
Abstract
The objective of this study was to characterize plasmids coharboring 16S rRNA methylases, bla CTX-M and virulence-associated genes in Escherichia coli and Klebsiella pneumoniae isolates from chickens in China. A total of 32 positive transconjugants exhibited coresistance to amikacin and cefotaxime in E. coli (24/281) and K. pneumoniae (8/93), and were identified by conjugation experiments and S1-pulsed-field gel electrophoresis. Polymerase chain reaction amplification assay detecting resistance genes showed that rmtB or armA gene accompanied with different bla CTX-M genes coexisted on 32 transferred plasmids. The bla CTX-M-98b gene was identified in chicken-derived E. coli and K. pneumoniae for the first time. The association between resistance genes and virulence genes was observed in the transferred plasmids; 68.8% (22/32) transferred resistance plasmids coharboring various virulence genes including traT, iutA, fyuA, msbB, and vagC genes with diverse proportions. Genetic stability tests revealed that 93.8% (30/32) transferred plasmids continued to exist in the host strain after continuous passage of 30 times in 15 days. Furthermore, 87.5% (28/32) conjugants showed no significant differences in growth rates compared with E. coli J53. Results of the growth competition assay showed that conjugants have low fitness cost, which indicated that there were no obvious negative effects on the host's growth. The combination of blaCTX-M -98b-rmtB-traT on 85-kb transferred IncF plasmids in E. coli, and bla CTX-M-14-rmtB-traT on 95-kb transferred IncF plasmids in K. pneumoniae were first identified in this study. These features of plasmids may contribute to the successful spread of resistance and virulence among pathogens of different sources and geographical origins.
Introduction
E
Furthermore, the pathogenicity of E. coli and K. pneumoniae is caused by specific virulence factors. It may cause more serious problem with the transmission of the plasmids when the virulence factors locate on plasmid (Calhau et al., 2013). More importantly, the rapid spread of conjugative plasmids coharboring resistance and virulence genes may lead to serious difficulties in the therapy and prevention of disease in animals. Some prevalent strains with plasmid-mediated resistance and virulence such as E. coli sequence type (ST) 131 has been reported (Woodford et al., 2009). However, the cause-and-effect relationships between resistance and virulence characteristics focus on the transferred plasmids is still largely unknown. The association of 16S rRNA methylase genes among ESBLs-producing strains with virulence genes on plasmids in E. coli and K. pneumoniae remains to be explored.
This study had three main objectives: (1) to study characteristics of transferred plasmids coharboring 16S rRNA methylase and bla CTX-M type genes in E. coli and K. pneumoniae in China; (2) to explore the association between resistance and virulence genes on transferred plasmids; and (3) to investigate the genetic stability and fitness cost of transferred plasmids.
Materials and Methods
Bacterial strains
A total of 281 E. coli and 93 K. pneumoniae isolates were collected from liver, spleen, heart, lung, gallbladder, and kidney of diseased or dead chickens in poultry farms among 13 provinces and municipalities in China (Anhui, Hubei, Henan, Sichuan, Shandong, Shanghai, Tianjin, Hebei, Jiangsu, Xinjiang, Heilongjiang, Shaanxi, and Ningxia) between January 2012 and December 2014. All isolates collected from different samples sources were identified by using an automated system (BD Diagnostic Systems, Sparks, MD).
Antimicrobial susceptibility testing
All isolates were examined for susceptibility to antimicrobial agents by the disk-diffusion method following the Clinical and Laboratory Standards Institute (CLSI) recommendations (CLSI, 2013). The following antimicrobials (Oxoid, Thermo Fisher Scientific, Basingstoke, UK) were tested: amikacin (AK), cefotaxime (CTX), ceftazidime (CAZ), gentamicin (CN), ampicillin/sulbactam (SAM), ciprofloxacin (CIP), levofloxacin (LEV), neomycin (N), florfenicol (FFC), norfloxacin (NOR), cefoxitin (FOX), doxycycline (DO), amoxicillin (AML), polymyxin B (PB), and imipenem (IPM). E. coli ATCC 25922 strain was used as the control strain. The results were interpreted according to CLSI instructions (CLSI, 2013). The isolates with coresistance to amikacin and cefotaxime were collected for further study.
Detection of 16S rRNA methylase and bla CTX-M genes
DNA templates were extracted by bacterial boiling method (Wei et al., 2015). The 16S rRNA methylase (Doi et al., 2007; Davis et al., 2010; Bueno et al., 2013) and bla
CTX-M genes (Zhang et al., 2014) were detected by polymerase chain reaction (PCR) amplification and information on the primers are listed in Supplementary Table S1 (Supplementary Data are available online at
Conjugation experiments and S1-pulsed-field gel electrophoresis (PFGE)
The transferability of plasmids was investigated by conducting mating-out assays (Poirel et al., 2010) using isolates with coresistance to amikacin and cefotaxime as donors and E. coli J53 as the recipient. Transconjugants were selected on MH agar plates containing amikacin (32 μg/mL), sodium azide (100 μg/mL) and cefotaxime (4 μg/mL). The positive transconjugants with coresistance to amikacin and cefotaxime were confirmed by antimicrobial susceptibility testing, and the donor strains and recipient strains were used as the control. The sizes of transferred plasmids were determined approximately by S1-PFGE (Sirichote et al., 2010).
Detection of resistance genes and virulence genes in transconjugants
DNA templates were obtained from boiled suspension of transconjugants cells. The resistance genes, which included 16S rRNA methylase and bla CTX-M genes in transconjugants, were detected by using the same method mentioned above. The presence of virulence genes in transconjugants was investigated by PCR amplification. Specific primers for 17 virulence genes (Johnson et al., 2000; Gow et al., 2009), including traT, msbB, vagC, fyuA, iutA, vagD, katP, espP, papAH, papC, papEF, hlyA, kpsMTIII, fimH, eae, stx1, and stx2, are shown in Supplementary Table S2, and the PCR reaction conditions were carried out with the protocol consisted of 4 min at 94°C followed by 30 cycles of 94°C for 30 s, annealing temperature for 30 s, 72°C for 30 s, and a final extension at 72°C for 10 min. The positive PCR products were identified by DNA sequencing.
Molecular typing of donor strains and transferred plasmids
The genetic relatedness of donor isolates was investigated by XbaI PFGE analysis, as previously described (Habeeb et al., 2014). PCR detection of the ST131 isolates was conducted according to a rapid detection method reported previously (Clermont et al., 2009). MLST of E. coli was conducted to confirm the identification of ST131 type strains (Tartof et al., 2005). PCR-based replicon typing (PBRT) for transferred plasmid harboring bla CTX-M and 16S rRNA methylase genes was analyzed according to Carattoli et al. (Carattoli et al., 2005).
Genetic stability, growth curve, and growth competition experiments
Stability of transferred plasmids was investigated by continuous passage of conjugants strains for 30 times in 15 days to explore whether the transferred plasmids could disappear at certain generation. Growth curve of conjugants harboring resistance plasmid and recipient E. coli J53 was determined and the growth competition experiments between conjugants and recipient strain were carried out based on the method reported previously (San Millan et al., 2010; Xu et al., 2015).
Results
Antimicrobial susceptibility
In total, 36 of the 281 (12.8%) isolates of E. coli and 15 of the 93 (16.1%) isolates of K. pneumoniae exhibited coresistance to amikacin and cefotaxime. The antimicrobial resistance rates of E. coli and K. pneumoniae were as follows, respectively: amikacin (100%/100%), cefotaxime (100%/100%), ceftazidime (52.8/40.0%), gentamicin (88.9%/86.7%), ampicillin/sulbactam (36.1%/53.3%), ciprofloxacin (52.8%/46.7%), levofloxacin (66.7%/46.7%), neomycin (86.1%/80.0%), florfenicol (44.4%/53.3%), norfloxacin (63.9%/53.3%), cefoxitin (25%/33.3%), doxycycline (52.8%/60%), amoxicillin (83.3%/86.7%), polymyxin B (8.3%/0), and imipenem (0/0).
16S rRNA methylase genes and bla CTX-M genes
The results showed that 72.2% (26/36) carried rmtB gene and 11.1% (4/36) carried armA gene in E. coli isolates, 8.3% (3/36) co-carried rmtB and armA genes, and 8.3% (3/36) co-carried rmtB and rmtD gene. For K. pneumoniae, 73.3% (11/15) carried rmtB gene and 20% (3/15) carried armA gene, and 1 isolate co-carried rmtB, armA, and rmtD gene simultaneously.
The detection of bla CTX-M genes revealed that all strains harbored either CTX-M-1 or CTX-M-9 group genes. The bla CTX-M-98b gene was identified for the first time both in chicken-derived E. coli (4/24) and K. pneumoniae (2/8). For E. coli isolates, 22.2% (8/36) carried bla CTX-M-79 gene, 19.4% (7/36) carried bla CTX-M-15 gene, 13.9% (5/36) carried bla CTX-M-14 gene, and bla CTX-M-3, 65, 55 were also found, which implied diverse distribution of bla CTX-M type genes. For K. pneumoniae isolates, 26.7% (4/15) carried bla CTX-M-14 gene, 20% (3/15) carried bla CTX-M-15 gene and 13.3% (2/15) carried bla CTX-M-79 gene, respectively.
Transferability of resistance plasmids and S1-PFGE
Results showed that 24 of 36 (66.7%) E. coli and 8 of 15 (53.3%) K. pneumoniae were transferred into E. coli J53 successfully, which were further confirmed by S1-PFGE. Overall, there were six kinds of plasmids with various sizes range from approximately 35 kb to 194 kb (Table 1).
A group, IncF type and carried virulence gene; B group, non-IncF type and carried virulence gene; C group, IncF type with no virulence gene; D group, non-IncF type with no virulence gene.
Detection of resistance genes and virulence genes in transconjugants
Results revealed that rmtB or armA genes existed in 32 transconjugant strains. In addition, the rmtB gene in particular maintained dominance, with 91.7% (22/24) derived from E. coli and 87.5% (7/8) derived from K. pneumoniae, respectively. For bla CTX-M genes, bla CTX-M-98b, 79, 15, 14, 3, 65, 55 and bla CTX-M-98b, 14, 79, 15 were detected in transconjugants derived from E. coli and K. pneumoniae, respectively (Table 1).
The results of virulence genes detection showed that 22 of the 32 (68.8%) transconjugants carried at least 1 virulence gene, including traT, iutA, fyuA, msbB, and vagC genes. Our results showed high frequency with 50% (11/22) of the transconjugants carrying traT gene, 9.1% (2/22) carrying fyuA gene, and 4.5% (1/22) carryig iutA gene, respectively. Interestingly, up to eight transconjugants carried two different virulence genes simultaneously. The transconjugants of SC54-4, SC56-4, SC53-10, and SC55-6 coharbored fyuA and iutA genes. Furthermore, HeN36-2 and SC23-1 coharbored iutA and traT genes, SD16-2 co-carried traT and msbB genes, and HuB14-3 coharbored iutA and vagC, respectively (Table 1).
The association between resistance genes and virulence genes was observed in the transferred plasmids: 68.8% transferred resistant plasmids coharboring various virulence genes, and traT gene emerged with high frequency on transferred plasmids. It was worth noting that the occurrence of combination of bla CTX-M-98b-rmtB-traT on plasmids in E. coli, and another combination of bla CTX-M-14-rmtB-traT on plasmids in K. pneumoniae were identified for the first time in this study.
Molecular typing of donor strains and transferred plasmids
A dendrogram of XbaI-PFGE patterns shows the relatedness of 24 E. coli and 8 K. pneumoniae isolates (Figs. 1 and 2), which indicated that all 32 isolates belonged to nonhomologous strains. MLST data showed that there were many sequence types such as ST98, ST987, and ST2464 (data not shown) and there was no isolate that belonged to ST131 type among 24 E. coli, which may be explained by previous report that E. coli ST131 was strongly associated with fluoroquinolone resistance but not for aminoglycosides (Matsumura et al., 2013).

XbaI–pulsed-field gel electrophoresis (PFGE) typing for 24 Escherichia coli isolates with resistance of cefotaxime and amikacin and transferred plasmid.
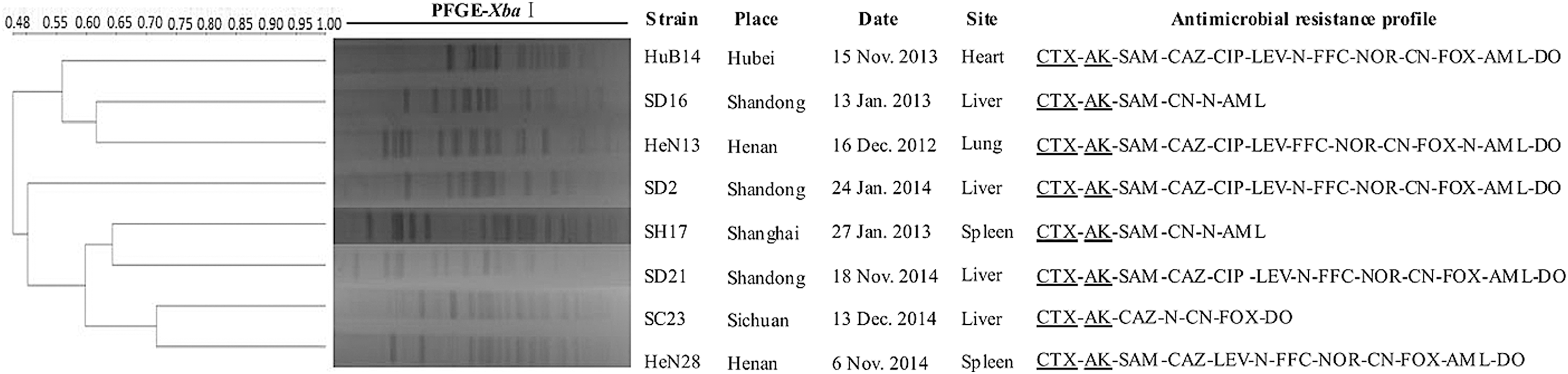
XbaI–pulsed-field gel electrophoresis (PFGE) typing for eight Klebsiella pneumonia isolates with resistance of cefotaxime and amikacin and transferred plasmid.
The results of PBRT showed that the IncF types plasmids accounted for 54.2% (13/24) in E. coli and 62.5% (5/8) in K. pneumoniae remained dominant compared to other replicon types (Table 1). In addition, we found that IncF plasmids had a high rate of 72.7% (16/22) in transferred plasmids coharboring resistant and virulence genes, which was consistent with the previous reports that IncF types plasmids existed frequently and were associated with resistance and virulence determinants (Villa et al., 2010). Furthermore, the replicons type of IncN, IncT, IncHI2, and IncP were also found in other transferred plasmids.
Genetic stability, growth curve, and growth competition experiments
The continuous passage of conjugants strains was carried out to investigate the stability of transferred plasmids in the recipient strain. Results showed that 93.8% (30/32) transferred plasmids existed in the host strain after continuous passage of 30 times in 15 days, which indicated that transferred plasmids were able to be kept steadily in the host strain. Growth curves of conjugants strains and E. coli J53 showed that 87.5% (28/32) of conjugants had no obvious negative effects on J53 growth, which indicated that the conjugants obtained remarkable transmission capacity. Results of the growth competition experiments showed that the fitness cost of conjugants may be the reason for the loss of transferred plasmids. However, the fitness cost of 14 conjugants, which were under 0.7, indicated that there were no obvious harmful effects on the growth of host J53 strain when there was competitive growth with wild J53 (Table 1).
Discussion
The characteristics and association of plasmids coharboring 16S rRNA methylases, bla CTX-M, and virulence genes in E. coli and K. pneumoniae isolates from chickens were investigated in this study. Our results showed that the isolates' coresistance to amikacin and cefotaxime was more prevalent among K. pneumoniae and E. coli. The isolates also showed high resistance rates to other aminoglycosides (gentamicin and neomycin), but relatively low resistance rates were observed for ciprofloxacin and florfenicol than shown in a previous report in China (Rao et al., 2014). We found that rmtB or armA genes existed on the transferred plasmid in all 32 conjugants strains and rmtB particularly maintained dominance. One possible consequence is that it may be beneficial to promote the spread of rmtB and armA genes, thereby accelerating the transmission of resistance to amikacin with the transfer of plasmids. These results were consistent with a previous study in China indicating that dominant rmtB and armA genes existed in multidrug-resistant (MDR) E. coli (Du et al., 2009) and the rmtB gene was the most prevalent 16S rRNA methylases gene in E. coli isolated from chickens in China (Xia et al., 2011). The bla CTX-M-98b gene, found in ducks and geese lately in China (Rao et al., 2014), was identified in chicken-derived E. coli and K. pneumoniae for the first time, which indicated that the bla CTX-M-98b gene has emerged and spread in some chicken farms in China. Furthermore, there were high proportions of rmtB and bla CTX-M genes that coexisted on transferred plasmids in this study, and the combination of rmtB-bla CTX-M-98b in E. coli and rmtB-bla CTX-M-14 gene in K. pneumoniae is cause for concern regarding the rapid transmission of MDR E. coli and K. pneumoniae that carry various resistance genes.
The location of virulence-associated genes in mobile genetic elements could facilitate the spread of virulence factors within bacterial communities. We found that 68.8% transferred resistant plasmids were coharboring virulence genes simultaneously. The virulence gene of traT has a resistance-associated outer membrane protein, which may contribute to serum resistance (Shin et al., 2014). It was notable that the high frequency rate of traT gene with 50% was confirmed in this study, which indicated its wide existence in E. coli and K. pneumoniae in China, which is largely overlooked at present. A previous study also showed that the traT gene was significantly more common in strains producing the blaCTX-M-1 group and the blaCTX-M-9 group ESBLs compared with CTX-M nonproducers (Pitout et al., 2005). The iutA and fyuA genes also showed a high frequency rate and could coexist with other virulence genes. The iutA gene is an aerobactin receptor belonging to siderophores and could be used as a predictor of pathogenicity, and the fyuA gene is a Yersinia bactin receptor (Mnif et al., 2013); both iutA and fyuA are encoding siderophores involved in bacteria pathogenicity (Rogers et al., 2011). Furthermore, The virulence-associated gene vagC has a possible function in stable plasmid inheritance (Szczepanowski et al., 2005). The vagC gene was found on one transferred resistant plasmid of HuB14-3 strain. The msbB gene encodes myristoyl transferase, which contribute the virulence to pathogens (Claes et al., 2014). The msbB gene existed on transferred plasmids of SD16-2. These evidences significantly supported the conclusion that various virulence genes located on plasmids could spread with the transfer of mobile genetic elements, and a part of them have emerged with high frequency in E. coli and K. pneumoniae in China.
The association between antimicrobial resistance and virulence of bacteria remains largely unknown. Recently the most commonly accepted view is that there was a direct correlation between quinolones resistance and loss of virulence (Da Silva et al., 2012). However, little was known about the association between 16S rRNA methylases genes and virulence genes located on plasmids. Our results showed that 68.8% (22/32) transferred plasmids coharboring resistance and virulence genes, which indicated specific virulence genes could exist on resistant plasmid carrying the rmtB or armA gene. Actually, the virulence plasmid combination with the antimicrobial resistance was not common (Bercot et al., 2008), though some plasmids have been proved to be coharboring resistance genes and virulence genes such as pRSB107 (Szczepanowski et al., 2005) and pO26-CRL (Venturini et al., 2010). We found there were five virulence genes located on plasmids with a remarkable frequency rate, which reflected in the fact that it has become a common phenomenon for there to exist plasmids coharboring resistance and virulence genes in E. coli and K. pneumoniae in China. The reason may be because these virulence genes are involved in bacteriocins, siderophores, cytotoxins, or adhesion and were necessary to hosts that are carrying resistance genes (Thomas et al., 2005), and the association between virulence genes and class 1 integrons related to antimicrobial resistance has been proved (Koczura, et al., 2013). In addition, results showed that IncF plasmids possessed a high rate in transferred plasmids coharboring resistant and virulence genes, which may be interpreted as meaning that IncF plasmids are likely to be beneficial to the bacterial host by providing virulence and antimicrobial resistance determinants but also encode several addiction systems that guarantee their maintenance and stability in the host cell (Carattoli, 2011). The combination of blaCTX-M -98b-rmtB-traT on 85-kb transferred IncF plasmids in E. coli, and bla CTX-M-14-rmtB-traT on 95-kb transferred IncF plasmids in K. pneumoniae was first identified in this study. The transmission of these specific plasmids has the potential to further expand the threat of multidrug-resistant pathogens. However, the 17 virulence genes that were investigated in this study still only reflected parts of the virulence characteristics of the transferred plasmids, and more indexes need to be involved in further study.
Genetic stability tests revealed that 93.8% of transferred plasmids were not lost from the host strain, which indicated high genetic stability of these plasmids in hosts. Some conjugants (87.5%) showed no significant differences in growth rates compared with J53, which indicated that the conjugants obtained remarkably stable transmission capacity. Previous study also showed that different combinations of bla CTX-M genes and 16S rRNA methylase genes have similar growth rates (Shin et al., 2014). The fitness cost of 14 conjugants was <0.7, which indicated that there were no obvious negative effects on the growth of the host strain. Previous study showed that the virulence and fitness have an opposite relationship (Beceiro et al., 2013). Our results showed that there were no significant effects of virulence genes and resistant genes on the fitness cost. These features may contribute to the successful spread of specific transferred plasmids among bacteria of different sources and geographical origins.
Conclusions
A total of 32 transferred plasmids root in E. coli and K. pneumoniae coharboring resistance genes and virulences genes were identified in this study. The rmtB and armA genes existed on the transferred plasmids and rmtB was kept particularly dominant. The bla CTX-M-98b gene was first identified in chicken-derived E. coli and K. pneumoniae in this study. Five kinds of virulence genes including traT, iutA, fyuA, msbB, and vagC genes located on the transferred resistant plasmids were confirmed and there were associations between virulence genes and resistance genes. It was worth noting that the occurrence of combination of bla CTX-M-98b-rmtB-traT on 85 kb transferred IncF plasmids in E. coli, and another combination of bla CTX-M-14-rmtB-traT on 95 kb IncF plasmids in K. pneumoniae.
Footnotes
Acknowledgments
This work was supported by “973” National Basic Research Program of China (project number 2013CB127200), Earmarked Fund for Modern Agro-industry Technology Research System (grant number CARS-41-K09), Special Fund for Agro-scientific Research in the Public Interest of China (201303044), the General Program of National Natural Science Foundation of China (31100102) and Science & Technology Pillar Program in Sichuan Province (grant number 2013NZ0025, 13ZC2578 and 2012GZ0001-1).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.