
Abstract
To survey the diversity of plasmid-mediated β-lactamases, and the replicon types of the plasmids in Escherichia coli in the community, 95 multidrug-resistant E. coli isolates from stool samples of pigs in Korea from 2001 to 2011 were analyzed, and all isolates were confirmed as β-lactamase-producing bacteria, which mostly carried genes encoding TEM (100%), ACC (95%), SHV (62%), OXA (28%), CTX-M (26%), and NDM-1 (15%). Moreover, each isolate contained at least three β-lactamases genes on its plasmid. The polymerase chain reaction–based replicon typing method revealed that among the 18 replicon types, 15 types were identified in this work, and IncF groups (FIB; 100%, Frep; 99%) and HI1 (98%) type were predominant. Other replicon types such as Y, B/O, A/C, P, FIA, N, HI2, I1, and W were also detected; moreover at least three types were identified in each organism. In conclusion, a high prevalence of various types of β-lactamases genes were identified in E. coli from pigs in Korea, and these genes appeared to be mainly associated with IncF group, IncHI1, and IncY plasmids.
Introduction
β-
Among Inc type plasmids, particular types are more frequently identified in multidrug-resistant E. coli strains and these play a role in dissemination of resistance (Antunes et al., 2010). IncI1, IncN type plasmids encoding β-lactamases (CTX-M, CMY-2, SHV-12, and TEM-1) were identified in Europe and the United States (Moodley et al., 2009; Antunes et al., 2010), which meant that resistant plasmids could transfer between food-producing animals and humans (Antunes et al., 2010). Recently, IncFII, IncA/C, IncN, and IncI1 plasmids carrying ESBL genes are considered epidemic-resistant plasmids in Enterobacteriaeceae (Carattoli, 2009). For Inc type grouping, using polymerase chain reaction–based replicon typing (PBRT) methods (Carattoli et al., 2005), 18 pairs of primers (FIA, FIB, FIC, HI1, HI2, I1-Iγ, L/M, N, P, W, T, A/C, K, B/O, X, Y, and FII replicon) are designed for 5-multiplex and 3-simplex PCRs to detect the major plasmid Inc groups among Enterobacteriaceae (Couturier et al., 1988).
Bacteria have been evolving an ability to resist antibiotics along with the discovery of new antibiotics by transfer of antibiotic resistance genes (Toleman et al., 2006a). The transfer of antibiotic resistance genes among bacteria is mostly driven by transposons-bearing wide host ranging plasmids (Toleman et al., 2006b). Resistance genes are linked to special sequences, called common regions (CRs), which are often found beyond but close to the 3′ conserved sequences of class 1 integrons (Batchelor et al., 2005; Toleman et al., 2006a). These CRs, designated ISCR elements (Toleman et al., 2006b), are related and resemble an unusual class of insertion sequences (IS) such as IS91 (Batchelor et al., 2005; Toleman et al., 2006a). ISCR elements are transposed by a process called rolling-circle replication and, consequently, they can transpose adjacent DNA sequences found upstream of their transposase genes (Toleman et al., 2006a). Studies about the role of ISCR and other mobile elements in plasmids, which have a wide range of hosts and contain multiple antibiotic resistance genes such as tetA (tetracycline), sul1, 2 (sulfanilamide), FloR (chloramphenicol/florfenicol), and bla NDM-1, have revealed that mobilization and duplication of the β-lactamase genes are mediated by ISEcp1 (Arduino et al., 2002; Bratu et al., 2005; Call et al., 2010). Modules in these plasmids containing antibiotic resistance genes also contained mobile elements, including the Tn21 transposon and the ISCR2, ISCR16, and IS26 insertion sequences (Qi et al., 2008). Consequently, plasmids seem to use mobile elements to gain and accumulate different antibiotic resistance genes (Féria et al., 2002).
The primary aim of this work was to investigate the presence and the distribution of plasmid Inc groups and plasmid-mediated β-lactamase genes in multidrug-resistant E. coli isolates collected from food-producing animals (e.g., pigs) in Korea, and furthermore, to elucidate the phylogenetic relationships among β-lactamase gene-encoding plasmids by PBRT methods. Therefore, this research endeavors to better understand the relationship of these E. coli isolates in pigs and the possible transmission to humans.
Materials and Methods
E. coli isolates and susceptibility testing
A total of 469 E. coli isolates (329 from Chonnam, 50 from Gyeongsang, 50 from Chungcheong, and 40 from Gyeonggi Province) were obtained from fecal samples of pigs in Korea between 2001 and 2011. They were directly collected from fecal samples of rectums of farmed pigs (two or three randomly selected at each farm). All isolates were identified using standard biochemical testing (Gram staining, TSI, indole, methyl-red-Voges-Proskauer, and urease). Antibiotic susceptibility tests were performed for each isolate using the disk-diffusion method on Müller-Hinton agar with β-lactam antibiotics (penicillin, P; ampicillin, AM; amoxicillin, AMC; ticarcillin, TIC; ticarcillin/clavulanic acid, TIM; cephalothin, CF; cefepime, FEP; cefoxitin, FOX; cefotaxime, CTX; imipenem, IPM; aztreonam, ATM; ceftazidime, CAZ; ceftiofur, CFT) and non-β-lactam antibiotics (spectinomycin, SP; erythromycin, E; lincomycin, LI; lincomycin/spectinomycin, LI+SP; kanamycin, K; gentamycin, GM; amikacin, AN; neomycin, N; streptomycin, S; trimethoprim/sulfamethoxazole, SXT; ciprofloxacin, CIP; enrofloxacin, ENO; tetracycline, TE; tylosin, TYLO; and tiamulin, TIA). For colistin (CL), minimum inhibitory concentrations were determined by broth microdilution method. All performance and evaluation were followed the guidelines of the Clinical and Laboratory Standards Institute (CLSI, 2011).
Resistance transfer assay
Conjugation experiments were carried out with donor isolates, which were resistant to β-lactam antibiotics and sodium azide–resistant E. coli J-53 as the recipient strain. Transconjugants were selected on MacConkey agar plates containing sodium azide (100 μg/mL) and cefotaxime (1 μg/mL) (Kim et al., 2014). Not only was the β-lactamase production of transconjugants confirmed by PCR amplification and sequencing analysis, but also its replicon typing by the PCR-based method.
Molecular analysis of β-lactamase
Genomic DNA was extracted using PureLink™ Quick Genomic DNA Mini Kits (Invitrogen, Carlsbad, CA), according to the manufacturer's recommendations. All isolates were screened by PCR amplification and sequencing for the Ambler classes A, B, C, and D of β-lactamase-encoding genes such as bla KPC, bla IMP, bla VIM, bla NDM, and bla OXA, bla TEM, bla SHV, bla CTX, and bla ACC (Table 1).
Plasmid characterization
Plasmid DNA was extracted using PureLink™ Quick Plasmid Miniprep Kits (Invitrogen) according to manufacturer's recommendations. The presence of mobile genetic elements, associated with the dissemination of β-lactamase genes, was identified using primers (Table 1), which were designed using NCBI/Primer_BLAST (
Phylogenetic grouping
Genomic DNA was extracted using PureLink™ Genomic DNA mini kits (Invitrogen) according to the manufacturer's instructions. Tranconjugants were phylogenetically grouped into A, B1, B2, or D using a triplex PCR to detect genes chuA, yjaA, and a DNA fragment of TspE4C2 (Clermont et al., 2000). Primers used for triplex PCR are listed in Table 1.
Results and Discussion
Genotypes of β-lactam resistance
The status of β-lactam antibiotic resistance of the 469 donor E. coli isolates was as follows: P (460/469, 98%), CF (450/469, 96%), TIC (425/469, 91%), CFT (420/469, 90%), AMC (400/469, 85%), TIM (300/469, 64%), FEP (280/469, 60%), FOX (175/469, 37%), CTX (135/469, 29%), AMT (31/469, 7%), CAZ (19/469, 4%), and IPM (9/469, 2%). Transfer of β-lactam antibiotic resistance was detected in 95 isolates out of 469 E. coli isolates in this study. Among the 95 isolates, the distribution of resistance was as follows: P (94/95, 99%), CF (91/95, 96%), CFT (83/95, 87%), TIC (81/95, 85%), AMC (70/95, 74%), TIM (51/95, 54%), FEP (42/95, 44%), FOX (34/95, 36%), CTX (24/95, 25%), ATM (7/95, 7%), CAZ (5/95, 5%), and IPM (2/95, 2%).
By PCR amplification, multiple classes of β-lactamase genes were identified on the plasmids in all isolates. The distribution of plasmid-mediated β-lactamase genes of our multidrug-resistant E. coli isolates is shown in Figure 1. TEM type was identified on all plasmids, ACC type was detected in 90 (n = 95, 95%) and followed by SHV (62%), OXA (28%), CTX (26%), NDM (15%), KPC (13%), CMY-2 (7%), VIM (5%), and CARB type was just detected in one isolate. Two or more β-lactamase genes were present in all isolates; bla TEM + bla ACC was the most common combination (by 95%), followed bla ACC + bla TEM + bla SHV (51%), bla ACC + bla TEM + bla CTX (26%), bla ACC + bla TEM + bla OXA (25%), and bla ACC + bla TEM + bla NDM (15%).
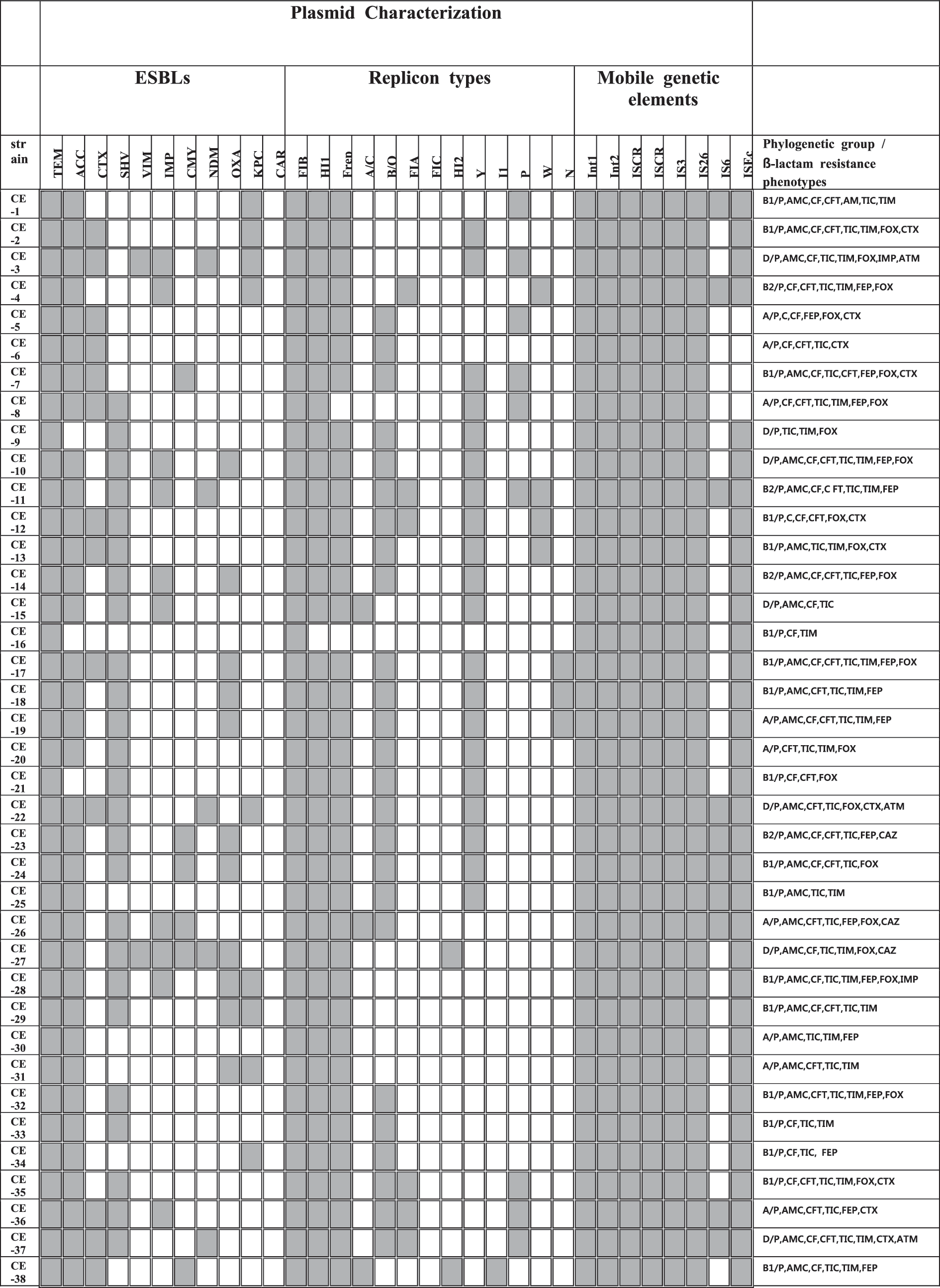
A heat-map summary of the results; β-lactamase present/absent profiles; the mobile genetic elements present/absent profiles; the non-β-lactamase antibiotics resistance profile for all 95 β-lactamase-positive Escherichia coli. Gray squares indicate feature present in that isolate, while white squares denote features that are lacking in the corresponding bacterial isolate.
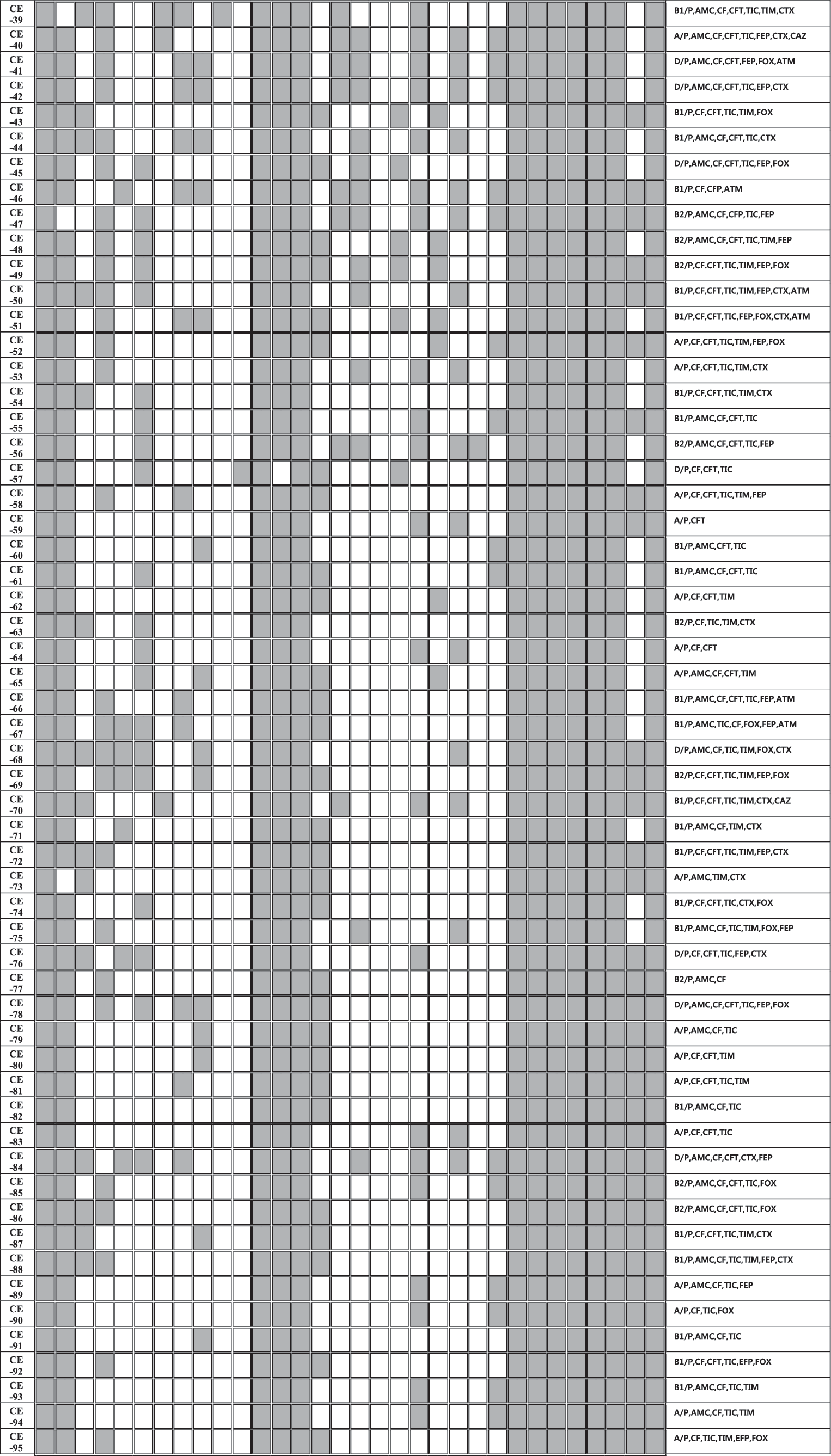
Among plasmid-encoded β-lactamases, TEM, CTX-M, and SHV types are widely distributed in human and animals (Carattoli, 2008). The prevalent genotypes vary by country; the TEM type is predominant in the United States and United Kingdom (Jacoby and Medeiros, 1991), and in France the SHV and TEM types were prevalent enzymes (Coque et al., 2008). In Korea, SHV, TEM, and CTX-M types were the main types detected, with the CTX-M type being the most prevalent (≈90%) ESBLs (Kim et al., 2005; Lee et al., 2009). In this study, three types were also detected, but main types were different as TEM, SHV, and ACC. Plasmid-mediated AmpC also has been founded globally (Philippon et al., 2002), and is increasing in livestock in Europe (Ewers et al., 2012; Hammerum et al., 2014). However, in Korea, reports of the prevalence of AmpC encoding plasmids in E. coli were rare, while plasmid-mediated AmpC in Salmonella isolates were reported in 2014 (Lee et al., 2014). Among 95 recipient plasmids in this study, the ACC type was identified at a high rate (95%). KPC-producing Enterobacteriaeceae also have disseminated globally (Bush, 2010), in addition, KPC-encoding plasmids are almost carrying other β-lactamase types, such as TEM, SHV, CTX-M, and CMY types (Bush, 2010). In this study, all 11 KPC-encoding plasmids were co-carrying other β-lactamases, such as TEM (11), ACC (10), CTX-M, SHV and IMP (4), OXA and NDM-1 (3), and VIM (1). Recently, IMP, VIM, and NDM-1 types, which are MBLs, are identified on transferable plasmids of Enterobacteriaceae worldwide (Bush, 2010). Of more than 80 ESBLs identified, 75% were plasmid-mediated enzymes (Bush and Jacoby, 2010). MBLs can hydrolyze all β-lactams without being inhibited easily. In addition, they often co-exist with other β-lactamases. Therefore, MBLs are global concerns (Bush, 2010). Ever since plasmid-mediated MBLs have been identified in 1994 (IMP in Japan, Osano et al., 1994), VIM types were isolated in Korea (in 2001; Yan et al., 2001). In this study, they were also identified in eight plasmids. The most prominent member of this class is NDM-1 type (Kumarasamy et al., 2010; Nordmann et al., 2011b), which was first discovered in a Klebsiella spp. isolate from an Indian patient previously hospitalized in New Delhi, India (Nordmann et al., 2011b). Since 2010, bacteria in the family Enterobacteriaceae encoding the bla NDM-1 gene have been detected in every continent except Central and South America (Nordmann et al., 2011b). Plasmids carrying the bla NDM-1 gene are diverse and associated with other β-lactamases genes (OXA types, VIM types, CTX-M types), AmpC cephalosporinase (CMY types), and aminoglycoside resistance genes (Kumarasamy et al., 2010; Nordmann et al., 2011b). Among our 95 isolates, 16 NDM-harboring plasmids were identified, and 15 plasmids of them co-carried the bla ACC gene, 12 plasmids co-carried the bla SHV gene, and 6 plasmids co-carried bla OXA gene. All 16 NDM-harboring plasmids co-carried more than 3 other β-lactamase genes with the NDM gene (Fig. 1). OXA types are one of the most common β-lactamases, which were first identified in a K. pneumoniae isolate from Turkey in 2003 (Poirel et al., 2004). Since then, they have been reported as a source of nosocomial outbreaks worldwide including Southern Europe and Africa, but not in the United States (Nordmann et al., 2011a). Identifications of OXA-producing Enterobacteriaceae are increasing in countries such as France, Germany, Spain, the Netherlands, and the United Kingdom (Nordmann et al., 2011a). In this study, the OXA-type β-lactamases gene was detected in 24 plasmids of the E. coli isolates.
Plasmid characterization
Distribution of replicon types of 95 β-lactamase gene-encoding plasmids is also shown in Figure 1. Among the known 18 replicon types, 15 types were detected in this study. The most common replicon type was FIB (n = 95, 100%), the second predominant type was Frep (n = 94, 99%), followed by HI1 (98%), Y (43%), B/O (35%), A/C (34%), P (26%), FIA (20%), N (19%), HI2 (8%), I1 (8%), and W (5%).
F family and HI1 types, which were predominant in this study, have been reported to be harbored by plasmids of E. coli, K. pneumoniae, and some Salmonella enterica, worldwide, and they were associated with ESBL genes, or plasmid-mediated quinolone or aminoglycoside resistance genes (Carattoli, 2009). IncF group plasmids (FIA, FIB, and Frep) have been associated with the spread of resistance genes such as bla KPC, quinolone, and plasmid-mediated aminoglycoside genes, and other Inc types (IncN, IncA/C, IncHI1, IncP) were also the vehicles for the spread of β-lactamase genes such as bla CTX, bla VIM, and bla CMY (Carattoli, 2009).
By PCR analysis, multiple patterns of mobile elements were detected in the plasmid collection studied here. Int1, Int2, ISCR1, ISCR2, IS3, and IS26 were detected in all isolates, and ISEcp1 (97%) and IS6 (46%) followed. Moreover, in all plasmids, at least five elements were co-existing. The transfer of their resistance traits in multidrug-resistant E. coli to other microorganisms could be explained with this result.
Susceptibility to non-β-lactamase antibiotics
All E. coli isolates were resistant to at least eight non-β-lactamase antibiotic agents. Among the 95 isolates, the distribution of antibiotic resistance was as follows: TIA, TYLO (95/95), E, S, N (94/95), K (93/95), TE (91/95), GM, SXT (78/95), CIP (75/95), NOR, ENO (70/95), AN (65/95), and CL (40/95).
Phylogenetic group analysis
E. coli strains can be divided into four main phylogenetic groups (A, B1, B2, and D) by phylogenetic analysis (Clermont et al., 2000). Virulent extraintestinal strains mainly belong to group B2 and, to a lesser extent, to group D (Clermont et al., 2000), whereas most commensal strains belong to groups A and B1. In this study, the less virulent group B1 and most commensal group A were predominant (43% and 26%), while more virulent groups, D and B2, were detected at the same rate as 15%, which was a similar result to human study (Jakobsen et al., 2010).
In conclusion, we demonstrated a high rate prevalence of plasmid-mediated β-lactamases in the fecal flora of pigs, which is an obvious risk for increasing prevalence in humans. Because E. coli is a zoonotic pathogen, genes encoding β-lactamase enzymes can be present in humans. The plasmids harboring these resistance genes co-carried mobile genetic elements, which can facilitate the transfer of drug-resistance genes among bacterial species. Consequently, the occurrence and spread of multidrug-resistant bacteria in humans and animals should be controlled by appropriate use of antibiotics in veterinary medicine and a prohibition on the subtherapeutic use of antibiotics in the animal industry.
Footnotes
Disclosure Statement
No competing financial interests exist.