
Abstract
Pomegranate rind has been found to inhibit numerous pathogens, mostly attributed to its tannin fraction. The present study was conducted to investigate the quorum sensing (QS) inhibition effect of tannin-rich fraction from pomegranate rind (TFPR) by using an indicator strain Chromobacterium violaceum. Meanwhile, its effect on biofilm formation and motility of Escherichia coli was evaluated. It was shown that TFPR inhibited QS-regulated violacein pigment production. Biofilm formation and motility of E. coli were also hindered by TFPR. Transcriptional analysis further showed that TFPR repressed expressions of curli genes (csgB and csgD) and various motility genes (fimA, fimH, flhD, motB, qseB, and qseC). Our findings indicated that TFPR has potential application for controlling E. coli contaminations or biofilms in the food industry.
Introduction
B
Searching for nontoxic QS inhibitors from nature draws considerable attention from researchers. Many components have been found to act as bacterial QS inhibitors and attenuate the QS-dependent infection process (Kalia, 2013). Pomegranate (Punica granatum), an ancient fruit in the Middle East, which has been consumed throughout the world for thousands of years, is being widely promoted for its various biological functions (Johanningsmeier and Harris, 2011). Anthocyanins and hydrolyzable tannins, especially ellagitannins, are the major phytochemical components of pomegranate (Tzulker et al., 2007). Beneficial effects of pomegranate products have been reported in several diseases, such as cancer (Vadhanam et al., 2013), cardiovascular diseases (Johanningsmeier and Harris, 2011; Ismail et al., 2012), diabetes (Banihani et al., 2013) and microbial infection (Kisla and Karabiyikli, 2013; Li et al., 2014a).
Escherichia coli is one of the most comprehensively studied bacteria. Several E. coli clones have evolved the ability to cause numerous diseases (Blattner, 1999; Kaper et al., 2004; Wu et al., 2014). Some E. coli strains are capable of attaching to a substance, an interface, or each other and forming a community called biofilm (Costerton et al., 1995). In the biofilm, the cells demonstrate stronger resistance against different environmental stresses (e.g., disinfectants and antibiotics) (Naves et al., 2008; Vestby et al., 2009). E. coli utilize SdiA (a homologue of LuxR transcriptional regulator) to sense AHLs produced from other species of microorganisms due to lack of AHLs synthesis genes (Yang et al., 2006; Yao et al., 2007). It has been found that sdiA gene is associated with biofilm formation of E. coli (Sharma et al., 2010; Shimada et al., 2013; Laverty et al., 2014).
In the present study, we measured the QS inhibitory effect of the tannin-rich fraction from pomegranate rind (TFPR) by a QS indicator strain C. violaceum. In addition, we also evaluated the effect of TFPR on QS-associated phenotypes in E. coli such as biofilm formation and motility. Moreover, quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR) was used to determine certain gene expression in E. coli grown in media with or without TFPR.
Materials and Methods
Bacterial strains and materials
Wild-type C. violaceum ATCC 12472 (American Type Culture Collection, Manassas, VA), which has been widely used as a QS indicator strain, was used for QS inhibition assays (El-Gohary and Shaaban, 2013; Burt et al., 2014). E. coli ATCC 25922 was used for both biofilm formation and motility study. Three additional strong biofilm-producing E. coli strains, EC464, EC592, and EC638, were also used in biofilm formation study, which were taken from our laboratory collection and were isolated from retail chicken (Wu et al., 2014). All strains were stored at −80°C in Luria–Bertani (LB) broth (Beijing Land Bridge Technology Co., Ltd., Beijing, China) supplemented with 30% glycerine and recovered in LB broth prior to use. The TFPR was prepared and purified as previously described, which consisted of 64.2% punicalagin (Li et al., 2014a). The dried TFPR was stored at 4°C prior to use.
QS inhibition assay
The effect of TFPR on QS-regulated production of violacein was investigated as previously described with some modification (Taganna et al., 2011; Zhu et al., 2011; Truchado et al., 2012), using wild-type violacein pigment-producing strain C. violaceum ATCC 12472. One hundred microliters of overnight culture of C. violaceum ATCC 12472 in LB broth (optical density (OD) of 0.5 at 600 nm) was added to autoclaved flasks with 9.9 mL of fresh LB broth containing different amount TFPR (at final concentrations of 0.625, 0.312, and 0 mg/L). All flasks were subsequently incubated at 30°C for 48 h. Then 2 mL of culture was taken to determine cell counts by serial dilution plating. The rest of the culture was centrifuged at 18,000 × g for 5 min to precipitate cells and insoluble violacein. After the supernatant was discarded, 3 mL of DMSO was added to the pellet and the solution was vortexed vigorously for 30 s to completely solubilize violacein. The solution was centrifuged at 18,000 × g for 5 min, and the absorbance of violacein-containing supernatants was read with a SmartSpec Plus spectrophotometer (Bio-Rad, Hercules, CA) at a wavelength of 585 nm.
Antimicrobial and antibiofilm assays
Antimicrobial effect of TFPR on E. coli was determined using a broth microdilution method according to the Clinical and Laboratory Standards Institute (CLSI, 2012). Briefly, 10 μL of E. coli overnight culture was inoculated to each well containing 190 μL of LB broth with TFPR (TFPR concentrations of 0, 0.078, 0.156, 0.312 and 0.625 mg/mL) in 96-well flat-bottom polystyrene plates (Costar, Corning, NY). The plates were then incubated at 37°C for 24 h.
Biofilm formation assay was performed following the method of Naves et al. (Naves et al., 2008) with slight modifications. E. coli was incubated at 37°C with shaking (100 rpm) in LB broth without NaCl (tryptone 10 g/L and yeast extract 5 g/L) overnight. The overnight culture was diluted with fresh LB broth without NaCl to OD600 value of ≈1.0, and was mixed with fresh LB broth without NaCl (1:99) containing different amount TFPR (final concentrations of 0, 0.781, 0.156, 0.312, and 0.625 mg/mL) and N-hexanoyl-l-homoserine lactone (final concentration of 1 μmol/mL; Sigma-Aldrich, St. Louis, MO). Also, 150 μL of diluted culture was transferred to 96-well flat-bottom polystyrene plates (Costar) per well. Sterile LB broth without NaCl containing different concentrations of TFPR was used as control. All plates were incubated at 28°C for 48 h without shaking. Cell growth was measured through ODs at a wavelength of 630 nm using a microplate spectrophotometer (model 680; Bio-Rad). Then plates were emptied and gently rinsed three times using distilled water, and all plates were oven dried at 55°C for 20 min. The biofilm was then stained with 160 μL of 1% crystal violet (Tianjin Kermel Chemical Reagent Co., Ltd., Tianjin, China) for 5 min and excess dye was rinsed three times with distilled water. After 20 min of oven drying at 55°C, 160 μL of ethanol was added and incubated for 10 min before ODs were obtained at a wavelength of 570 nm. Specific biofilm formation (Niu and Gilbert, 2004) was calculated by attached and stained bacteria (OD570nm) normalized with cell growth (OD630).
Microscopy
Overnight culture of E. coli ATCC 25922 was diluted with fresh LB broth without NaCl to OD600 value of ≈1.0, and was mixed with fresh LB broth without NaCl (1:99) with or without TFPR. One milliliter of dilution was seeded into a sterile tube, pre-placed on a glass slide, and incubated at 28°C for 48 h without agitation. After incubation, suspended culture was discarded and rinsed with phosphate-buffered saline to remove suspended cells remaining on slides and all slides were air dried for 1 h. Biofilm was stained using 4'-6-diamidino-2-phenylindole solution in the dark for 20 min and washed with phosphate-buffered saline (Capita et al., 2014; Kim and Park, 2013). Biofilm was examined under a digital inverted fluorescence microscope (EVOS, Advanced Microscopy Group, Bothell, WA).
Motility inhibition assays
Swimming
Swimming motility was evaluated as previously described (Vestby et al., 2014) with some modifications. LB broth with 0.3% (wt/vol) agar was autoclaved then kept in a water bath at 55°C and N-hexanoyl-l-homoserine lactone (1 μmol/mL) was added. TFPR was added to the medium to achieve final concentrations of 0.312 and 0.625 mg/mL, and medium without TFPR was used as control. All medium were poured into six-well plates (Costar) 2 h prior to use. One microliter of overnight cultures of E. coli ATCC 25922 was stabbed to the center of the semisolid agar halfway down. All plates were incubated at 37°C for 12 h, and photos were taken.
Swarming
For swarming, LB broth containing 0.5% (wt/vol) agar, 1 μmol/mL N-hexanoyl-l-homoserine lactone, and 0, 0.312, or 0.625 mg/mL TFPR were poured into six-well plates 2 h prior to use. One microliter of overnight cultures of E. coli ATCC 25922 was incubated at the center of the semisolid agar. Plates were incubated at 37°C for 24 h, and photographs were taken. Swarming area was measured using Adobe Photoshop CS6 version (Adobe, San Jose, CA).
Quantitative RT-PCR
Bacterial cells of E. coli ATCC 25922 treated with different concentrations of TFPR were collected when cells grew to an OD600 of ≈1. An RNA purification Bacteria kit (Tiangen, Beijing, China) was employed to extract RNA in different samples according to the manufacturer's instructions. RNA samples were then reverse transcribed into cDNA with a TaKaRa PrimeScript reagent kit (Perfect Real Time; TaKaRa, Kyoto, Japan) according to the manufacturer's directions, and all cDNA samples were kept at −20°C prior to use. The concentration and quality of RNA and cDNA were determined with a nucleic acid and protein mini-spectrophotometer (Nano200; Hangzhou Allsheng Instruments Co., Ltd., Hangzhou, China).
RT-PCRs were performed with the IQ5 system (Bio-Rad) in a 25-μL SYBR Premix Ex TaqII (TaKaRa) system according to the manufacturer's instructions. The cycling conditions were as follows: 1 cycle of 95°C for 30 s, 45 cycles of 95°C for 5 s, and 60°C for 30 s, and a dissociation step of 95°C for 15 s and 60°C for 30 s. After completion of 45 PCR cycles, melt curve data were generated. Gene-specific primers were used (Table 1) and rpoA was used as a housekeeping control (Table 1). Data analysis was based on the relative quantitation method (Livak and Schmittgen, 2001).
p < 0.05.
p < 0.01.
F, forward; R, reverse.
Statistical analysis
All experiments were performed in triplicate. Independent Student t tests were used for statistical analysis by SPSS19.0 (IBM, New York, NY). A p value of <0.05 was considered to be statistically significant.
Results
Inhibition of QS
In general, concentration of TFPR and pigment production of C. violaceum showed an inverse relationship (Fig. 1). Pigment production was decreased when TFPR was used at 0.312 or at 0.625 mg/mL (around 49.65 and 29.88% of the control level, respectively). In addition, cells number of C. violaceum was determined using serial dilution plating method. It showed that TFPR at concentrations used had no significant antimicrobial effect on C. violaceum (data not shown).
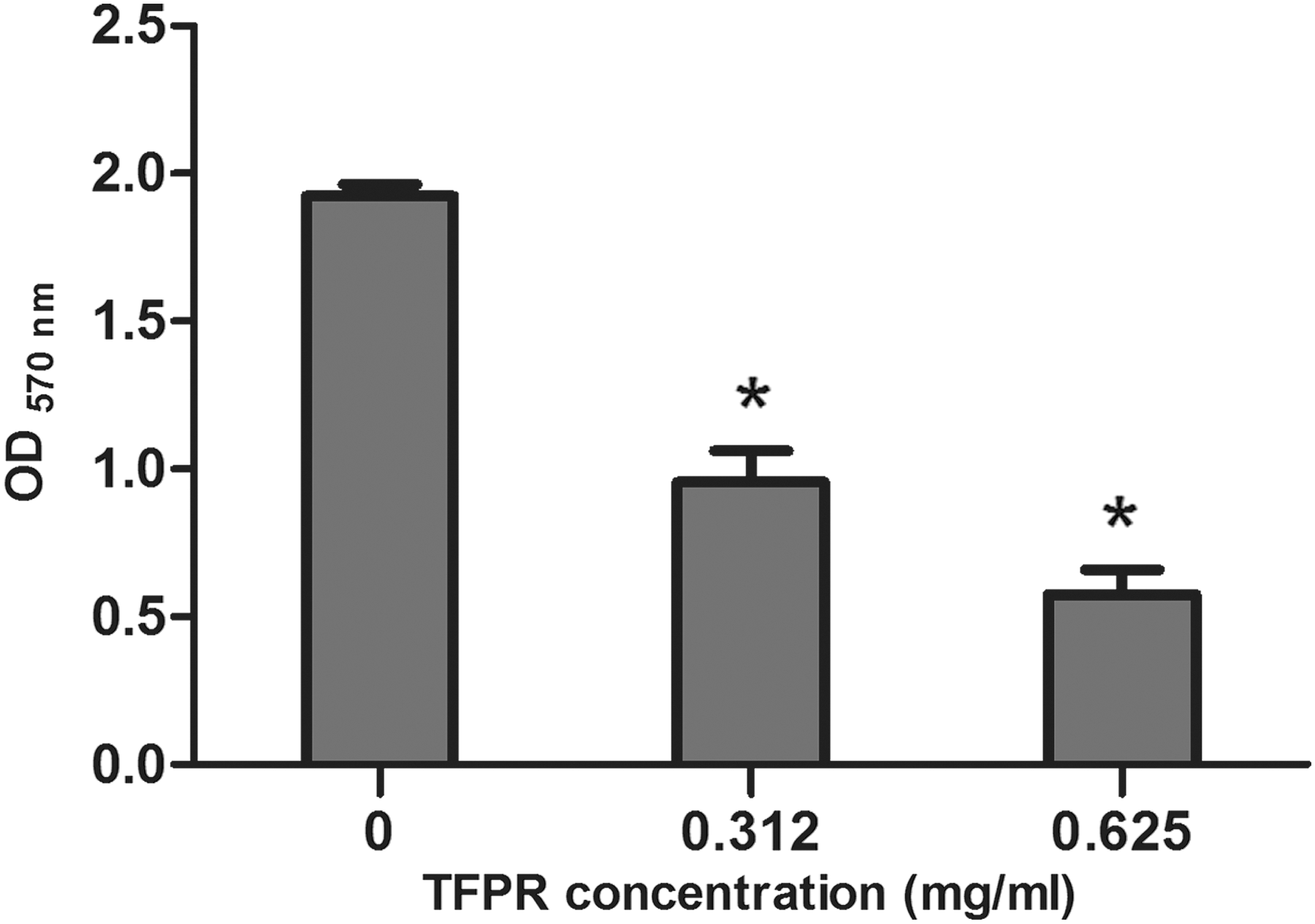
Inhibition of violacein production by Chromobacterium violaceum at different concentrations of tannin-rich fraction from pomegranate rind (TFPR). *p < 0.01.
Effect of TFPR on E. coli bacterial growth and biofilm formation
TFPR showed no apparent antimicrobial activity against E. coli at concentrations used in this study using a broth microdilution method (data not shown). Then the crystal violet assay was used to evaluate the effect of TFPR on biofilm formation of E. coli. As shown in Figure 2, a marked and dose-dependent inhibition of biofilm formation of 4 different E. coli strains by TFPR was observed. For E. coli ATCC 25922, biofilm formation was inhibited by 64.50% at 0.625 mg/mL of TFPR and inhibited by 17.78% at 0.078 mg/mL.
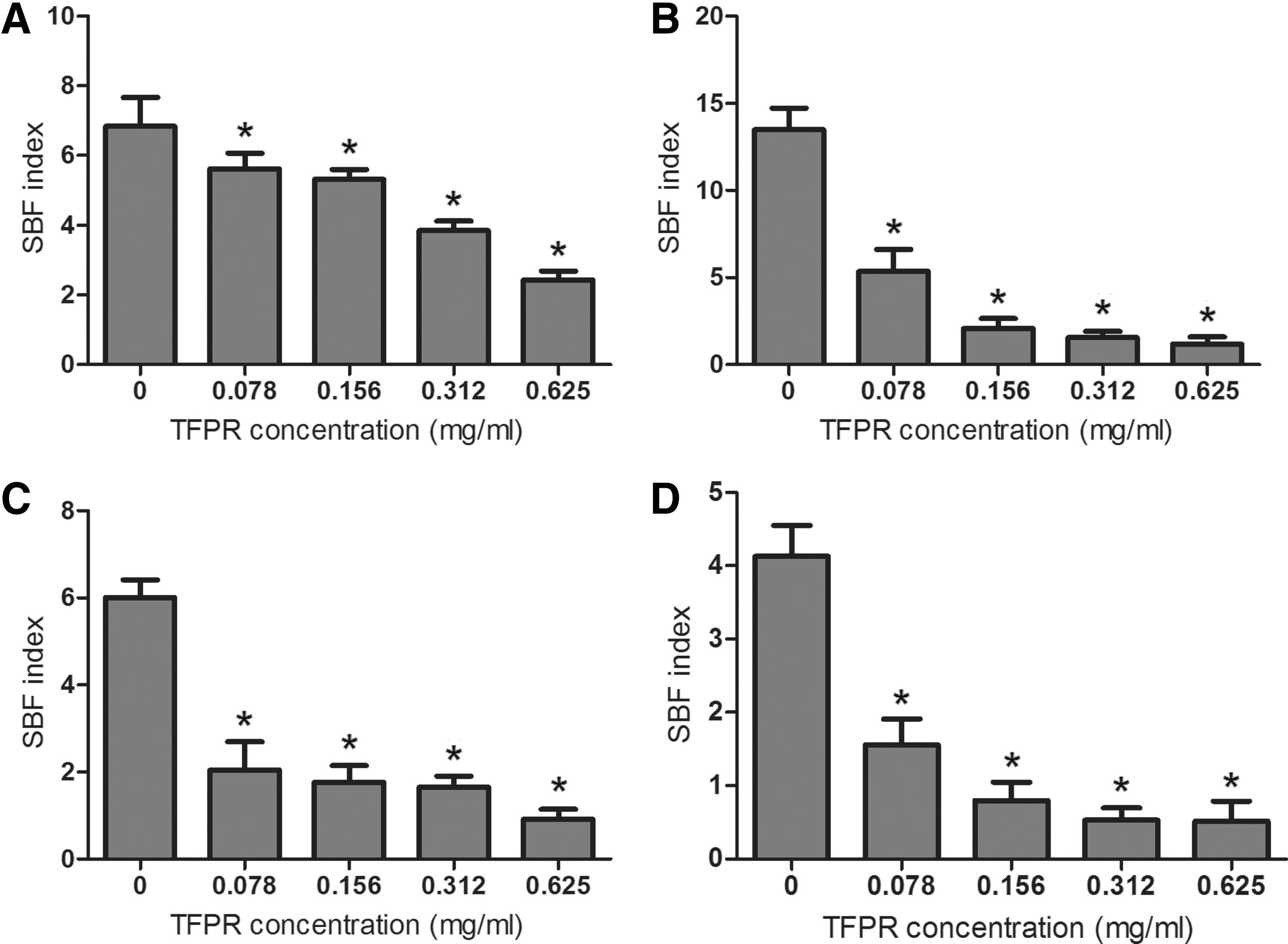
Inhibition of Escherichia coli biofilm formation by tannin-rich fraction from pomegranate rind (TFPR).
A digital inverted fluorescence microscope was utilized to visualize the effect of TFPR on biofilm. Inhibition of biofilm of E. coli ATCC 25922 that grew on the glass surface was confirmed by the decrease of blue fluorescence dots density (Fig. 3).
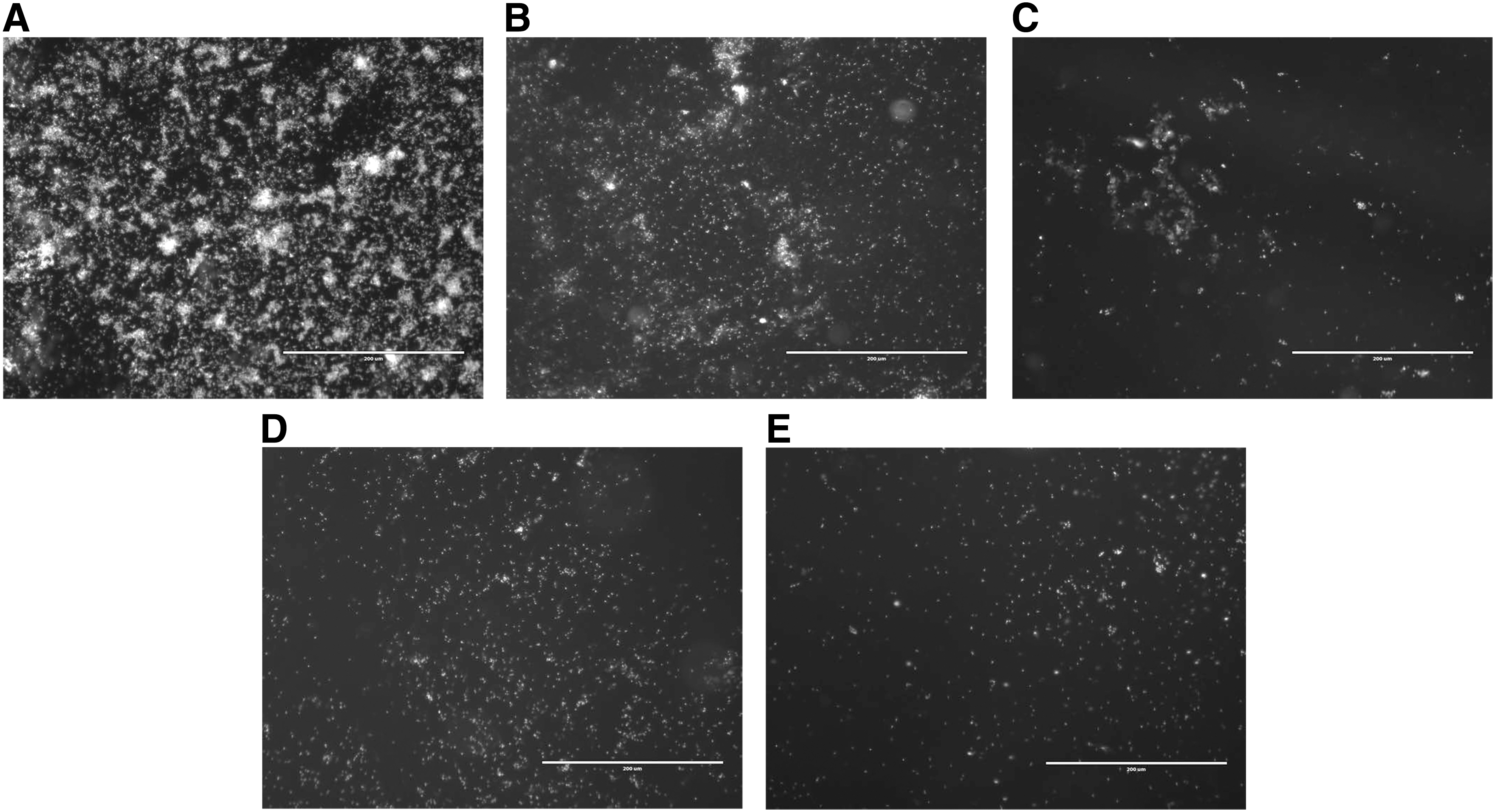
Biofilm formation of Escherichia coli ATCC 25922 stained by 4'-6-diamidino-2-phenylindole on glass surface upon the addition of tannin-rich fraction from pomegranate rind (TFPR).
Reduction of motility
As shown in Figure 4, TFPR reduced the motility of E. coli. For swimming motility, after treatment with TFPR at 0.156 and 0.312 mg/mL, the halo diameters were 73.43% and 21.31% of that of the control group (without TFPR), respectively. Similarly, swarming diameters of E. coli were 66.95% (at 0.156 mg/mL of TFPR) and 68.24% (at 0.312 mg/mL of TFPR) compared to the control group.
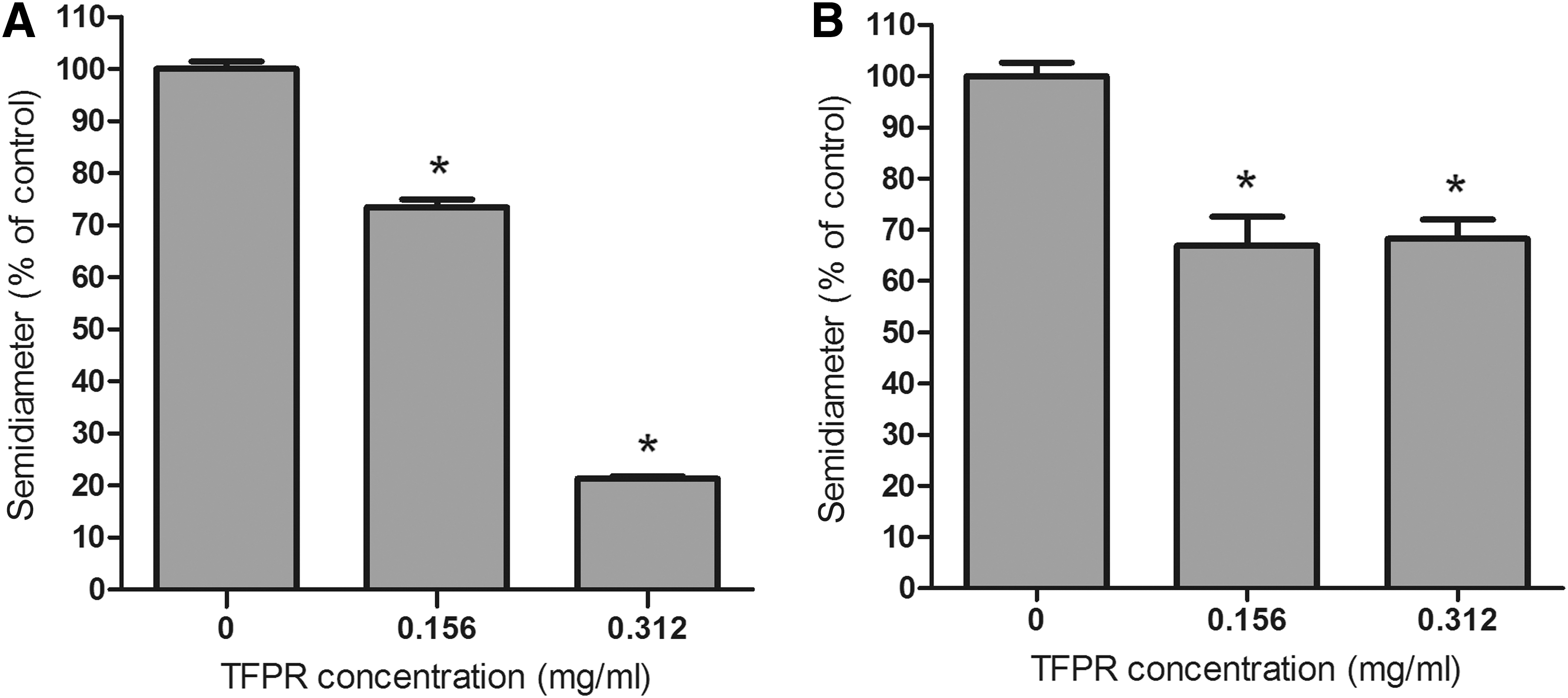
Tannin-rich fraction from pomegranate rind (TFPR) inhibits motility of Escherichia coli ATCC 25922.
Transcriptional changes of biofilm and motility-related genes in E. coli treated with TFPR
To investigate the genetic basis of biofilm and motility inhibition of E. coli, qRT-PCR was carried out to examine the differential expressions of related genes in E. coli with or without TFPR treatment. The gene expression patterns of several genes associated with biofilm formation and motility were downregulated by TFPR at the range of 0.039–0.156 mg/mL (Table 1). Curli genes (csgB and csgD) were repressed by TFPR (Table 1). TFPR also downregulated the expression of some motility-related genes (fimA, fimH, flhD, motB, qseB, and qseC) to various degrees.
Discussion
Biofilms are ubiquitous in nature and a frequent source of infections (Costerton et al., 1999). Bacteria cells in biofilms are more resistant to different disinfectants (Vestby et al., 2009). Therefore, biofilms have become increasingly problematic in food industries (Srey et al., 2013). In contrast with compounds aiming at killing or inhibiting the growth of microorganisms, antibiofilm compounds are capable of controlling biofilm formation without posing selective pressure on bacteria. These compounds may make bacteria unable to establish an infection (Rasmussen and Givskov, 2006; Cotar et al., 2013) or more susceptible to different agents (Taganna et al., 2011). Bacteria have been capable of displaying virulent behaviors, including biofilm formation, only until the number of their population reaches a definite threshold by using quorum-sensing systems (Kalia and Purohit, 2011). This allows bacteria to initiate an intensive and coordinated attack to host in a high population, which increases the probability that any defenses will be overwhelmed (Raina et al., 2009; Galloway et al., 2012). A QS system could represent an ideal target of controlling a wide range of bacterial infections, and compounds showing QS inhibition capacity could have a promising application prospect (Defoirdt et al., 2013; Starkey et al., 2014).
Production of violacein by Chromobacterium violaceum is regulated by the AI-1 QS system (McClean et al., 1997). Therefore, a wild-type strain C. violaceum ATCC 12472 was used to evaluate the QS inhibitory capacity of TFPR as previously described (Taganna et al., 2011; Zhu et al., 2011; Truchado et al., 2012). In the present study, TFPR demonstrated an inhibitory effect on production of violacein of C. violaceum in a concentration-dependent manner without interfering with its growth (Fig. 1). Therefore, we hypothesized that the inhibitory effect on violacein production is caused by disruption of AI-1 QS signaling systems rather than by inhibition of bacterial cell growth, which will generate selective pressure for the development of resistant bacteria (Kolar et al., 2001). Similar findings have been reported for punicalagin (the main component of TFPR) and urolithins (punicalagin metabolites produced by colon microbiota), which are shown to exert anti-QS effects on Salmonella and Yersinia enterocolitica, respectively (Gimenez-Bastida et al., 2012; Li et al., 2014b), and tannin-rich fraction from Terminalia catappa was also found to inhibit QS (Taganna et al., 2011).
QS has been considered to play a significant role in formation of biofilms (Miller and Bassler, 2001; Hall-Stoodley et al., 2004; Bai and Rai, 2011). Ravichandiran et al. have demonstrated that biofilm formation in E. coli could be reduced by inhibiting SdiA (Ravichandiran et al., 2013). Since TFPR showed inhibitory activity against the AI-1 QS system, it is assumed that TFPR may also interfere with the biofilm formation in E. coli. The observation from crystal violet dye assay (Fig. 2) and microscopy assay (Fig. 3) supported this hypothesis. E. coli biofilm formation was significantly inhibited by TFPR at a concentration range of 0.156–0.625 mg/mL in a concentration-dependent manner, which was in accordance with fluorescence microscope observation. Based on these findings, we hypothesized that TFPR inhibits E. coli biofilm formation possibly by interfering with AHL-based QS dysfunction. Similarly, the tannin-rich fraction from T. catappa and urolithins reduced biofilm formation of different bacteria (Taganna et al., 2011; Gimenez-Bastida et al., 2012).
Curli (adhesive fimbrial structures) could contribute to E. coli biofilm formation by binding cells together, and is thought to be a virulence characteristic (Ryu and Beuchat, 2005; Uhlich et al., 2006; Rendon et al., 2007). In the present study, TFPR was found to inhibit curli genes expression (csgD and csgB). Several other phytochemicals have been found to be capable of downregulating curli genes and reducing bacteria biofilm formation (Ren et al., 2005; Hancock et al., 2010; Vikram et al., 2010; Truchado et al., 2012; Lee et al., 2014b). Nevertheless, bacteria develop biofilm through a complex process, involving different genes and environmental factors (Hall-Stoodley et al., 2004; Parsek and Greenberg, 2005), and the exact mechanism by which TFPR inhibits E. coli biofilm formation need to be further investigated.
Bacterial motility has also been one of the phenotypes regulated by QS system (Miller and Bassler, 2001). A close link lies between bacterial biofilm and motility (Beloin et al., 2008; Wood, 2009). In soft agar, individual bacterial cells swim in the medium, while migrating cooperatively over the surface, which is defined as swarming (DiLuzio et al., 2005). Swarming is a QS-related multicellular behavior, while swimming seems to be at the individual cell level (Kohler et al., 2000; Vestby et al., 2014). Both swarming and swimming motility were significantly affected by TFPR, which indicates that TFPR could affect motility directly in addition to inhibition of AHL-based QS, which agreed with transcriptional changes in several key motility genes, such as type 1 fimbriae genes (fimA and fimH), flagellar transcriptional activator gene (flhD), flagellar motor protein (MotB), and qseB and qseC genes, which activate flhDC transcription (Sperandio et al., 2002). This is consistent with other studies in which motility of human opportunistic pathogen Pseudomonas aeruginosa is inhibited by ellagitannin-rich extracts, such as berry extracts and pomegranate extracts (Vattem et al., 2007; O'may et al., 2012).
In conclusion, TFPR was capable of interfering with AHL-based QS in C. violaceum. Moreover, TFPR inhibited biofilm formation and motility of Escherichia coli, possibly partly through AHL-based QS inhibition. TFPR also caused transcriptional changes in some biofilm and motility-related genes in E. coli. These findings suggest that TFPR has the potential to be applied to control persistent E. coli contamination and biofilms in the food industry.
Footnotes
Acknowledgments
This work was supported in part by New Century Excellent Talent Support Plan (NCET-13-0488), the Twelve-five Science and Technology Support Program (No. 2015BAD16B08), National Natural Science Foundation of China (31301498), Fundamental Research Funds in Northwest A&F University (Z109021424), International Collaboration Partner Plan (A213021203) in Northwest A&F University, Special Fund for Sino–US Joint Research Center for Food Safety (A200021501), and Start-up Funds for Talents in Northwest A&F University (Z111021403).
Disclosure Statement
No competing financial interests exist.