collected and resuspended in PBS. The purified spore suspension was again centrifuged at 430 × g for 15 min, and the pellet was collected and used as the parasite materials.
Viability assay by staining with Hoechst 33342-PI
A double staining with Hoechst 33342 and PI was performed according to Yokoyama et al. (2009), with slight modifications. A working solution of Hoechst 33342 (1 mM dissolved in PBS, bisbenzimide H33342 trihydrochloride, Sigma) and PI (0.02 mg/mL dissolved in PBS, Sigma) were prepared. Twenty-five microliters of spore suspension, 25 μL of PI solution, and 50 μL of Hoechst 33342 solution were combined in a 1.5-mL microtube and kept in the dark at 5°C for 60 min. After a coverslip was placed over a drop of 25 μL of the mixture on a glass slide, the spores were observed with a fluorescent microscope (BX60; Olympus, Tokyo, Japan). Spore viability was enumerated on the basis of 100 spores; spores with only nuclei showing blue fluorescence (Hoechst 33342 positive and PI negative) were considered viable (Fig. 1), whereas spores with even one nucleus showing red fluorescence (Hoechst 33342 positive and PI positive) were considered dead.
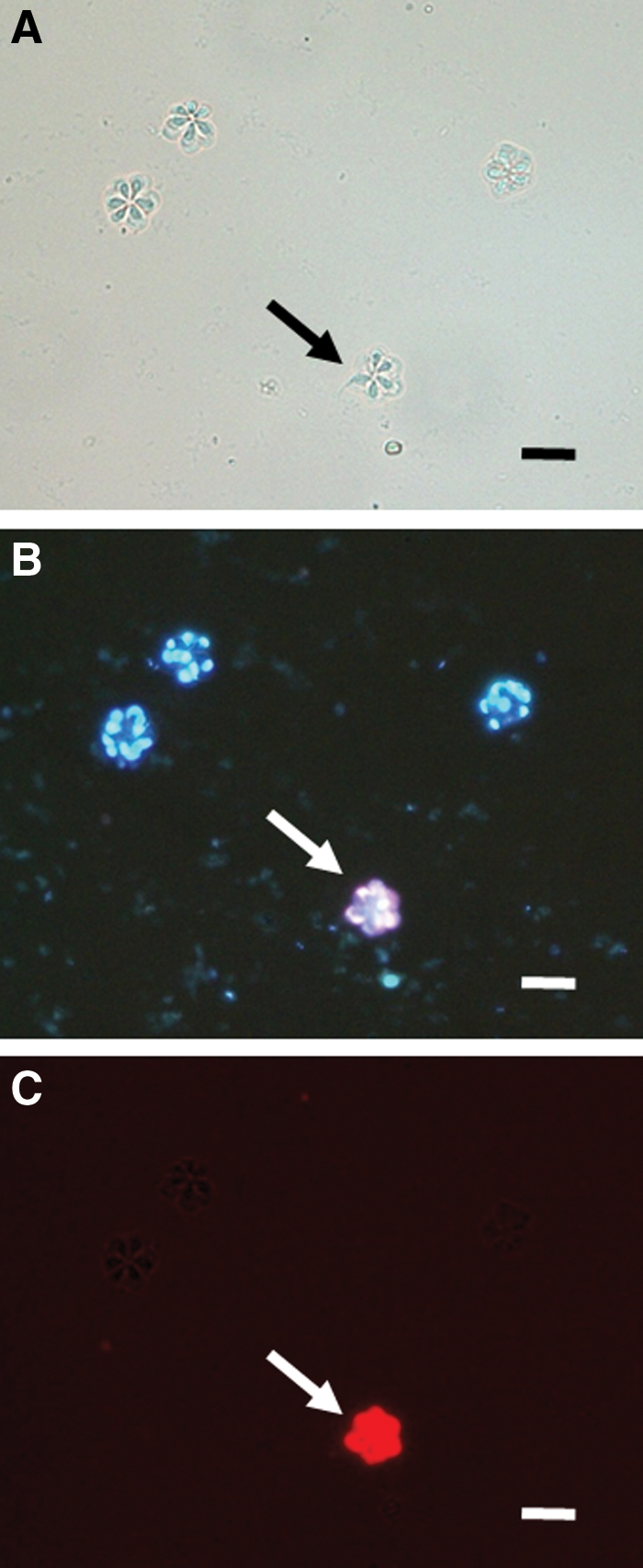
Fresh spores of Kudoa septempunctata stained with Hoechst 33342 and propidium iodide (PI) observed under normal light
Treatments inactivating K. septempunctata spores
For temperature, microwave, and chemical treatments, all experiments were conducted in triplicate, and the number of spores counted in each replicate was 100. Chi-square analysis (p < 0.01) was performed to estimate statistically significant differences between treated and untreated spores.
Temperature
Spores were stored in a deep-freezer (−80°C) for 16 h, a refrigerator (4°C) for 3 days, or a chilled incubator (0°C) for 3 days. Additionally, spores were put in a 0.2-mL tube and heated to 80°C for 10, 30, and 60 s with an iCycler (Bio-Rad, Hercules, CA). Untreated spores were used as the negative control. As positive controls, spores were incubated at 75°C for 5 min, and −20°C for 16 h, which is the inactivation treatment recommended by the Ministry of Health, Labor and Welfare, Japan. Additionally, to investigate in vitro long-term viability of spores at low temperatures, the spore suspensions supplemented with an antibiotic mixture (500 U/mL penicillin G and 500 μg/mL streptomycin) were held at 0°C and 4°C. Thereafter, spore viability was monitored over 2 months.
Microwave
Preliminary tests with noninfected olive flounder slices by visual observation showed that microwave conditions at 100 W and 170 W for 10 s, and 300 W for 5 s, preserved raw flounder slices without heat denaturation. Thus, spore suspensions in a 15-mL plastic tube were placed in a microwave oven (NE-EH211; Panasonic, Osaka, Japan), and treated at 100 W for 5–15 s, 170 W for 5–15 s, and 300 W for 5–10 s, followed by assessment of spore viability.
Chemicals
Besides sashimi, olive flounder is often used in dishes (zuke) soaked in liquor (Japanese sake) and soy sauce. Recently, kabosu hirame, flavored olive flounder fed with a type of citrus fruit (Citrus sphaerocarpa) containing abundant limonene, has been successfully commercialized. Therefore, ethanol and d-limonene were included in chemical treatment tests. Solutions of ethanol, d-limonene (Wako Pure Chemical Industries, Ltd., Osaka, Japan), and sodium chloride were added to spore suspensions of K. septempunctata and placed in the dark at 4°C. Concentrations and durations of ethanol were 5, 15, 25, 50, 70, and 100% for 1 h, and 25% for 5 min and 15 min. Concentrations and durations of d-limonene were 0.1, 1.0, and 10.0 μL/mL for 1 h, 1.0 and 10.0 μL/mL for 5, 15, and 60 min. Concentrations and durations of sodium chloride were 0, 3, 6, 9, 17.5, 35, 80, and 160‰ for 5 min. After centrifugation of the suspensions, the supernatant fluid was discarded, the pellet was resuspended in PBS, and then spore viability was determined as described above.
Measurement of TJ barrier integrity of Caco-2 cells
The human adenocarcinoma cell line Caco-2 (European Collection of Cell Cultures [ECACC] 86010202) was cultured in Eagle's minimal essential medium (MEM; Nissui Pharmaceutical, Tokyo, Japan) supplemented with 2 mM glutamine (MP Biomedical, Santa Ana, CA), 10% fetal bovine serum (Gibco/Life Technologies, Carlsbad, CA) and 1% nonessential amino acids (Gibco) at 37°C in 5% CO2. For differentiation of Caco-2 cells in vitro, the Biocoat HTS Caco-2 Assay System (BD Biosciences, Bedford, MA) was used. Caco-2 cells were suspended in MEM medium, seeded at 2 × 105 cells/well in the Biocoat cell culture insert, and then incubated at 37°C in 5% CO2. After 24 h, the culture medium was replaced with enterocyte differentiation medium with MITO and serum expander of the Caco-2 assay system and incubated for 24 h. After 24 h, the medium was again replaced with the same new medium and incubated for 24 h. The cell monolayer integrity was checked by measuring transepithelial electrical resistance (TER) with an epithelial voltohmmeter (World Precision Instruments, Sarasota, FL). Inserts showing a TER of more than 1000 Ωcm2 were used for the assay. Fresh spores of K. septempunctata were suspended in enterocyte differentiation medium with MITO and serum expander of the Caco-2 assay system. As an untreated control, fresh spore suspensions (1 × 106 spores/well) were inoculated into the cell culture inserts, and the TER was measured every hour.
To verify spore inactivation by several treatments, spores of K. septempunctata treated with 25% ethanol for 5 min, 80°C for 10 s, limonene at 10 μL/mL for 5 min, and salinities at 0, 35, and 160‰ for 5 min were subjected to the Caco-2 assay described above. Data of 25% ethanol, 80°C, and limonene treatments were obtained from three wells, whereas data of salinity treatments were from four wells containing an identical number of spores. Statistical analyses (p < 0.01, Student t-test) were based on comparison with the negative (medium) control.
Results
Effects of temperature on spore viability
The mean viability of untreated spores (the negative control) was 90%, whereas those of spores incubated at 75°C and −20°C (the positive controls) were 0% (Fig. 2). Spores stored at −80°C for 16 h were killed, but those at 0°C and 4°C for 3 days were still viable (90.7% and 90.4%, respectively). Heat treatments (80°C) for 10−60 s were found to eradicate the spores. The long-term viability tests of spores kept at 0°C and 4°C revealed that approximately 80% of spores remained viable for 2 months in the low temperatures, although the viability decreased 57% at Day 74 (Fig. 3).
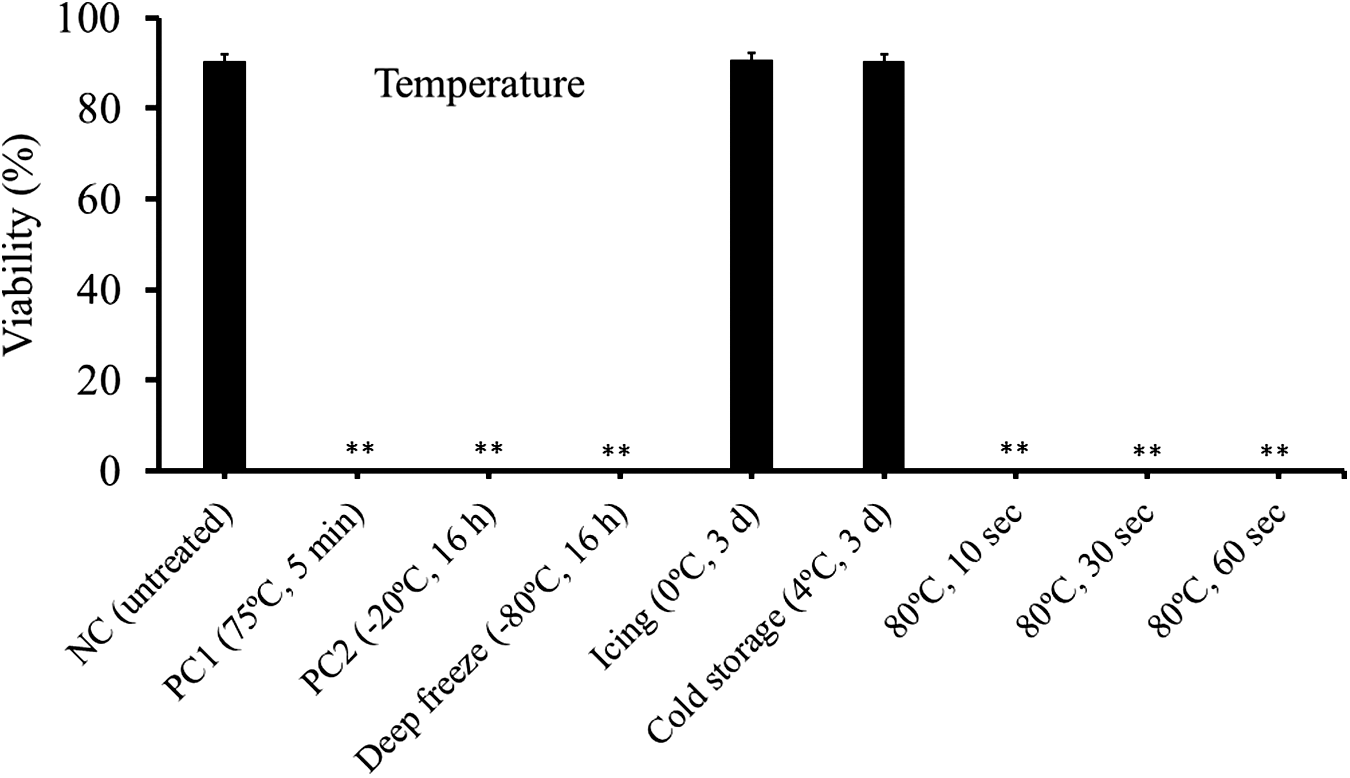
Viability of Kudoa septempunctata spores incubated for different lengths of time at several temperatures as determined by Hoechst 33342 and propidium iodide stain. NC shows the negative control (no incubation). PC1 and PC2 indicate the positive controls (heating at 75°C for 5 min and freezing at −20°C for 16 h), respectively. **Significantly different from the negative control (p < 0.01). Vertical bars represent standard error.
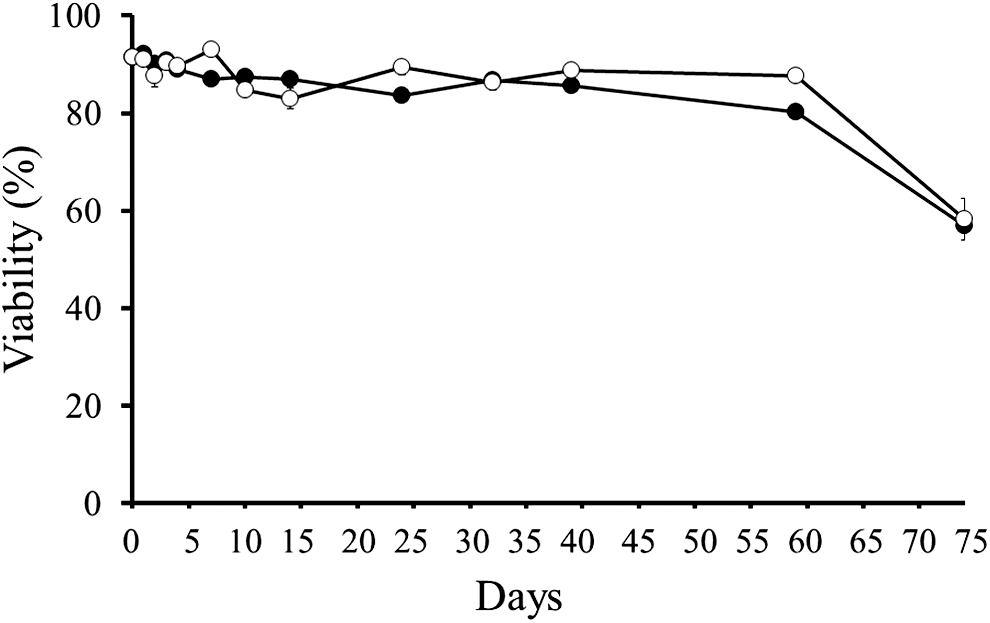
Long-term viability of Kudoa septempunctata spores kept at 0°C (black circles) and 4°C (white circles) over time as determined by Hoechst 33342- propidium iodide stain. Vertical bars represent standard error.
Microwave
Spores were not fully inactivated by all microwave conditions tested in the present study (Fig. 4). Moderate sporicidal effects were observed in the microwave condition at 300 W for 10 s, but this condition denatured the flounder meat as observed by visual observation.
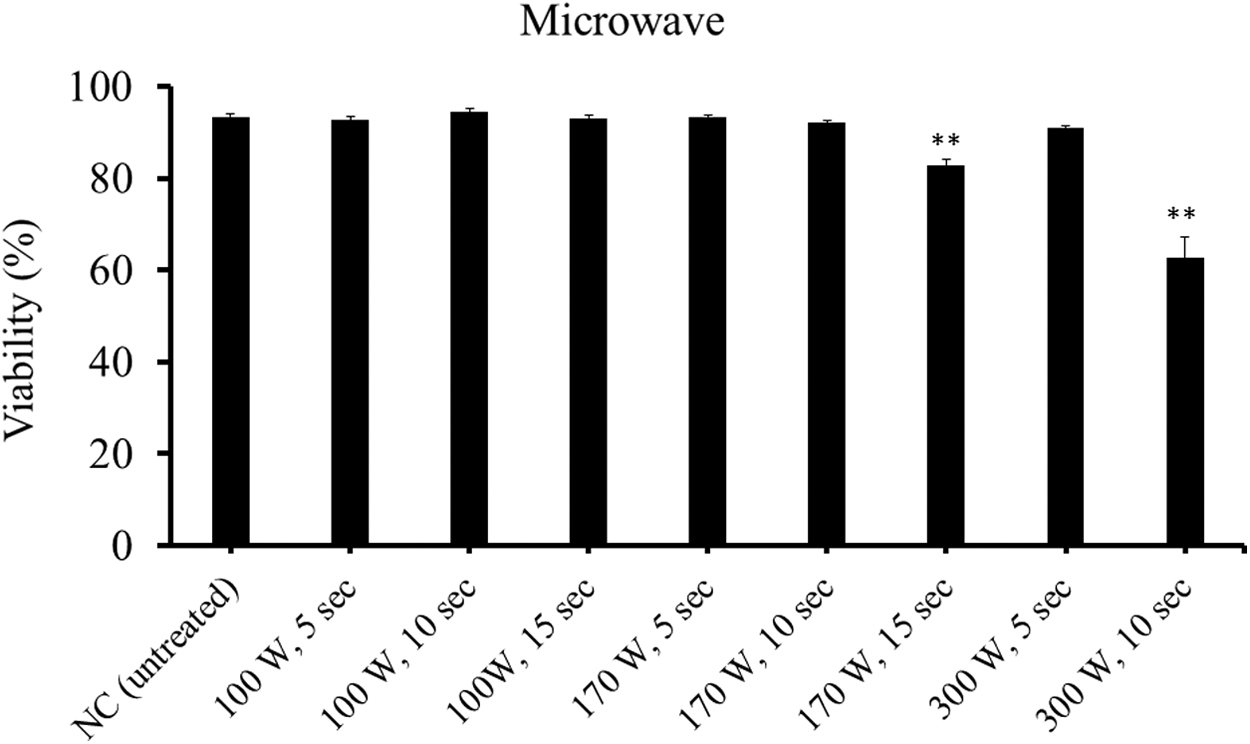
Viability of Kudoa septempunctata spores treated with microwave radiation at various outputs for different lengths of time as determined by Hoechst 33342-propidium iodide stain. NC shows the untreated negative control. **Significantly different from the negative control (p <0.01). Vertical bars represent standard error.
Chemicals
Treatments with more than 25% ethanol for 1 h completely inactivated spores (Fig. 5). In treatments with 25% ethanol, even 5 min was enough to kill the spores. Treatment with limonene at 0.1 μL/mL for 1 h was not effective, but those at 1 and 10 μL/mL for 1 h was effective (Fig. 6). Those at 1 μL/mL for 5 min and 15 min were only moderately (approximately 50% viability) effective (Fig. 6). By contrast, 10 μL/mL-limonene inactivated spores consistently between 5 min and 60 min. In a salinity equivalent to seawater (35‰) and one-half seawater (17.5‰), more than 70% of spores were viable, whereas freshwater (0‰) and 160‰ sodium chloride solution killed spores (Fig. 7).
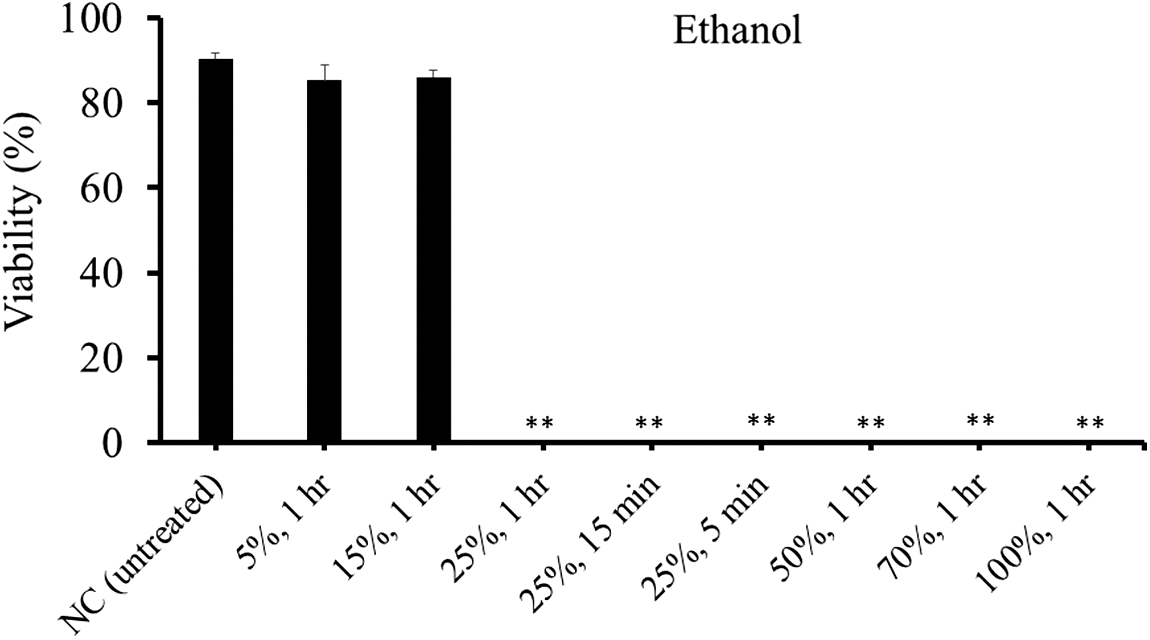
Viability of Kudoa septempunctata spores treated with ethanol at various concentrations for different lengths of time as determined by Hoechst 33342-propidium iodide stain. NC shows the untreated negative control. **Significantly different from the negative control (p < 0.01). Vertical bars represent standard error.
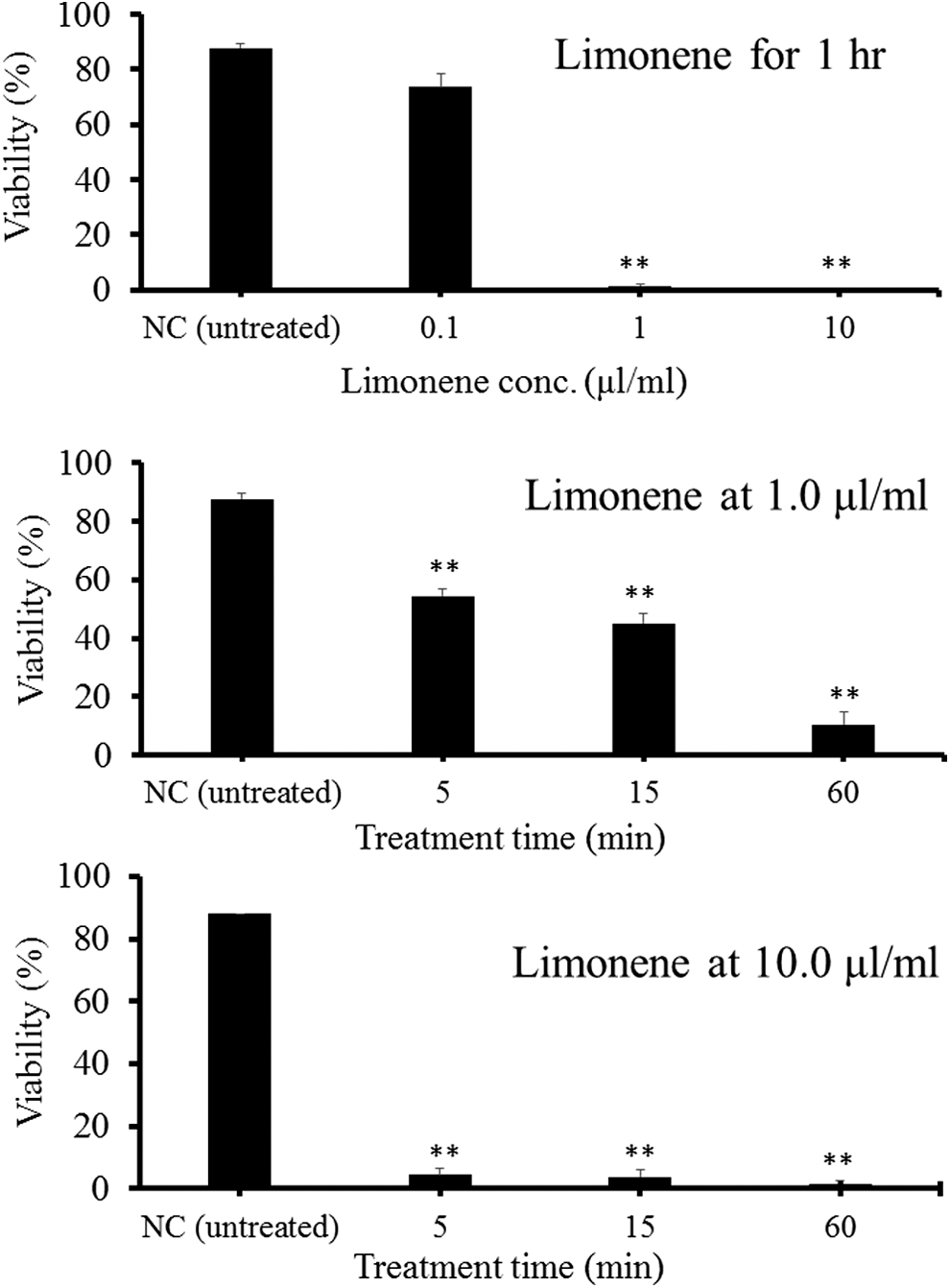
Viability of Kudoa septempunctata spores treated with limonene solution at various concentrations for different lengths of time as determined by Hoechst 33342-propidium iodide stain. NC shows the untreated negative control. **Significantly different from the negative control (p < 0.01). Vertical bars represent standard error.
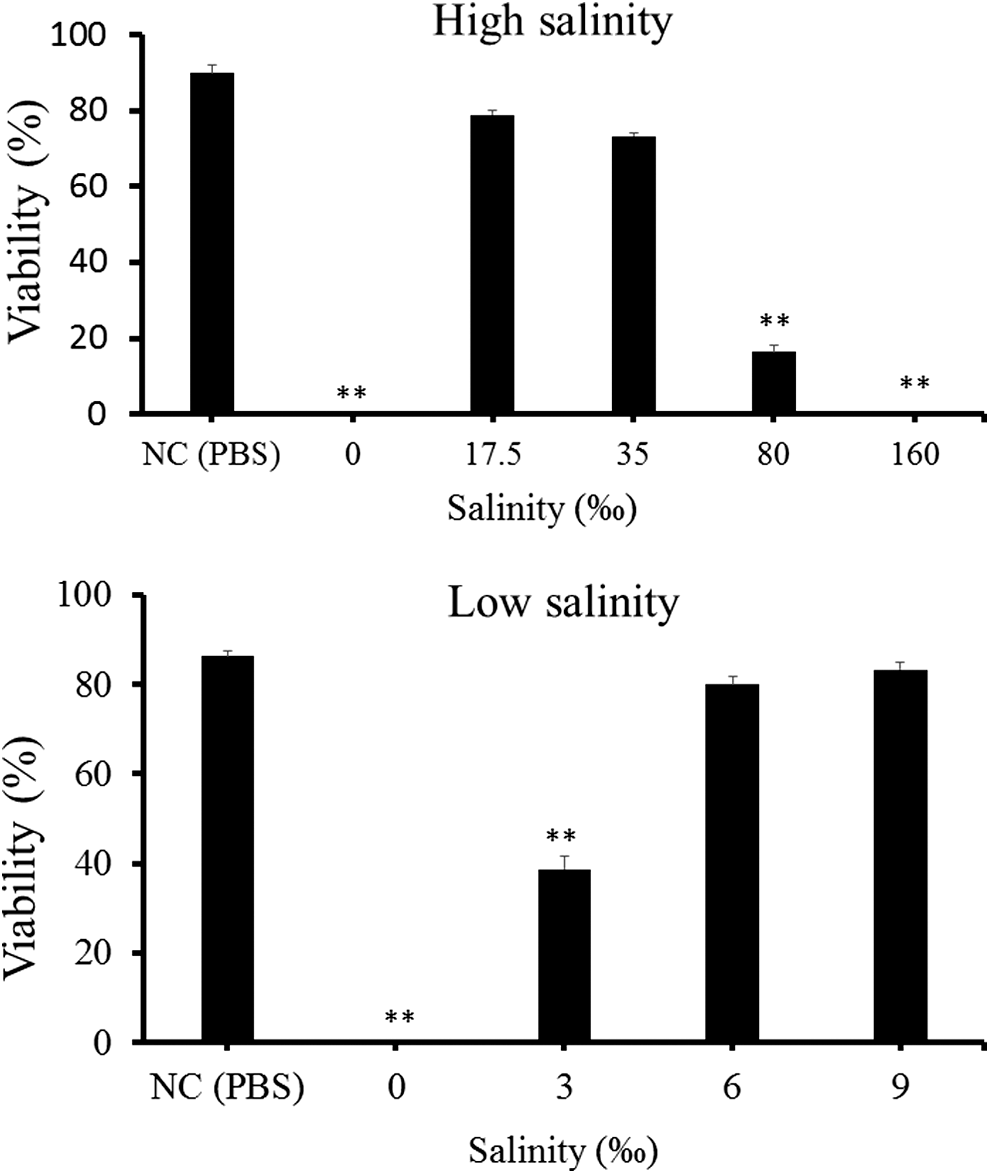
Viability of Kudoa septempunctata spores treated with sodium chloride solutions at high (top) and low (bottom) salinities for 5 min as determined by Hoechst 33342-propidium iodide stain. NC shows the negative control treated with phosphate-buffered saline (PBS). Asterisks indicate results that were significantly different from the negative control (*p < 0.05, **p < 0.01). Vertical bars represent standard error.
Measurement of TJ barrier integrity of Caco-2 cells
The TER in the wells inoculated with fresh spores of K. septempunctata sharply decreased by 80% after 2 h, but the TER was not significantly changed in wells with spores treated by 25% ethanol, 80°C for 10 s and 10-μL/mL limonene for 5 min (Fig. 8). In another experiment to evaluate the effect of sodium chloride, spores treated with 0‰ and 160‰ salinity did not affect the TER (Fig. 8). Spores treated with 35‰ salinity reduced the TER (by 64%) but not as much as the untreated fresh spores (by 80%).
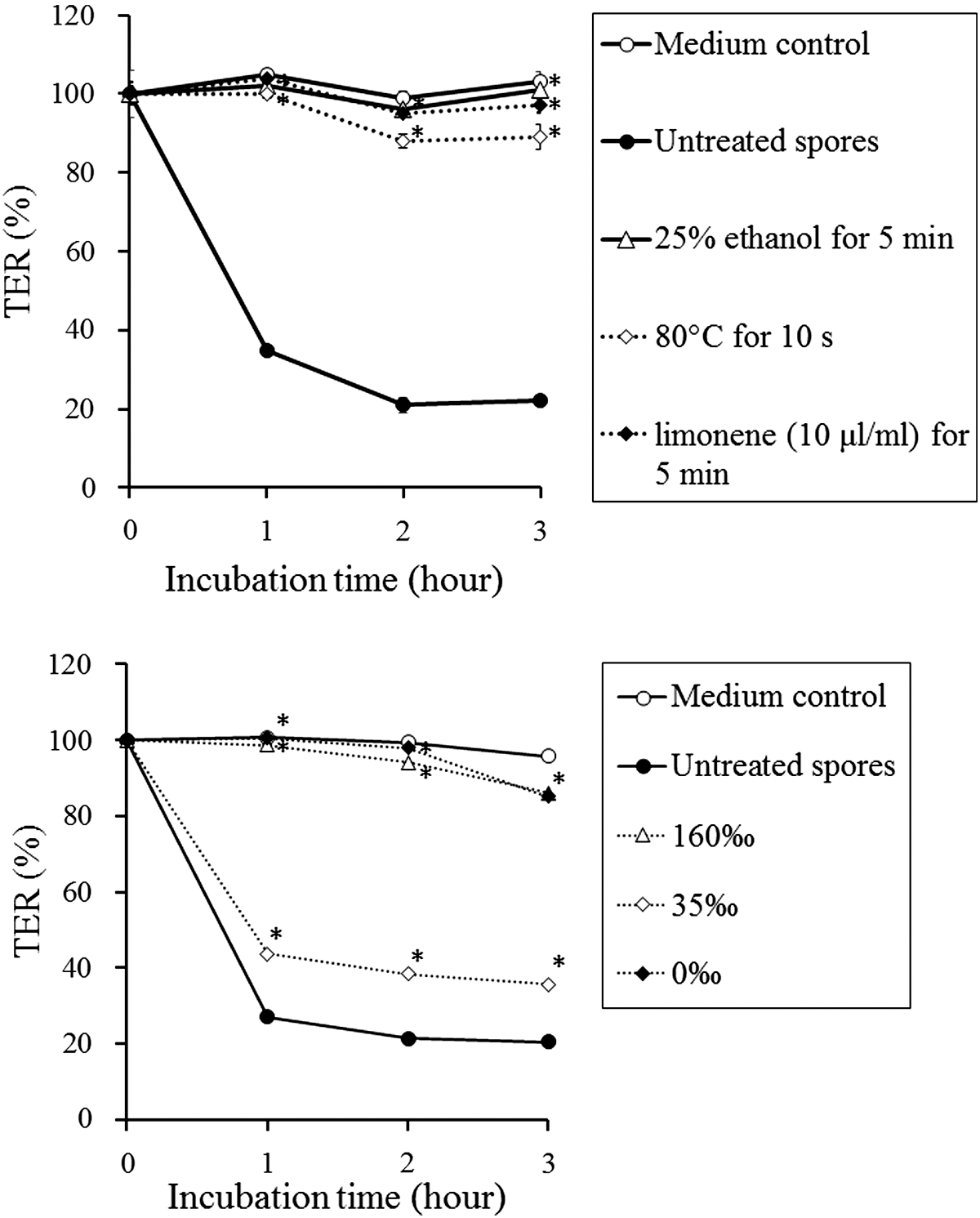
Transepithelial electrical resistance (TER) in the Caco-2 cell monolayer assay was measured for 3 h, with TER at time 0 represented as 100%. Each value is the mean ± standard deviation of three (top graph) or four (bottom graph) wells containing identical numbers of spores. Asterisks show the significant difference from the negative (medium) control (p < 0.01, Student t-test).
Discussion
The present study demonstrates that double staining with Hoechst 33342 and PI is a useful tool to evaluate the viability of K. septempunctata spores. Multicellularity of myxosporeans often caused mixed staining patterns of nuclei, showing blue and red fluorescence, resulting in confusion of judgment on spore viability (Yokoyama et al., 2009). However, the results of the Hoechst 33342-PI staining assay defined in the present study corresponded well to those of the toxicity evaluated by the Caco-2 cell assay, suggesting that the vital staining assay could be a potential indicator of spore toxicity.
Several temperature treatments were found to inactivate K. septempunctata spores. Deep-freezing of spores at −80°C for 16 h was effective, but it is likely that the texture of the treated meat is not suitable for eating as sashimi. Ohnishi et al. (2014) indicated that K. septempunctata spores were inactivated by air blast freezing at −80°C for 1 h, but that the meat quality deteriorated after freezing. Short-term heating at 80°C for several seconds is how the Japanese dish shabu-shabu, a hot-pot dish of thinly sliced meat dipped in boiling broth, is cooked. Although shabu-shabu with yellowtail (Seriola quinqueradiata) slices has become popular in Japan (referred to as Buri-shabu), eating olive flounder as shabu-shabu has not generally been accepted. The long-term (>2 months) survival of spores kept at 0°C and 4°C indicates that cold storage is impractical for detoxification of infected flounder. However, for research purposes, it suggests that spores can be preserved in a refrigerator for a long time. Spores kept in the refrigerator for up to a month may be used for toxicity tests. Yokoyama et al. (1997) reported that spores of M. artus from the trunk muscle of the common carp Cyprinus carpio remained viable for 15 mo at 5°C.
Treatment with microwaves and ethanol did not yield any practical condition for inactivating K. septempunctata spores in olive flounder. Microwave-treated meat is highly susceptible to heat denaturation even under moderate power output (300 W) for 10 s, in which only approximately 30% of spores were killed (Fig. 4). Greene and Babbitt (1990) applied microwave cooking to prevent arrowtooth flounder (Atheresthes stomias) infected with K. thyrsites from postmortem myoliquefaction. However, it will be hard to practice as long as olive flounder is consumed raw. Treatment with 25% ethanol for 5 min killed spores, but it also appears impractical. Even if infected olive flounder were immersed in 25% ethanol for 5 min, the ethanol will not penetrate deep into the musculature. Intravenous injection of 25% ethanol is likely impossible without killing the olive flounder.
By contrast, in-feed treatment with limonene may be feasible for inactivating K. septempunctata infecting olive flounder in vivo. d-Limonene is one of the most common terpenes with a lemonlike odor and is widely used as a flavoring agent in many food items and beverages (Sun, 2007). d-Limonene is generally considered to have a low toxicity, but the toxic effects were also reported at 5.6–6.6 g/kg in mice (Sun, 2007). Because there is no information on the limonene concentration in the meat of kabosu hirame (flavored flounder) on the market, toxicity for humans can hardly be estimated. It is necessary to evaluate a total amount of limonene ingested by humans who eat olive flounder with the limonene applied at 10 μL/mL. Nevertheless, it is worth considering whether oral administration of limonene is effective to treat and prevent olive flounder infection with K. septempunctata.
Results of salinity tests provide us with some suggestions for potential control methods. In a land-based culture facility, cleaning the tank using freshwater is recommended as is infection with Enteromyxum leei (Yokoyama and Shirakashi, 2007). It may prevent transmission of K. septempunctata spores to the alternate invertebrate host (and likely vice versa), hence impeding completion of the parasite life cycle. A high salinity treatment may be applied to the cooking process because 160‰ salinity is approximately equivalent to the salinity of soy sauce. However, only dipping the flounder slice in soy sauce will not be effective because the high salinity of soy sauce works only on the surface of the sashimi.
Conclusions
In vitro inactivation of K. septempunctata spores by several temperature and chemical treatments was demonstrated, using the viability assay with Hoechst 33342-PI and the toxicity assay with Caco-2 cells. These treatments may be useful for preventing parasite contamination in equipment and materials for an experiment in a laboratory. However, application of these treatments to infected flounder in fish farms or markets needs further studies.
Footnotes
Acknowledgments
We are grateful to Drs. Koh-ichiro Mori, Jun Satoh, Tohru Mekata, and other staff members of the National Institute of Aquaculture for providing the fish samples. This research was partly funded by the Ministry of Agriculture, Forestry and Fisheries of Japan (No. 2403).
Disclosure Statement
No competing financial interests exist.