
Abstract
Many issues have been found to be related to food preservation and food contamination caused by various pathogenic bacteria in recent years. Many antibacterial agents act efficiently against Gram-positive foodborne bacteria; however, they are less effective against Gram-negative foodborne bacteria. In the present study, an attempt has been made to evaluate the antibacterial activity of pyrolysis oil manufactured from Pinus densiflora (PLO) against two Gram-negative foodborne pathogenic bacteria, Salmonella Typhimurium and Escherichia coli O157:H7. PLO possessed potent antibacterial activity against both foodborne pathogenic bacteria, as indicated by inhibition zones of 10.33–12.33 mm and minimum inhibitory concentration and minimum bactericidal concentration values of 250–500 μg/mL and 500–1000 μg/mL, respectively. PLO at the minimum inhibitory concentration exhibited an inhibitory effect on the viability of the bacterial pathogens with leakage of 260 nm absorbing materials, an increase in the relative electrical conductivity, and loss of salt tolerance capacity. PLO exhibited promising antibacterial activity against both of the Gram-negative foodborne pathogenic bacteria and thus it can be utilized in the food sector and pharmaceutical industries for the development of antibiotics and preservatives.
Introduction
D
Other foodborne pathogens are considered to be emerging because of the role of food in their usual transmission (Armstrong et al., 1996). Infection with Escherichia coli O157:H7 has emerged as a major cause of bloody diarrhea and acute renal failure (Simopoulos, 2004; Scallan et al., 2011). Outbreaks of bacterial infection, generally associated with consumption of beef, have been reported in Australia, Canada, Japan, the United States, European countries, and southern African countries (Marija et al., 2013).
Control of food spoilage and pathogenic microorganisms is mainly achieved by chemical treatment; however, the use of synthetic chemicals is limited due to a number of undesirable aspects with the consequent development of environmental pollution problems (Devlieghere et al., 2004; Barbosa et al., 2009; Faleiro, 2011). Awareness among consumers has led to a search for natural antibacterial agents (Faleiro, 2011). Many natural compounds and oils of plant origin have been successfully tested against foodborne pathogens (Deans and Ritchie, 1987; Miksusanti et al., 2008; Cattelan et al., 2013; Diao et al., 2013, 2014; Dussault et al., 2014). However, most natural compounds tested are primarily effective against Gram-positive pathogenic bacteria, while they are ineffective or less effective against Gram-negative pathogenic bacteria (Cattelan et al., 2013; Diao et al., 2013, 2014; Dussault et al., 2014).
Japanese red pine (Pinus densiflora Siebold and Zucc.) is an evergreen tree found throughout East Asian countries including Korea and Japan (Kim et al., 2011). Different parts of this plant have been utilized in traditional medicines to cure various diseases (Satoh et al., 1996). Pyrolysis oils (PLO) are the liquid products obtained by the burning of biomass materials such as agricultural and forestry byproducts, algal biomass, and plant materials at 500°C or higher (Lemoine et al., 2013). These PLOs contain a number of natural compounds that can be utilized in the formulation of drugs against Gram-negative foodborne pathogenic bacteria.
In the present study, the antibacterial potential of pyrolysis oil made from PLO was evaluated along with the mode of its antibacterial action against two Gram-negative foodborne pathogenic bacteria (Salmonella Typhimurium and E. coli O157:H7).
Materials and Methods
Production of PLO and chemicals
PLO was produced from bark-free wood chips of Japanese red pine (P. densiflora Siebold and Zucc.) obtained from the Korea Forest Research Institute using the standard pyrolysis process (Patra et al., 2015). The sawdust of P. densiflora was pyrolized in a lab-scale fluid-bed fast pyrolysis procedure at 500°C in a nitrogen atmosphere with a pyrolysis product residence time of 1.3 s. The manufactured PLO was kept at 4°C until further analysis.
Standard kanamycin and other chemicals were purchased from Sigma (Sigma-Aldrich Co., St. Louis, MO). Nutrient broth (NB) media was obtained from Difco (Becton, Dickinson and Company, Sparks Glencoe, MD).
Chemical analysis of PLO
Detailed analysis of the chemical composition of volatile compounds present in PLO was conducted using a gas chromatography–mass spectrometry system (JMS 700 Mstation; Jeol Ltd., Tokyo, Japan) equipped with an Agilent 6890N GC DB-5 MS fused silica capillary column (Agilent Technologies, Santa Clara, CA) according to (Patra et al., 2015). Samples (1 μL of 100 times-diluted samples in methanol) were injected manually in splitless mode through the injector. Identification of volatile compounds are done by the computer matching of mass spectra using the Wiley and National Institute of Standards and Technology libraries for the gas chromatography–mass spectrometry (GC-MS) system.
The chemical structures of some selected antimicrobial compounds were drawn using the ACD Chemsketch software (
Evaluation of antibacterial activity of PLO
Microbial strains
Foodborne pathogenic bacteria were obtained from American Type Culture Collection (ATCC, Manassas, VA) and maintained in NB media at −80°C until further use. Two strains each of two Gram-negative foodborne pathogenic bacteria, Salmonella Typhimurium (ATCC 19586 and 43174) and E. coli O157:H7 (ATCC 35150 and 43890), were used in the present study to evaluate the antibacterial efficacy of PLO.
Determination of antibacterial activity of PLO
The antibacterial potential of PLO was evaluated by disc diffusion assay (Bajpai et al., 2013). Sterile filter paper discs 8 mm in diameter (Advantec; Toyo Roshi Kaisha Ltd., Tokyo, Japan) impregnated with 1000 μg/disc of PLO diluted in 5% dimethyl sulfoxide (DMSO) was used. Kanamycin at 40 μg/disc was used as a positive control, while 5% DMSO served as a negative control. Each assay was carried out in triplicate, and the diameter of zone of inhibition was measured in millimeters after incubation as 37°C for 24 h.
The minimum inhibitory concentration (MIC) of PLO was tested by the twofold serial dilution method (Bajpai et al., 2013). Different concentrations of PLO (2000–125 μg/mL) were added to NB media containing the test pathogen and were incubated at 37°C overnight. The lowest concentration of PLO that did not show any visible growth of test organisms was determined as MIC, while the concentration that did not show any growth of bacterial colonies when grown on NA plates was defined as minimum bactericidal concentration (MBC).
Antibacterial mode of action of PLO
Effects of PLO on cell viability
The effects of PLO on cell viability were evaluated according to standard procedure (Bajpai et al., 2013). Active cultures, Salmonella Typhimurium ATCC 43174 (ST) and E. coli O157:H7 ATCC 43890 (EC) were used in the study. For each bacterium, 1 mL of stock solution (approximately 107colony-forming units [cfu]/mL) was transferred to 2-mL vials, centrifuged at 9,950 × g for 10 min and the pellets were then resuspended in 1 mL of phosphate-buffered saline (PBS). Each tube containing 900 μL of resuspended bacterial suspension were inoculated with 100 μL PLO at MIC and incubated at 37°C. For viable cell counts, 100 μL of sample were collected at 2-h intervals for 8 h, diluted appropriately with PBS, spread on nutrient agar (NA) plates, and incubated at 37°C for 24 h, after which the colonies were counted.
Release of 260-nm absorbing cellular materials
The effects of PLO on the release of 260 nm absorbing cellular materials from ST and EC were determined (Carson et al., 2002). Briefly, 2 mL of bacterial culture in peptone water (0.1 g/100 mL) were added to PLO at MIC for treatment and 5% DMSO for the control and incubated at 37°C. Samples were collected at each time interval (0, 60, and 120 min) and centrifuged at 1,210 × g, after which the optical density (OD) of the supernatant was measured at 260 nm using a spectrophotometer (ASP-3700; ACT Gene Inc., Piscataway, NJ). The results were expressed as the OD at 260 nm with respect to time.
Effects of PLO on cell membrane permeability
The cell membrane permeability of bacteria was determined using standard method (Kong et al., 2008). ST and EC culture were incubated at 37°C for 10 h, then centrifuged at 2,480 × g for 10 min. The bacteria were subsequently washed with 5% glucose to reduce their electrical conductivity to levels close to that of 5% glucose. PLO at MIC was added to 5% glucose solution and its electrical conductivity was measured and denoted as L1. Similarly, PLO was added to the isotonic bacterial solution, completely mixed, and incubated at 37°C, during which its conductivity was measured every 2-h time interval for 8 h and denoted L2. Bacterial solution in 5% glucose was boiled for 5 min and then used as the control (L0). All measurements were taken using a conductivity meter (Con 6; LaMotte Company, Chestertown, MD) and the relative electrical conductivity was calculated as follows
Effects of PLO on leakage of potassium ions
Leakage of potassium (K+) ion from the bacterial cells was determined by Lee et al. (2002). Briefly, ST and EC suspended in peptone water (0.1 g/100 mL) were treated with the MIC of PLO (treatment) and 5% DMSO (control), then incubated at 37°C. The leakage of free K+ ions in the bacterial suspensions was measured after every 2 h for 8 h by a photometric procedure using a kalium/potassium kit (Quantofix, GmbH, Wiesbaden, Germany). The results were expressed as the amount of extracellular free K+ (mg/L) released in the growth media at each time interval.
Effects of PLO on salt tolerance
The loss of salt tolerance capacity of ST and EC treated with PLO at MIC was investigated by Miksusanti et al. (2008). PLO (treatment) and 5% DMSO (control) were added to overnight-grown cultures of bacteria, after which the samples were incubated at 37°C for 60 min. The samples were then serially diluted and spread on NA plates supplemented with NaCl (0, 2.5, 5, and 10%). The number of cfu on each NA-NaCl plate after 24 h of incubation at 37°C was compared to that on the control NA plate and expressed as log cfu/mL.
Scanning electron microscopy (SEM) analysis
The effects of PLO on bacterial cell morphology were analyzed by SEM (Bajpai et al., 2013). Briefly, overnight-grown culture of ST and EC treated with PLO at MIC (treatment) and 5% DMSO (control) were centrifuged at 3000 rpm for 5 min, then washed twice with 50 mM phosphate buffer (pH 7.2). The culture was subsequently spread on glass slides and fixed with 2.5% glutaraldehyde, after which samples were dehydrated with different concentrations of ethanol (50–100%) followed by t-butanol. Finally, the glass slide was sputter coated with platinum and observed by SEM (S-4100, Hitachi, Tokyo, Japan).
Statistical analysis
All numeric data represent the means ± standard deviation. The variance of the sample data was determined by Duncan's test using Statistical Analysis Software (SAS) version 9.4 (SAS Inc., Cary, NC).
Results
Chemical analysis of volatile compounds of PLO
Different classes of volatile compounds present in PLO analyzed by GC-MS are presented in Table 1. All identified volatile compounds were categorized into seven different classes, namely, phenols (16.68%), furans (14.03%), and benzenes (11.51%) found in higher quantity, followed by alkanes (7.03%), acids (4.16%), ketones (2.47%), and others (42.28%) (Table 1). The GC-MS spectra and chemical structure of some selected antibacterial compounds present in PLO are shown in Figure 1.
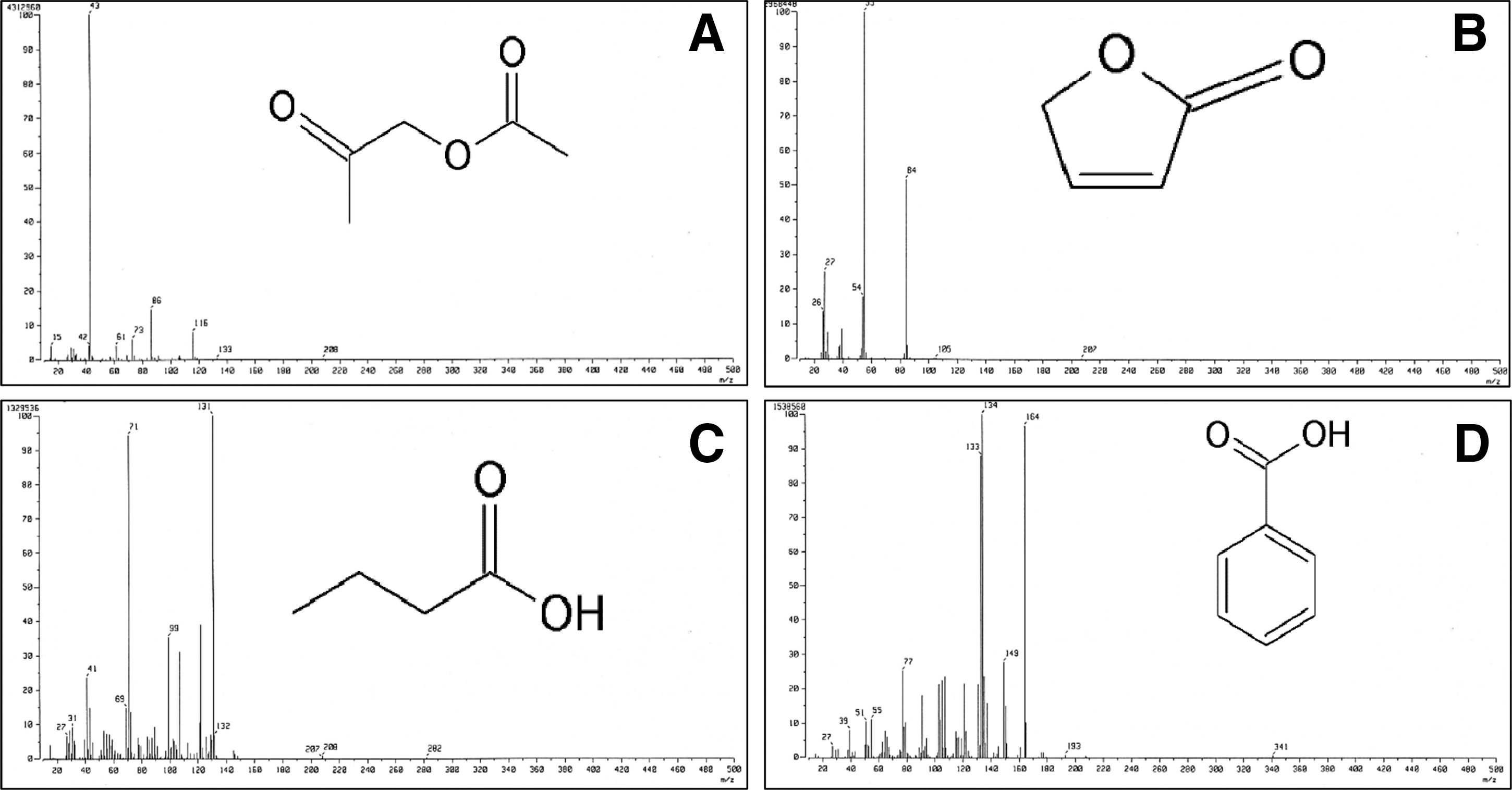
Chemical structure and spectra of some compounds with antimicrobial activity present in pyrolysis oil of Pinus densiflora Siebold and Zucc.
Antibacterial activity of PLO
PLO at 1000 μg/mL inhibited all strains of Salmonella Typhimurium and E. coli O157:H7 with 10.33–12.33 mm inhibition zone (Table 2). PLO displayed significantly higher inhibition activity toward Salmonella Typhimurium than E. coli O157:H7 at p < 0.05. Kanamycin at 40 μg/mL had higher antibacterial activity than PLO, but 5% DMSO did not have any activity. The MIC and MBC values against Salmonella Typhimurium and E. coli O157:H7 ranged from 250 to 1000 μg/mL (Table 3).
Data are expressed as the means ± standard deviation (n = 3).
Values in the same column with different superscripts are significantly different (p < 0.05).
Effects of PLO on cell viability
Based on the sensitivity of the four strains of the tested foodborne bacteria, one strain from each species (ST and EC) with higher antibacterial activity (Table 2) were selected for further investigation to elucidate the possible mode of antibacterial action of PLO against Gram-negative bacteria. The effects of PLO on cell viability are presented in Figure 2. PLO at MIC reduced the viability of the bacteria with respect to time. PLO had significantly higher activity against ST, showing complete inhibition on NA plates after 4 h of incubation (Fig. 2A). PLO inhibited the viability of EC after 8 h of incubation (Fig. 2B). No change in the number of cfu was observed on the control plates for both bacteria.
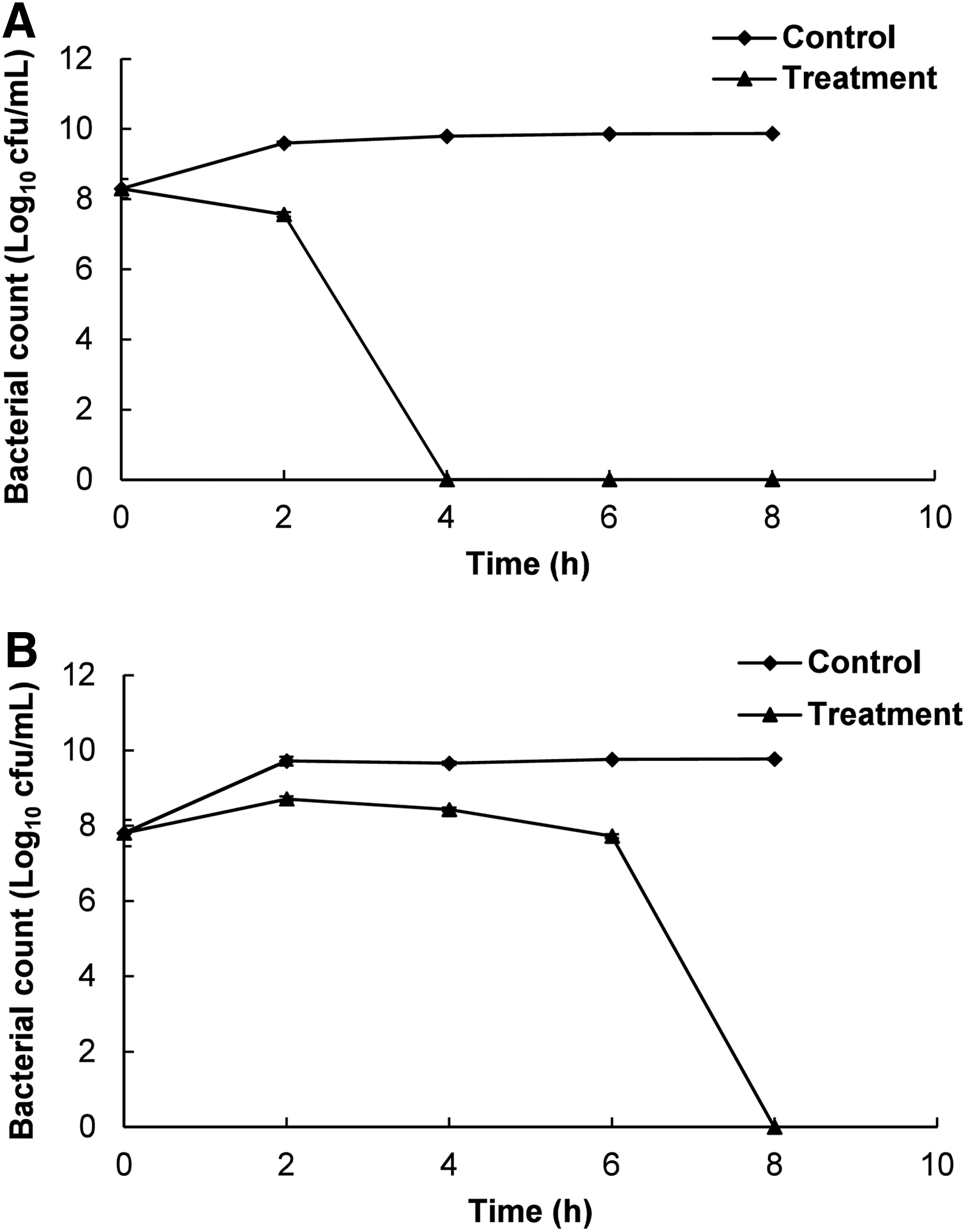
Effects of pyrolysis oil of Pinus densiflora Siebold and Zucc. (PLO) at the minimum inhibitory concentration on viability of the tested foodborne pathogenic bacteria.
Effects of PLO on the release of 260-nm absorbing materials
There was a continuous release of cellular material, as evident from the increase in OD values in both PLO-treated ST and EC with increasing incubation time (Fig. 3). However, there was no distinct change in the untreated control. Cellular leakage was more severe for ST than EC when both were exposed to PLO for 60 min (Fig. 3).
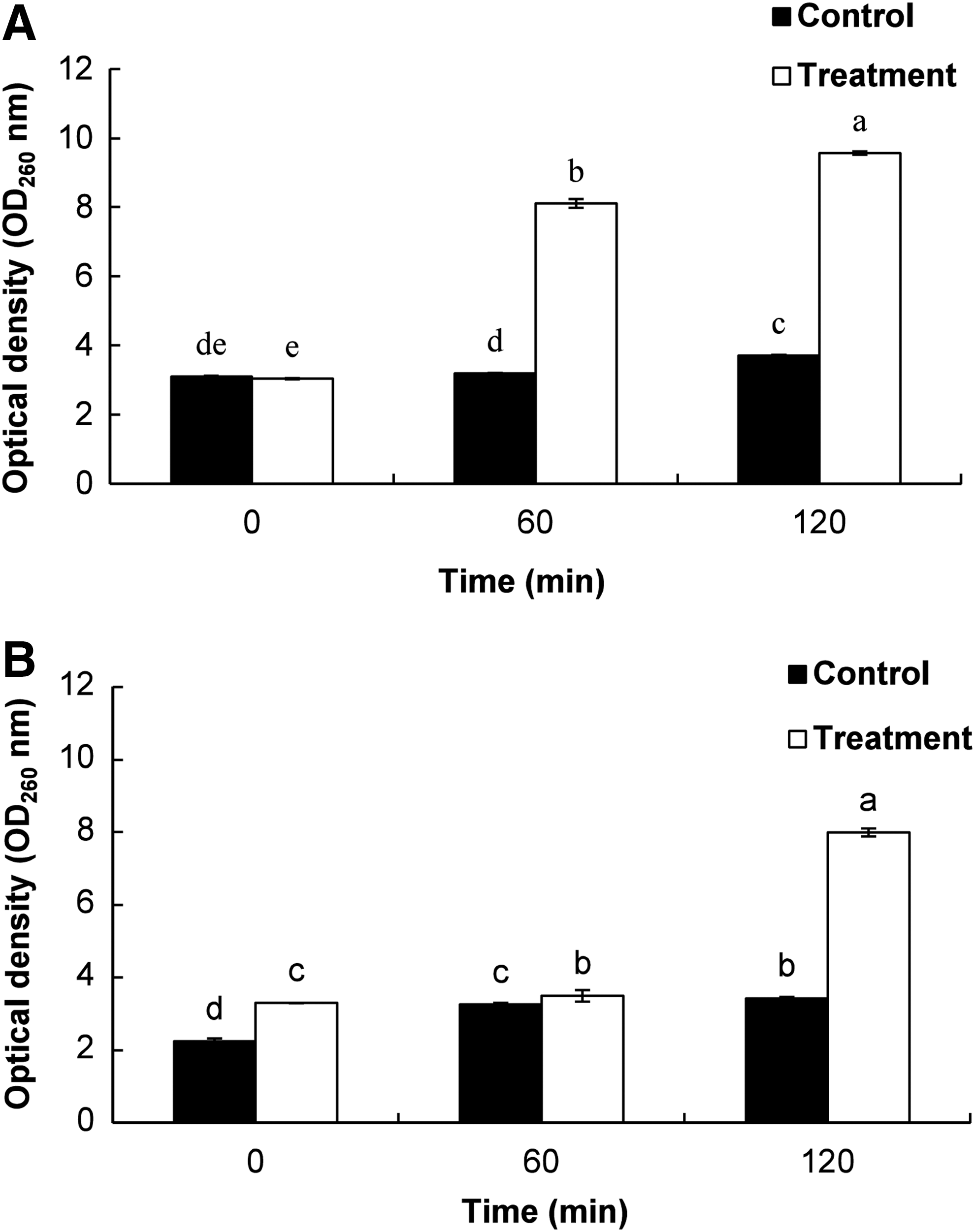
Effects of pyrolysis oil (PLO) of Pinus densiflora Siebold and Zucc. at the minimum inhibitory concentration on the release rate of 260-nm absorbing material on the tested foodborne pathogenic bacteria.
Effects of PLO on cell membrane permeability and K+ ion leakage
PLO-treated bacteria showed a continuous increase in the relative electrical conductivity with respect to time (Fig. 4). DMSO-treated controls showed no increase in the relative electrical conductivity. The bacterial cells treated with MIC of PLO showed a continuous release of intracellular K+ ions with respect to time, whereas DMSO-treated control bacteria showed no K+ ion leakage (Fig. 5).
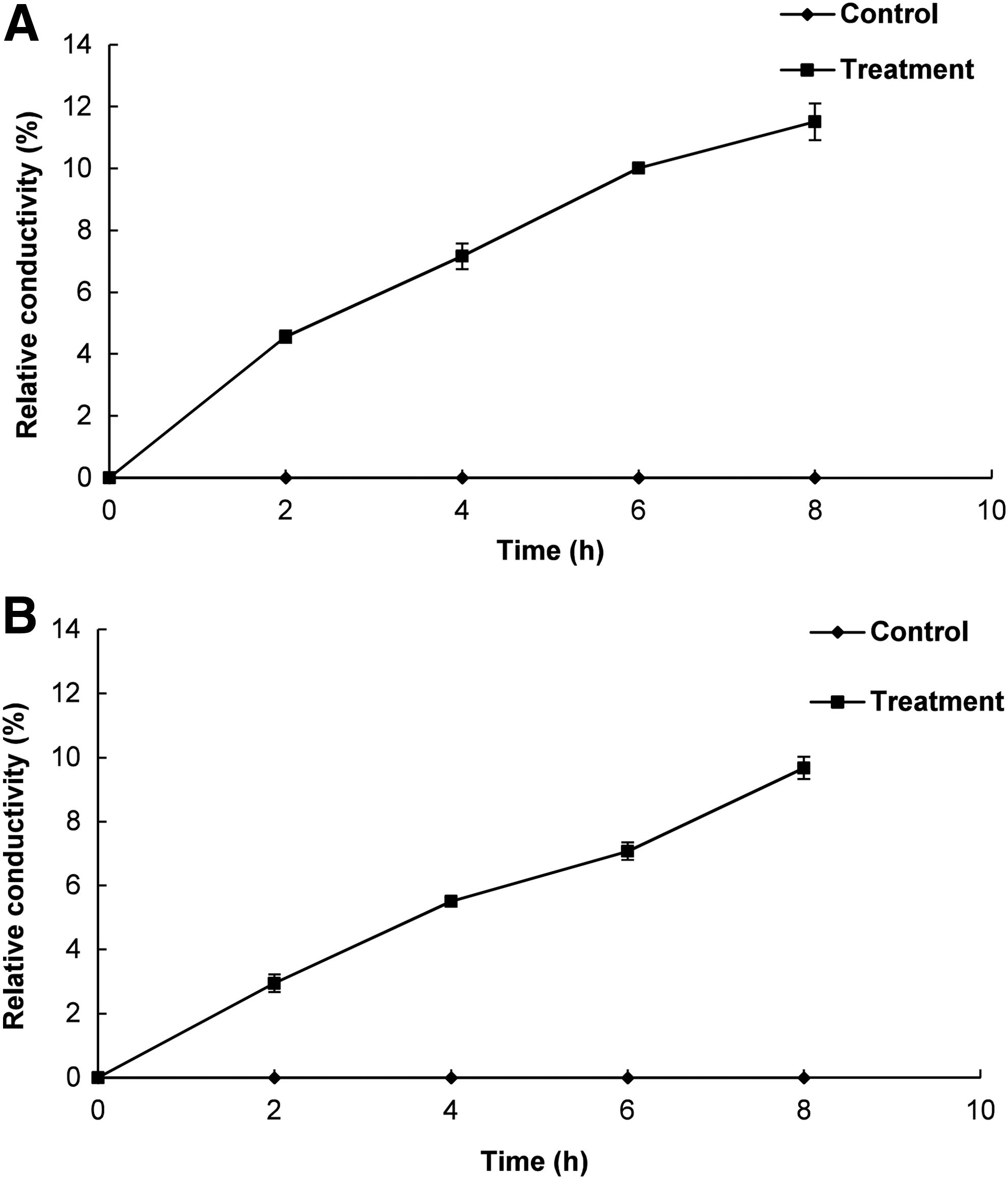
Effects of pyrolysis oil (PLO) of Pinus densiflora Siebold and Zucc. at the minimum inhibitory concentration on the cell membrane permeability of the tested foodborne pathogenic bacteria.
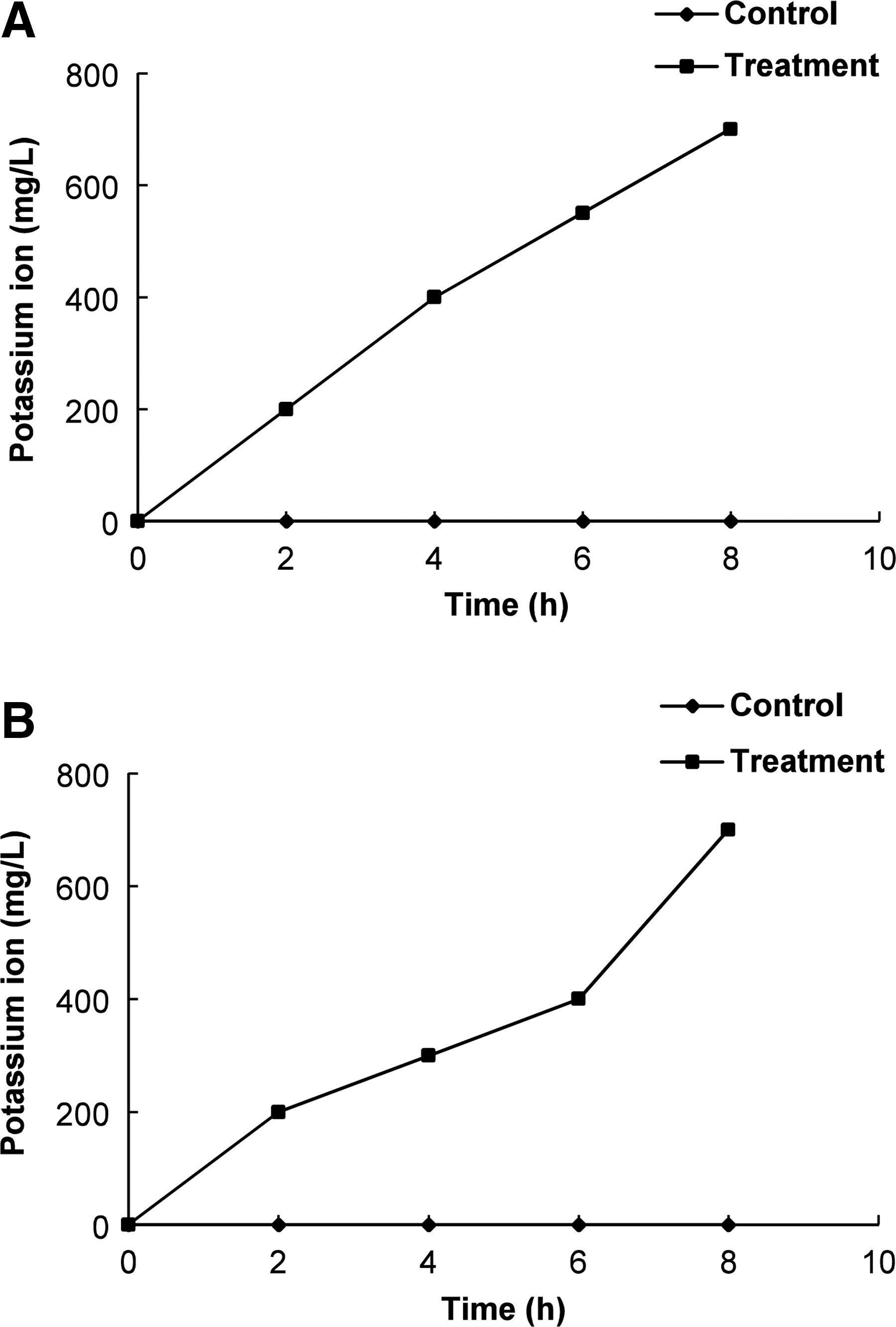
Effects of pyrolysis oil (PLO) of Pinus densiflora Siebold and Zucc. at the minimum inhibitory concentration on the leakage of potassium ions from the tested foodborne pathogenic bacteria.
Effects of PLO on loss of salt tolerance capacity
PLO-treated bacteria showed reduced levels in response to different concentrations of NaCl (0, 2.5, and 5%) relative to DMSO-treated control bacteria (Fig. 6). The treated and control bacteria showed no potential growth in the media supplemented with 10% NaCl.
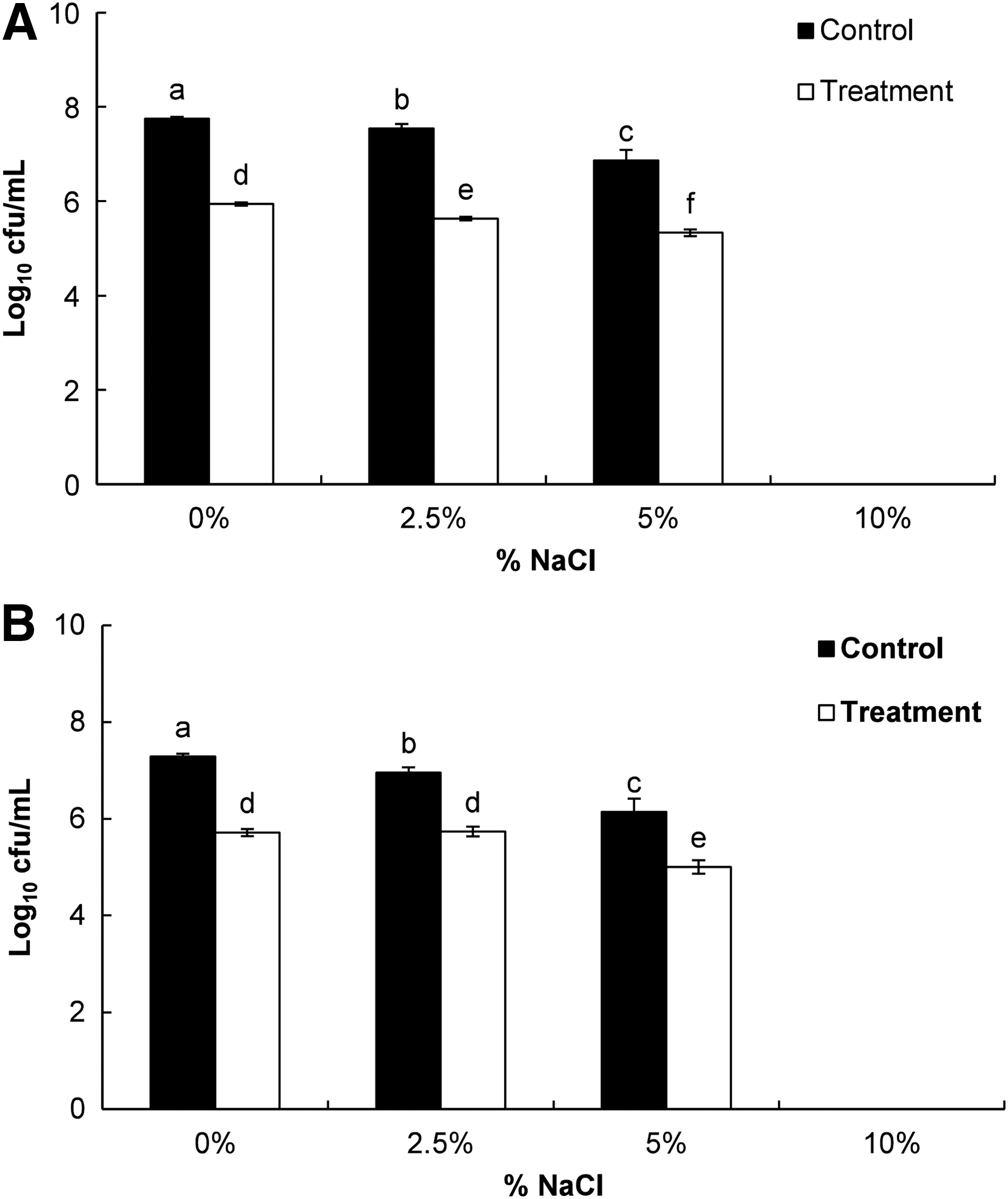
Effects of pyrolysis oil (PLO) of Pinus densiflora Siebold and Zucc. at the minimum inhibitory concentration on loss of salt tolerance of the tested foodborne pathogenic bacteria.
SEM analysis
The SEM images showed that PLO caused a detrimental effect on the morphology of the bacterial cells, resulting in complete lysis of bacteria (Fig. 7B and D), whereas the control bacteria treated with DMSO showed distinct and regular cell surfaces (Fig. 7A and C).
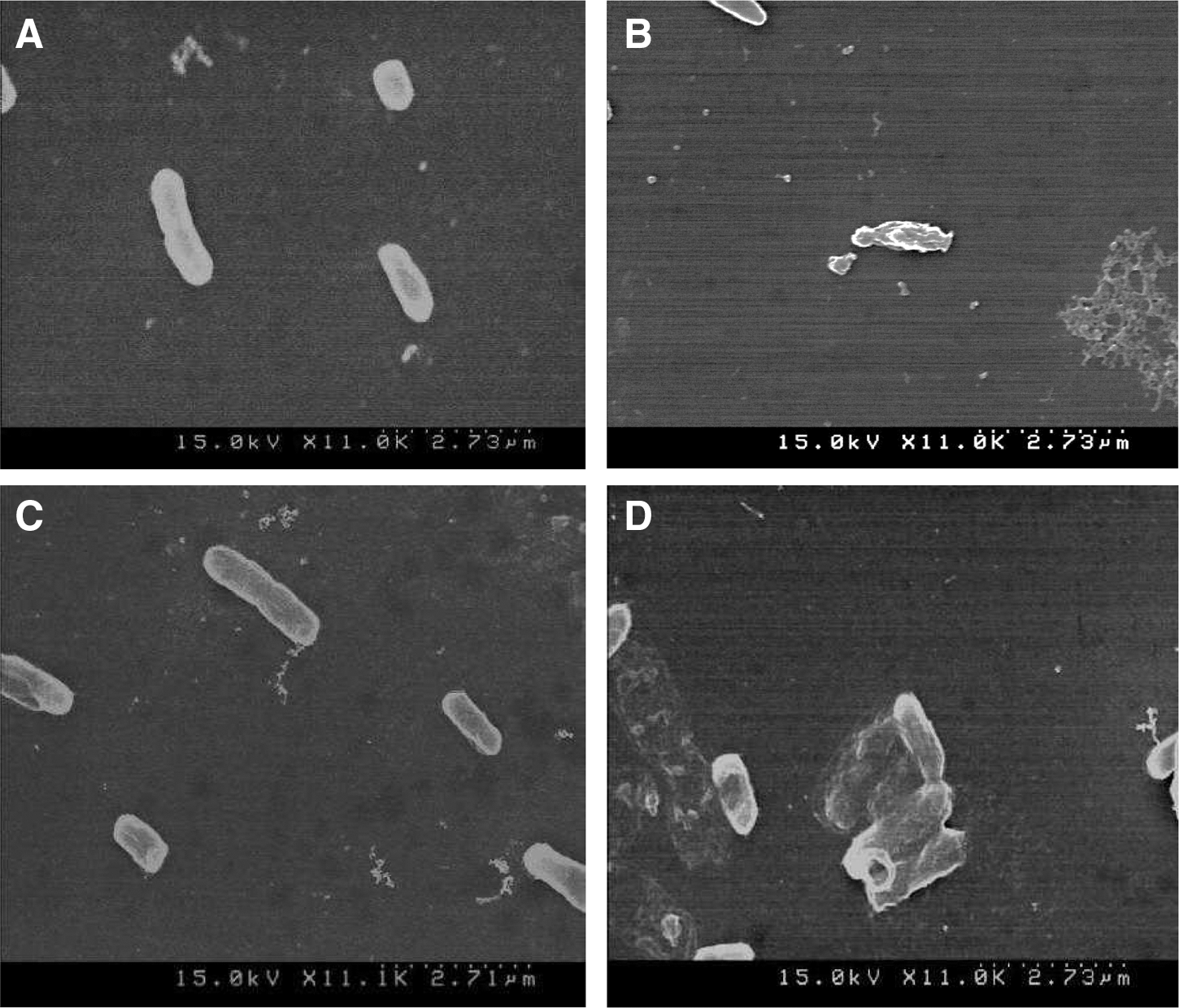
Scanning electron microscopy analysis of Escherichia coli O157:H7 ATCC 43890
Discussion
Foodborne pathogenic bacteria affect the nutritional quality of the food to a great extent, which leads to various harmful diseases (Diao et al., 2014). In the present study, an attempt was made to test the bactericidal effect of PLO and depict a possible hypothesis for its antibacterial mode of action against Gram-negative bacteria.
The chemical composition of volatile compounds in PLO analyzed by GC-MS analysis was described in detail in our earlier publication (Patra et al., 2015). Several compounds present in PLO, including 2-propanone, 2(5H)-furanone, butanoic acid, and benzoic acids (Fig. 1), were previously reported to possess antimicrobial potential (Kawase et al., 2001; Namkung et al., 2011; Pundir and Jain, 2011; Babynin et al., 2012). PLO exerted potential inhibitory effects on both foodborne pathogenic bacteria with very low MIC and MBC values (Tables 2 and 3). Similar results were previously reported for PO effects on other pathogenic bacteria (Bedmutha et al., 2011; Phukan et al., 2013). The antibacterial potential of PLO might be due to presence of furan, ketone, and acid classes of compounds, particularly the antimicrobial compounds present in PLO (Table 1).
The mechanism of action of PLO against Gram-negative bacteria was predicted based on its effects on the viability of bacterial cells, membrane permeability, and salt tolerance capacity. When bacterial cells were exposed to the MIC of PLO, a significant decrease in the number of cfu was observed relative to the untreated control bacteria (Fig. 2). Gram-negative bacteria possess a hydrophilic outer membrane due to the presence of lipopolysaccharide molecules; however, these lipopolysaccharide barriers might not be completely impermeable to hydrophobic PLO molecules, and some might have traversed slowly inside the bacterial cells, causing cell lysis (Kotzekidou et al., 2008). A similar hypothesis was reported previously (Nikaido, 1996; Helander et al., 1998; Kotzekidou et al., 2008).
When PLO-treated ST and EC were tested at 260 nm, a significant increase in the OD values with time was observed (Fig. 3). These data indicated significant loss of intracellular materials such as metabolites, ions, and nucleic acids, into the bacterial suspensions that were absorbed at 260 nm (Cox et al., 2003). The leakage may have occurred due to lysis following accumulation of excessive amounts of PLO inside the cell (Denyer, 1990). Similarly, a continuous increase in the relative electrical conductivity and loss of K+ ions in the PLO-treated bacterial sample was observed relative to the untreated control (Figs. 4 and 5), which might also have been due to the normal lysis, disruption, and death of bacteria caused by the accumulation of PLO in the intracellular region of the bacterial cell (Cox et al., 1998; Bajpai et al., 2013).
Following exposure of the bacterial cells to PLO and subsequent growth on NA plates supplemented with different concentrations of NaCl (2.5 and 5%), significant decreases in the number of cfu were observed relative to the DMSO-treated control bacteria (Fig. 6). This might have occurred because PLO influenced the ability of the membrane to osmoregulate cells to resist the increased salt concentration, which is in accordance with the results of previous studies (Carson et al., 2002; Miksusanti et al., 2008). Furthermore, SEM analysis revealed distinct morphological alterations on the treated bacterial cell surfaces (Fig. 7), further supporting our hypothesis.
Conclusions
The results of this study revealed that PLO possesses significant in vitro antibacterial activity against two Gram-negative foodborne bacteria, Salmonella Typhimurium and E. coli O157:H7. Although the exact mode of action of PLO against the bacteria is still not clear, the results of cell viability, release assays, and effects on salt tolerance capacity revealed that the loss of integrity of the cell membrane and loss of vital intracellular constituents could be possible mechanisms of action of PLO against these foodborne pathogenic bacteria. Overall, PLO can be effectively utilized in formulations of antibacterial drugs for treatment of Gram-negative bacteria during food preservation and processing.
Footnotes
Acknowledgment
This research was conducted with support from a research fund provided by Yeungnam University in 2014.
Disclosure Statement
No competing financial interests exist.