
Abstract
Salmonella enterica (nontyphoidal) is one of the major causes of foodborne diseases in the United States and worldwide. Molecular typing methods are significant tools used to better understand the transmission and ecology of Salmonella in order to implement pre-harvest control measures. The objectives of this study were to describe the Salmonella genotypes, the distribution of isolate subtypes from different ecological niches (i.e., barn environment, nursery, and individual pigs) and their evolution over time in a longitudinal study conducted in three finishing sites (housing pigs from 10 weeks of age until slaughter at 24–26 weeks of age). Among the 107 Salmonella isolates submitted for pulsed-field gel electrophoresis (PFGE) analysis, there were 25 distinct subtypes. PFGE genotyping results were consistent with the serotype findings. A large number of distinguishable PFGE patterns (i.e., within the same serovar) were observed and different combinations of subtypes were identified within and across sites and cohorts. New subtypes may result of the introduction of new strains, genetic changes, or ongoing transmission of evolved strains within the production system. The same subtypes were detected intermittently during the study period, which suggests the persistence of indistinguishable subtypes in this production system. In addition, this study suggests persistence of the same subtype over several cohorts of pigs and potential residual contamination from the barn. Factors affecting adaptation and transmission of Salmonella within and among ecological systems (e.g., finishing pigs, nursery, and environment) should be further investigated. Understanding genotypic diversity of Salmonella in different ecological niches during pre-harvest may contribute to the development of more targeted and cost effective control programs during nursery and finishing phases.
Introduction
S
The majority of control measures have been focused on decreasing the Salmonella prevalence (fecal shedding) in swine during pre-harvest (Lo Fo Wong et al., 2002; Fosse et al., 2009; Soumpasis et al., 2012), which is expected to decrease carcass cross-contamination in the slaughterhouse. Despite the fact that certain serovars pose more of a threat to human health than others (Pires and Hald, 2010; CDC, 2013a, 2013b), the control measures at the farm level have been focused on prevalence reduction (Dahl et al., 1997; Funk and Gebreyes, 2004; Fosse et al., 2009; Alban et al., 2010; Soumpasis et al., 2012). Molecular typing methods can be used to better understand the transmission and ecology of Salmonella in the farm environment (Vigo et al., 2009; Molla et al., 2010; Kich et al., 2011) and pork contamination (Wonderling et al., 2003; Piras et al., 2011; Hernandez et al., 2013). Pulsed-field gel electrophoresis (PFGE) with XbaI restriction enzyme has been used to determine the genetic relatedness of strains from different sources (i.e., swine environment, trucks, individual pigs, and carcasses), mainly in cross-sectional studies (Wonderling et al., 2003; Gebreyes et al., 2004b; Gebreyes et al., 2006; Kich et al., 2011; Denis et al., 2013; Hernandez et al., 2013). These methods are also used routinely to investigate human outbreaks (Ribot et al., 2006). There are fewer reports of using these methods to evaluate changes in strain relatedness over time and within farms (Vigo et al., 2009; Molla et al., 2010). Such knowledge might be used to develop and implement more targeted control measures and surveillance programs based on specific serovars and genotype distribution.
This study was conducted to gain a better understanding of the transmission and ecology of Salmonella and demonstrate the relatedness of Salmonella isolates from different ecological niches (i.e., barn environment, nursery, and individual pigs) and evolution over time in three finishing sites, belonging to a unique production system in the midwestern United States.
Materials and Methods
Salmonella isolates and serotyping
All of the Salmonella isolates in this study originated from another study that is described in detail elsewhere (Pires et al., 2013a; Pires et al., 2013b). The samples were collected during a longitudinal study on a multisite, farrow-to-finish production system located in the Midwestern United States. Briefly, the production system had three-site management (i.e., breeding and farrowing, nursery and finishing housed at a separated site), with all-in/all-out management in nursery and finishing sites. This system consisted of 2 farrowing sites (F1 and F2), 2 nursery sites (N1 and N2), and 12 finishing sites and transitioned from 2 farrowing sites to one during the study period. One barn (housing 1000 pigs) in three separated finishing sites (sites A, B, and C) was selected, and six consecutive cohorts of pigs were followed (18 cohorts total). Sites A and B were finishing sites (age 10–26 weeks). Site C transitioned to a weaning-to-finishing site after the second cohort of pigs. For the weaning-to-finishing cohorts at site C, piglets were placed in the barn at age 3 weeks and remained until marketing. Finishing site A received pigs from nursery N1 in all cohorts; site B received pigs from nursery N2 in four cohorts and from N1 and N2 in the last two cohorts of pigs. The first cohort for site C was supplied from nursery N2. For all other cohorts, piglets were placed directly from the farrowing sites due to the transition to a weaning-to-finishing site. Fifty individual pig fecal samples per cohort were targeted for collection every 2 weeks for 16 weeks (18 cohorts ×50 pigs/cohort ×8 collections = 7200 fecal samples targeted; 6836 samples actually collected).
In order to evaluate the Salmonella status of the cohort of pigs prior to entering the finishing phase, 10 pooled fecal samples (5 g fresh fecal material collected from five different locations on the same pen floor, 25 g/pool) were collected from the nursery rooms approximately 1 week prior to their movement to the finishing barns. Contamination of the selected barns was assessed by culture of barn environmental samples (drag swabs and swabs from floors, walls, gates, and feeders/drinkers; total of 20 samples per cohort) after cleaning and disinfection and before placement of each cohort of pigs in the barn. Fecal, nursery, and environmental samples were cultured using standard methods described elsewhere (Pires et al., 2013a). Salmonella was cultured from 6.6% (453/6836) of the individual fecal samples, 36.5% (67/170) of the pooled nursery samples and 11.1% (40/360) of the environmental samples (Pires et al., 2013a; Pires et al., 2013b).
A total of 187 Salmonella-positive pigs (defined as a pig that had at least one Salmonella-positive fecal sample) were identified in this study. A total of 446 isolates (out of 453) from individual pig samples were serotyped and used as the sampling frame for the PFGE genotyping study. Identification of serotypes was conducted according to standard methods by MSU Diagnostic Center for Population and Animal Health and the National Veterinary Services Laboratory, Ames, IA (Pires et al., 2014).
PFGE genotyping
Salmonella isolates from individual pigs, pooled nursery and environment barn samples were genotyped using PFGE. The isolates were selected in order to investigate spatiotemporal variation between sites, within cohort and between cohort and their respective nursery and barn environment. The isolates were selected systematically based on the following criteria: (1) the top three serovars (by frequency of isolation): S. Derby, S. Agona and S. Johannesburg; (2) matched individual pig isolates of the same serovar were selected within site, within cohort, and between sites; (3) matched individual pig isolates of the same serovar present in the nursery or environment and respective cohort; and (4) matched isolates of the serovars Salmonella Meleagridis, Litchfield, and Infantis and correspondent nursery and environment samples. A systematic selection was conducted across and within sites, cohorts, and pigs.
The sample origins for the 107 isolates (1 isolate per positive sample) selected for inclusion in this study were as follows: 89 isolates from individual pig samples (of 446 total isolates), 14 isolates from pooled fecal samples from source nurseries (of 62 isolates), and 4 isolates from barn environment samples (of 40 isolates).
Briefly, PFGE was conducted according to a Centers for Disease Control and Prevention standardized protocol (Ribot et al., 2006). Overnight culture cells (200 μL) were lysed and digested with a single restriction enzyme (XbaI). The digested agarose-embedded DNA was then separated using a CHEF-DR III pulsed-field gel electrophoresis system (BioRad Laboratories, Hercules, CA). The gel was stained in ethidium bromide (1 μg/mL), and a Gel Doc 2000 (BioRad) was used to capture fingerprint images. Captured images were subsequently analyzed within GelCompar II (v. 6.5, Applied Maths, Austin, TX) by applying hierarchical agglomerative clustering techniques (unweighted pair group method with arithmetic means) to similarity matrices calculated using the dice coefficient of similarity. Salmonella enterica servoar Braenderup H9812 was used as a molecular reference marker. Band tolerance and optimization settings of 1.5% were used. PFGE patterns were considered distinct and given a unique pattern number based on at least one band difference in the pattern (Barrett et al., 2006); however, the relatedness and clustering was also considered in the interpretation of the results. Fragments that were smaller than 20 kb were not included in the cluster analysis. The proportion of samples positive for each subtype was described across sites, cohorts, and pigs.
Results
PFGE analysis identified 25 distinguishable subtypes among the107 isolates. Dendrograms were constructed to examine the similarity of Salmonella banding patterns from samples collected across and within sites, cohorts, and pigs. In general, PFGE subtypes clustered by Salmonella serovar (Supplementary Fig. S1, Table 1; Supplementary Data are available online at
Three sites (A,B, C).
Six cohorts in each site (site A, B, and C, cohorts C1 to C6).
Total Salmonella pig isolates per PFGE subtype.
Total of pigs per PFGE subtype.
Twelve cohorts had Salmonella isolates of the same serovar identified in both nursery and finishing pig samples. All 12 cohorts had the same PFGE subtypes identified in both the nursery and pig isolates. For example, the PFGE subtype 11 of Salmonella Derby was identified in the nurseries and respective pigs of cohort 5 of site A and cohort 4 of site B (Fig. 1A, Table 1). Similarly, the same PFGE subtype 4 of Salmonella Agona was identified in nursery samples and individual pigs in cohorts 1 and 2 of site A (Fig. 1B, Table 1). Four cohorts had Salmonella isolates of the same serovar identified in barn environment samples and finishing pig samples. Matching subtypes between environment and individual pig samples were identified in 3 of these 4 cohorts (75%) (Supplementary Fig. S1, Table 1). For example, subtype 24 of serovar Johannesburg was observed in environment and respective individual pig samples in cohorts 2 and 4 of site B (Fig. 1C, Table 1).
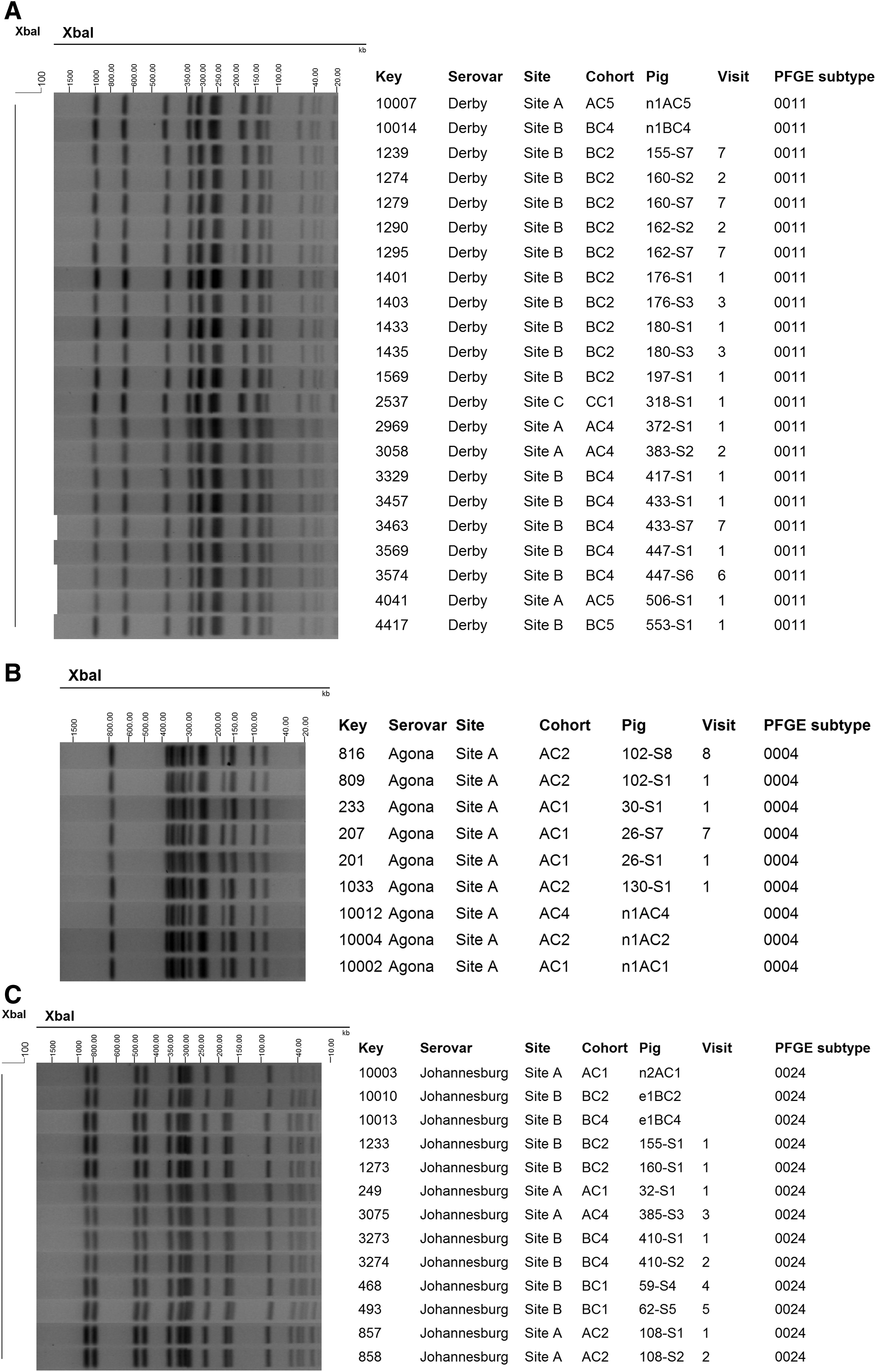
Partial dendrogram of pulsed-field gel electrophoresis (PFGE) subtypes of Salmonella among swine individual and pooled nursery fecal samples and barn environment and their association with serovar, site, cohort, sample type, and visit.
A large number of distinguishable PFGE patterns within serovar were identified within site and cohort, but distinguishable PFGE patterns within serovars were highly related, and the small band differences may represent small genetic changes to strains that occurred within the same farm. In site A, 12 PFGE subtypes were identified, 3 of which were common to the other two sites (Table 1). In site B, 12 distinguishable subtypes were observed. Four of those were common to sites A and C. In site C, two out of six patterns were identified in sites A and B as well (Table 1). In site A, two or three distinct subtypes of S. Agona were identified within each of three cohorts (cohorts 1, 2, and 4); two different subtypes of Salmonella Derby were identified within each of cohorts 2 and 5; and two different subtypes of Salmonella Johannesburg where identified in cohort 1 (Table 1). Similarly, in site B three subtypes of Salmonella Derby were found within cohorts 2 and 6, and two subtypes of S. Johannesburg within both cohorts 1 and 4 were identified (Table 1). In site C, two subtypes of Salmonella Derby were identified in cohort 3 (Table 1).
Within a serovar, indistinguishable PFGE subtypes were identified across sites and across cohorts within sites. For instance, PFGE subtype 11 of serovar Salmonella Derby was identified in all three sites (cohorts 4 and 5 of site A; cohorts 2, 4, and 5 of site B; cohort 1 of site C;) in consecutive cohorts and nonconsecutive cohorts (Fig. 1A, Table 1). The PFGE subtype 4 of serovar Salmonella Agona was observed in consecutive and nonconsecutive cohorts of the same site (cohorts 1, 2, and 4 of site A) (Fig. 1B, Table 1). Similarly, the PFGE subtype 25 of Salmonella Johannesburg (Fig. 1C, Table C) was observed in two sites, in consecutive and nonconsecutive cohorts (cohorts 1, 2, and 4 of site A; cohorts 1, 2, and 4 of site B).
Discussion
In this study, bacterial typing methods, serotyping, and PFGE were used to investigate genetic dynamics and spatiotemporal diversity of Salmonella in finishing swine. Genotyping results regarding the PFGE subtypes were similar to other studies (Gaul et al., 2007; Molla et al., 2010) in that the cluster analysis grouped the same serovars together.
Results from the PFGE genotyping indicated that the 107 isolates belonged to one of 25 subtypes distinguishable by at least one band difference. Classic standards for the interpretation of PFGE patterns suggest that three-band differences between patterns should be interpreted as “closely related” strains and six-band differences should be interpreted as “possibly related” (Tenover et al., 1995). However, more recent criteria suggest that only isolates displaying indistinguishable patterns should be included for cluster detection (Barrett et al., 2006). PFGE is a useful technique within inherent limitations. For instance, pattern similarity may not reflect genetic similarity between groups or isolates. Computer simulated genetic sequences and simulated chromosome restriction showed inadequate correlation between the calculated similarity using a single enzyme and the actual genetic similarity using the entire genome (Singer et al., 2004). A three-band difference between two strains could theoretically result from the single point mutation in a restriction site. Conversely, strains with indistinguishable patterns may be genetically different because bands of the same size do not always represent the same genetic material (Davis et al., 2003), and insertions or deletions that are smaller than 1–2% of the total chromosome size may not visibly alter the position of the band (Barrett et al., 2006). In this study, stringent criteria for defining subtypes was used to identify instances of between-pig and between-cohort transmission of clonal subtypes. The pattern differences observed in this study may be a result of the introduction of novel strains, very recent genetic changes, or ongoing transmission of strains that evolved within the production system at some point in the past. Therefore, clustering of isolates by pattern similarity should also be used in the interpretation because isolates with small pattern differences may represent within-farm transmission with accompanying genetic changes. Defining indistinguishable subtypes as those with at least 85% and 95% banding pattern similarity would result in 13 and 15 different subtypes, respectively, rather than 25.
Different combinations of Salmonella PFGE subtype within cohorts and sites were identified in the present study. The new subtypes were frequently detected in association with the introduction of a new group of finishing pigs in the barn or site. However, increases in the genetic diversity over time resultant from genetic changes in persistent strains within this production system could not be ruled out. Overwhelmingly, Salmonella serovars isolated from both nursery and individual pig samples within a cohort were of the same subtype, further strengthening our previous findings regarding the importance of Salmonella infection during the nursery phase for Salmonella prevalence and serovar type during the finishing phase (Pires et al., 2013a, 2014).
Furthermore, intermittent presence of the same subtype was observed among cohorts and sites, which suggests the circulation of indistinguishable subtypes in this production system during the entire 3-year study period. The persistence of the same genetic patterns within sites (mainly sites A and B) might reflect the presence of the same strains during the time period of follow-up in these two sites (Pires et al., 2013a) and the common nursery source of the last two cohorts (Pires et al., 2013a). Interestingly, the same subtype patterns of serovars S. Derby and S. Johannesburg were identified in sites A and B in cohorts with distinct nurseries. This might suggest a common infection source of Salmonella prior to arrival at the nursery (e.g., farrowing rooms, transportation trucks, and breeding stock). This production system had all-in/all-out management at nursery and finishing sites. During the study period, the system was changed from two farrowing sites to one, while the nursery sites remained unchanged (N1 and N2) (Pires et al., 2013a). Longitudinal studies conducted from farrowing to the finishing phase have demonstrated persistence of the same serovar and genotypes during the distinct production phases (farrowing, nursery and finishing stages) (Funk et al., 2001; Kranker et al., 2003; Dorr et al., 2009; Keelara et al., 2013). The present study has shown not only genotypic diversity within and between production stages, but also diversity within cohorts and pigs. These studies support that control of Salmonella targeted at finishing swine may require surveillance as early as the farrowing room or nursery.
Strain diversity in terms of serovars and subtypes was observed among sites. Sites A and B had the highest number of distinct PFGE subtypes (12). Such diversity might be explained by a combination of factors that predispose to entry and transmission of new strains. Despite the finishing sites belonging to the same production system and following the same standard operation procedures (e.g., feed sources, prophylactic and therapeutic protocols, biosecurity measures), differences related to management practices and barn building age and design might contribute to subtype persistence and diversity. Previous work has shown that the duration of Salmonella shedding was affected by management of sick animals (i.e., treatment with antibiotics). Pigs from cohorts with a higher proportion of treatments had a shorter Salmonella shedding duration (Pires et al., 2014). In addition, contact with wildlife (e.g., birds and rodents) was more likely to occur during the summer, due to the natural ventilation system and opening of lateral curtains. Wildlife contact might lead to increased transmission of certain serovars and strains to the pigs (Funk and Gebreyes, 2004; Andres et al., 2013; Andrés-Barranco et al., 2014). Therefore, these management differences (e.g., biosecurity, rodent control, treatment protocols) and differences in building characteristics might partly explain the genotypic diversity observed among sites and cohorts. The lower diversity of subtypes at site C might be explained by a combination of factors specific to this site, such as the different source of pigs, production type (weaning to finishing versus finishing), time disparity of the last four cohorts of pigs (followed up after the end of sampling at sites A and B), or due to a lower Salmonella prevalence at this site (Pires et al., 2013a).
Indistinguishable PFGE subtypes of isolates from environmental and pig origins were found in three out of four cohorts with available environment isolates, indicating the potential for Salmonella transmission via contaminated environment. Finishing barns were cleaned and disinfected between batches of pigs. Disinfectants (Synergize; Preserve International, Turlock, CA, USA, and VirskonS; Antee International UK, Sudbury, Suffolk, UK) were alternated following the standard operating procedure of the production system. These findings suggest that transmission occurs secondary to residual contamination within the barn and highlights the challenges of eliminating Salmonella from a barn environment (Gebreyes et al., 2004b; Mannion et al., 2007; Molla et al., 2010; Hernandez et al., 2013). A variety of subtypes were identified in individual pig samples, without any corresponding nursery or environment isolates. On one hand, this disparity might be due to the introduction of new subtypes by trucks, feed, or wildlife. Sampling of feed, trucks, or wildlife were not part of this study. On the other hand, it might be due to the diagnostic test characteristics (i.e., sensitivity) and consequently being unable to isolate Salmonella.
One of the limitations of this study was the inclusion of only one swine production company. Nevertheless, the selected production system is representative of the swine industry in the United States, and the serovars described in this study were common to other North American swine production systems. Another limitation of this study was related to the diagnostic tests and isolate selection. Related to the diagnostic test, despite fecal culture being considered the “gold standard,” it is an imperfect test (Love and Rostagno, 2008), which might misclassify the samples from different ecological niches and be unable to identify phenotyping and genotyping relatedness among different sample types. Moreover, pigs can be infected with more than one serovar (Funk, 2003; Garrido et al., 2014), and in the present study only one colony per fecal sample was selected for serotyping and PFGE analysis. In addition, in the present study, the repeated samples tested for PFGE were selected with an interval of 2 weeks or more, meaning the distinct subtypes identified in the same pig might be due to new infections occurring at different time points.
Conclusions
Genotypic diversity of Salmonella isolates within and across cohorts and sites was evident, as different combinations of subtypes were isolated in each cohort of pigs. New subtypes may result of the introduction of new strains, genetic changes, or ongoing transmission of evolved strains within the production system. The intermittent presence of the same subtype might reflect the establishment and adaptation of strains in different ecological niches and production stages. The presence of the same subtype over several cohorts of pigs was explained in part by persistence of the same subtype in nurseries and potential residual contamination from the barn. The diversity of subtypes elucidates the complexity of infection and circulation of Salmonella serovars and genotypes in commercial swine systems. Potential factors affecting adaptation and transmission within and among ecological systems (e.g., finishing pigs, nursery, and environment) should be further investigated. Understanding these factors may lead to more focused and cost-effective control programs through targeted interventions that have the most impact on Salmonella persistence and transmission during nursery and finishing phases.
Footnotes
Acknowledgments
Funding was provided, in part, by National Pork Board and U.S. Department of Agriculture Natural Resources Inventory, Epidemiologic Approaches to Food Safety grant 2007-01775. The authors thank the participating pork producers and their staff for collaborating in the investigation and staff and students at Michigan State University for their technical support.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.