
Abstract
NmlR, a negative transcription regulator in the MerR family, is involved in oxidative and nitrosative stress response in Neisseria gonorrhoeae and Haemophilus influenzae. In this study, the objective was to characterize the role and the regulon of NmlR in the foodborne Listeria monocytogenes. An L. monocytogenes nmlR null mutant strain was constructed. Transcriptomes of strain 10403S wild type (WT) and ΔnmlRlm strains grown to the stationary phase were determined by mRNA sequencing. Differential expression analyses revealed 74 genes with altered expression levels (>9-fold difference), comprising 46 negatively and 28 positively regulated genes. Twenty-four NmlRlm-dependent genes overlap with the members of previously identified regulons of HrcA, a negative regulator of heat response in L. monocytogenes, and of alternative sigma factor σH. Phenotypic characterization revealed that the ΔnmlRlm strain survived significantly less than the WT under acid stress (pH 2.5 for 1 h) and oxidative stress (3% hydrogen peroxide for 1 h). In addition, nmlRlm deletion also resulted in a significant decrease (p < 0.0005) of cell length and enhanced intracellular growth in a differentiated macrophage-like U937 cell line during entry into stationary phase. These findings indicate that NmlRlm is not only involved in oxidative stress response but also contributes to other characteristics such as acid tolerance and intracellular growth, either through direct regulation or co-regulation with other regulators such as HrcA and σH.
Introduction
L
After ingestion by a host, L. monocytogenes has to survive acidic and alkaline conditions in the host's gastrointestinal tract. Within a phagosome, L. monocytogenes encounters acidic conditions, reactive oxygen species, and reactive nitrogen species before it can escape to multiply in the cytosol of a host cell (Vazquez-Boland et al., 2001). Reactive oxygen species are found not only inside the macrophage but are also produced as a byproduct of aerobic processes (Imlay, 2002). The oxidants can react with cellular components to produce reactive electrophiles (Nguyen et al., 2009). Resistance mechanisms to reactive oxygen, nitrogen, and electrophile species are important for L. monocytogenes' survival and persistence in the host cell. The regulator PerR has been found to participate in the oxidative stress response in Bacillus subtilis and Staphylococcus aureus (Rea et al., 2005). In the perR mutant, oxidative stress response genes such as kat (catalase), trxB (thioredoxin reductase), lmo0641 (homolog of ZosA in Bacillus), fri (iron-binding ferritin), and hemA (homolog of hemX and involved in heme biosynthesis in Bacillus) were upregulated compared to WT L. monocytogenes (Rea et al., 2005).
NmlR is a MerR-type transcription regulator first characterized in Neisseria gonorrhoeae (so-called NmlRng), and a homolog of the NmlR regulator was also identified in L. monocytogenes (Kidd et al., 2005). NmlR can act as an activator and a repressor under different conditions: for example, NmlRng is an activator of adhC under oxidative stress by diamide, while under normal growth conditions, the NmlR acts as a repressor (Kidd et al., 2005). Roles of L. monocytogenes NmlR (or NmlRlm) regulator encoded by lmo1478 in strain EGD-e (or LMRG_00931 in strain 10403S) gene could clarify how L. monocytogenes responds to different environments, specifically in the switch from saprophytic to pathogenic stages. This study focused on the role of NmlRlm in stress response and the characterization of NmlRlm regulon.
Materials and Methods
Bacterial strains
L. monocytogenes strain 10403S was used as a wild-type (WT) strain in this study. Escherichia coli strain DH5α was used as a host cell for plasmid transformation. Under routine growth conditions, L. monocytogenes was cultured in the brain-heart infusion (BHI) medium (Difco) and E. coli was grown in the Luria-Bertani medium (Difco) at 37°C with 200 rpm shaking. For long-term storage, 20% sterilized glycerol was added into the overnight culture and stored at −80°C.
Mutant construction
A 285 bp in-frame deletion using splicing by overlap extension polymerase chain reaction (SOE-PCR) and allelic exchange mutagenesis was undertaken to construct a ΔnmlRlm
mutant. The procedures used for mutant construction were similar to previous studies (Wiedmann et al., 1998). The SOE-PCR primers used in this study are shown in Supplementary Table S1 (Supplementary Data are available online at
RNA isolations for RNA sequencing, quantitative real-time polymerase chain reaction, and 5′ RACE
To allow comparisons of transcriptomes from previous studies (Hu et al., 2007; Oliver et al., 2009; Chaturongakul et al., 2011), each RNA sample from WT or ΔnmlRlm was isolated from an early stationary-phase culture defined as OD600 of 1.0 with an additional 3-h incubation (Chaturongakul et al., 2011).
For RNA samples used in RNA sequencing (RNAseq), the RNAlater® Solution (Ambion) was added to the bacterial cell culture. A RiboPure RNA extraction kit (Ambion) was used for RNA extraction and DNase treatment. The level of DNA contamination was evaluated by quantitative real-time polymerase chain reaction (qRT-PCR) on the housekeeping rpoB gene. An Agilent 2100 Bioanalyzer and an Agilent RNA 6000 Pico Kit (Agilent Technologies) were used to determine RNA quality and quantity. rRNA depletion was performed using a MICROBExpress ™ Bacterial mRNA Enrichment Kit (Ambion) and confirmed by the Agilent 2100 Bioanalyzer.
For RNA samples used in qRT-PCR and 5′ RACE, early stationary-phase cultures were collected in 10% acid phenol:ethanol (Bergholz et al., 2013). RNA was extracted using TRIzol (Invitrogen) and zirconia beads. To eliminate DNA contamination in the RNA sample, DNase (Promega) was used. RNeasy® Mini Kit (Qiagen) was used for RNA clean-up. RNA concentration was measured using a NanoDrop spectrophotometer.
RNAseq and data analysis
An Ion PGM™ Sequencer (Life Technology) was selected for RNAseq. The Ion Total RNA-Seq Kit (Ambion) was used in preparing a whole transcriptome library. The template was prepared using an Ion OneTouch™ 200 Template (Ambion) kit and reagents for sequencing reaction were from an Ion PGM™ Sequencing 200 kit.
To prepare for data analysis, three C++ programs written on Linux were used to convert raw sff files, determine sequence basic information, and trim sequences. Raw sff files from the Ion PGM sequencer were converted to fastq files. Fastq files were used to determine basic information (i.e., total bases and total reads) and sequence quality using the FastQC program (
The raw sequence data (.bam files) of all samples and summary of RNAseq results (with RPKM values) were submitted to GEO data repository under accession number GSE70265. For small RNA annotation, small RNAs were retrieved from the Broad Institute L. monocytogenes Genome Database website (Listeria monocytogenes Sequencing Project, Broad Institute of Harvard and MIT;
Quantitative real-time polymerase chain reaction
After RNAseq analysis, 11 genes that showed significant changes in expression levels (fold change >9) when compared between WT and ΔnmlRlm mutant were selected to perform qRT-PCR (i.e., cheA, ftsE, groS, hrcA, infC, LMRG_0412, pdxS, purK, rpoA, rpoB, and trxB). The experiments were performed in three biological replicates. Expression level of the lmo1477 gene, located divergently upstream from nmlRlm and previously suggested to be NmlR dependent in other organisms (Kidd et al., 2005), was also assessed. The transcript level of a housekeeping gene gap, encoding for glyceraldehyde-3P-dehydrogenase, was used for normalization. qRT-PCR XS-probes (Biolegion) and primers (Supplementary Table S1) were designed for this study by the Primer Express program (Applied Biosystems). Standard curves for copy number determination of RNA expression were calculated from threshold cycles of varying copy numbers of WT genomic DNA. TaqMan® RNA-to-CT1-step kits (Ambion) were used in all qRT-PCR. qRT-PCR was performed in ABI7500 (Applied Biosystems), and ABI7500 SDS v1.5.1 software was used to analyze the results.
NmlRlm promoter mapping
To determine the transcriptional start site of nmlRlm , 5′ RACE was performed. A Transcriptor High Fidelity cDNA Synthesis kit (Roche) was used for cDNA synthesis. Primer sequences used in 5′ RACE are shown in Supplementary Table S1. An RNase H (Promega) was added to cDNA to digest any remaining RNA, and cDNA was purified using the QIAquick® PCR Purification kit. Tdt (Promega) was used to add A-tails, and then, the sample was used as a template in the first round PCR with PRTN and GSP-1 primers. The purified PCR product was used as a template for the second round PCR with PM1N and GSP-2 primers. The second round PCR product was ligated to pGemT Easy vector (Promega) by T4 Ligase (Promega). Ligated pGemT Easy was transformed into DH5α competent cells. Plasmid was extracted from the transformant using a QIAprep® Spin Miniprep Kit (Qiagen) and sent to determine the insert sequences using universal T7 promoter and Sp6 promoter primers (Macrogen).
Stress survival assays
L. monocytogenes WT, ΔnmlRlm , and cnmlRlm , were used in oxidative, nitrosative, acid, and alkali stress assays. Oxidative stress agents used in this study were cumene hydroperoxide (CHP; Fluka), hydrogen peroxide (H2O2; Sigma), and sodium hypochlorite (NaOCl; Sigma). One milliliter of early stationary-phase culture was treated with each stress agent. Both treated and untreated cultures were incubated at 37°C for 15 min and 60 min with 200 rpm shaking. For nitrosative stress, early stationary-phase cultures were treated with S-nitrosoglutathione (GSNO; Sigma). Both treated and untreated cultures were incubated at 37°C for 60 min with 200 rpm shaking. For acid and alkaline stress conditions, 12 M HCl was added until the pH of the bacterial culture reached 2.5 and 12 M NaOH was added until the pH rose to 12, respectively. Treated and untreated cultures with acid and alkali stress agents were incubated at 37°C for 60 min with 200 rpm shaking. After incubation, treated and untreated cultures were spread on BHI plates for colony-forming unit (CFU) enumeration. The experiments were repeated at least three times.
Intracellular growth assay
Human myelomonocytic cell line U937 was used to study intracellular growth of L. monocytogenes (Caron et al., 1994; Robbins et al., 2010). For routine maintenance of U937, the Roswell Park Memorial Institute 1640 medium (RPMI 1640) (Gibco) was added with 10% fetal bovine serum (Gibco), 1% final concentration of 100×-penicillin/streptomycin (Gibco), and 5 mM of
Transmission electron microscopy
L. monocytogenes WT, ΔnmlRlm , and cnmlRlm mutant strains were analyzed by transmission electron microscopy (TEM; Center of Nanoimaging, Faculty of Science, Mahidol University). The length and width of each bacterial strain were measured using the program ImageJ 1.48 (Schneider et al., 2012).
Statistical analyses
Results from all phenotypic characterization assays were compared for differences in survival abilities and in sizes through SPSS 18.0 program using T-tests and analysis of variance (ANOVA) at 95% confidence interval. Tukey's honest significant difference was used in post-hoc analysis of multiple comparisons. Statistical significance was established at a p-value <0.05.
Results
Regulation of nmlRlm gene and characterization of NmlRlm regulon during early stationary phase
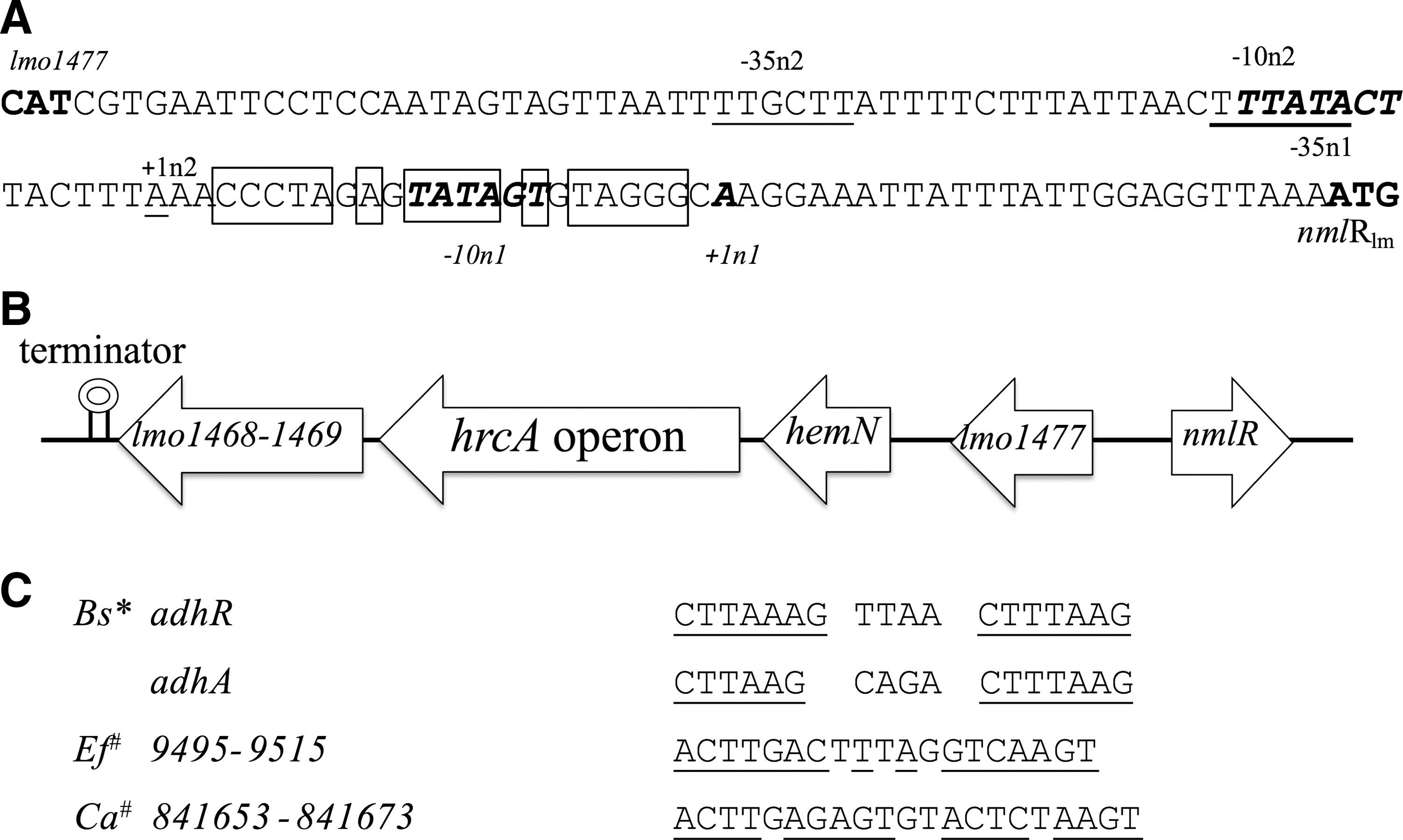
Transcriptional start sites of nmlRlm were determined by 5′ RACE using two gene-specific primers GSP-1 and GSP-2. Amplification with GSP-2 primer yielded two PCR product sizes, and sequencing results suggested two transcriptional start sites. Two putative promoter regions were deduced as shown in Figure 1A. NmlR binding sites typically locate in dyad symmetry (Fig. 1C). The putative dyad for NmlRlm is shown in Figure 1A.
RNAseq using Ion PGM™ sequencer was used to determine transcriptomes of WT and ΔnmlRlm strains. Details on sequence outputs are shown in Supplementary Tables S2 and S3. EDGE-Pro program was used to identify differentially expressed genes between WT and ΔnmlRlm strains (>9-fold difference by RNAseq). The 74 genes identified are listed in Table 1. Forty-six genes were expressed at higher levels in the ΔnmlRlm strain than in the WT strain, indicating negative regulation associated with NmlRlm; while 28 genes were expressed at higher levels in the WT than in the ΔnmlRlm mutant, indicating positive regulation associated with NmlRlm. One small RNA with higher expression level (>5-fold difference) in the ΔnmlRlm mutant than in the WT was identified, that is, glucosamine-6-phosphate-activated ribozyme glmS.
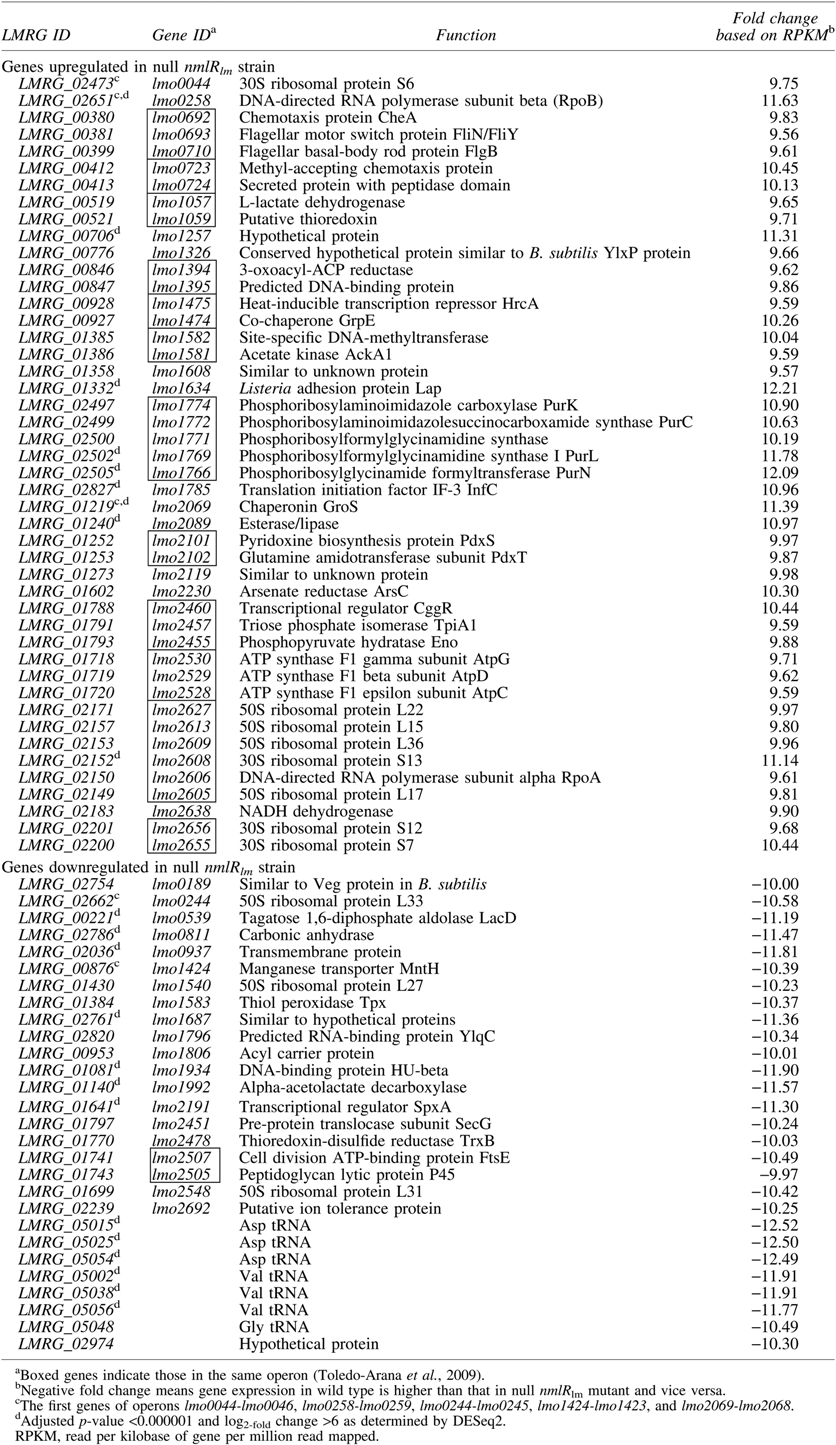
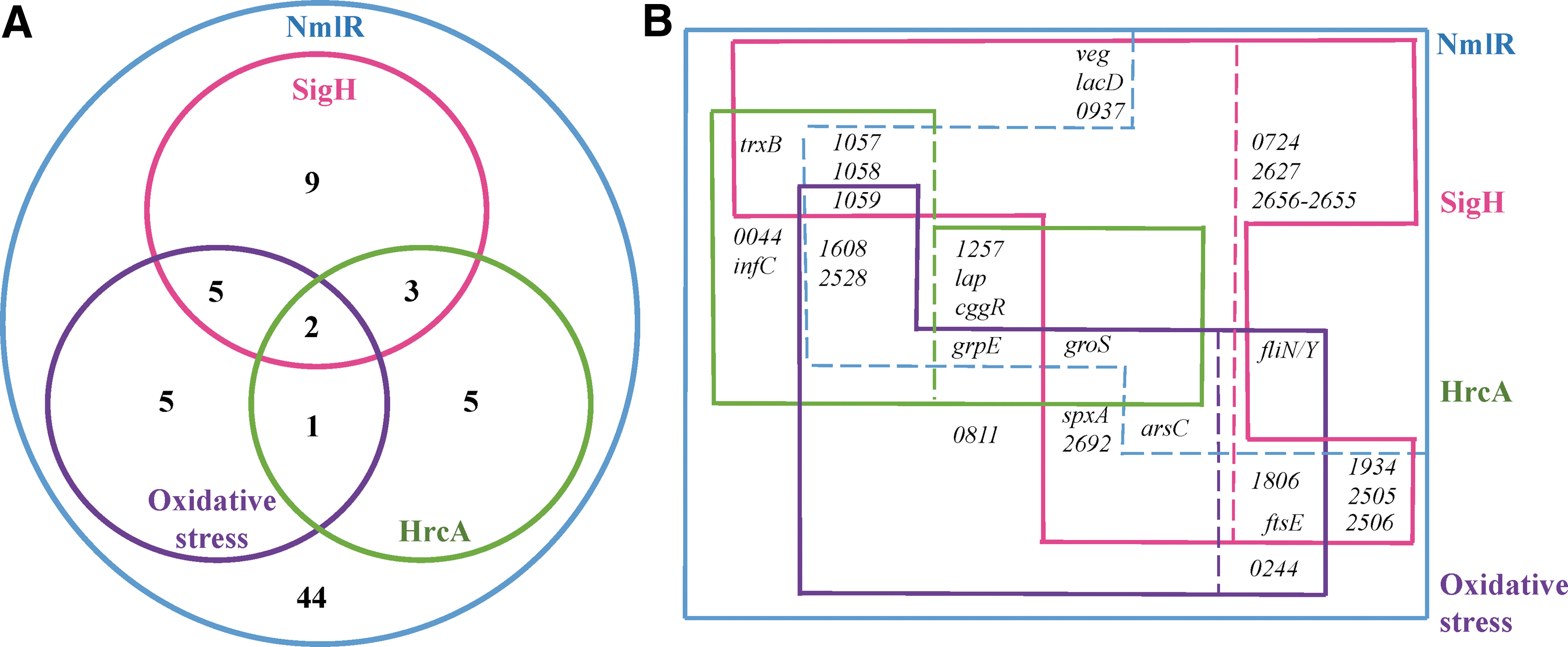
NmlRlm-regulated genes were grouped into operons by comparing with identified operons of L. monocytogenes EGD-e (Toledo-Arana et al., 2009; Caspi et al., 2014). Among 74 NmlRlm-regulated genes, at least 13 operons were identified (Table 1). RNAseq also showed that key regulators in L. monocytogenes are regulated by NmlRlm, for example, stress response regulator HrcA, chaperonin GroS, and central glycolytic gene regulator (CggR), suggesting the role of NmlRlm in both survival under stressful conditions and energy metabolism (Table 1). By comparing the NmlRlm regulon to those previously identified (Hu et al., 2007; Chaturongakul et al., 2011), we found that 30 of the NmlRlm-dependent genes overlapped with those in HrcA, σH, and oxidative stress (ClO2) regulons (Fig. 2).
qRT-PCR was performed to confirm the results from RNAseq. Eleven genes were selected: nine upregulated (cheA, groS, hrcA, infC, LMRG_0412, pdxS, purK, rpoA, and rpoB) and two downregulated (ftsE and trxB) in the ΔnmlRlm mutant. Although the RNAseq result suggested that NmlRlm regulated expression of the cggR, the first gene in cggR-gap-pgk-tpi-pgm-eno operon, the expression levels of the gap gene, a housekeeping gene used in qRT-PCR normalization in previous studies, were not significantly different in WT (5.78 ± 0.14 log copy number) and mutant (5.70 ± 0.97). Therefore, we continued to use the gap gene for qRT-PCR normalization. All selected genes showed changes in expression levels of more than 1.5-fold, the cutoff value for determination of differential expression by qRT-PCR. The infC gene showed the highest increase in fold change at 9.94 in the ΔnmlRlm mutant compared to the WT, while the cheA gene showed the lowest degree of fold change at 1.51. All selected genes for qRT-PCR correlated with the results from RNAseq except for the trxB gene (Supplementary Table S4). In RNAseq, the trxB gene was expressed at a higher level (10.03-fold) in WT than in the ΔnmlRlm mutant, while in qRT-PCR, trxB expression was higher in the ΔnmlRlm mutant than in the WT strain (1.80-fold).
Phenotypic characterization of ΔnmlRlm strain
To investigate the role of NmlRlm in oxidative stress response, three oxidants, that is, CHP, H2O2, and NaOCl were used in survival assays of L. monocytogenes WT, ΔnmlRlm , and cnmlRlm strains. The average log reductions of WT, ΔnmlRlm , and cnmlRlm strains along with corresponding standard deviations are shown in Table 2. The ANOVA test showed no significant differences at the 95% confidence interval among strains treated with 13 mM CHP and 0.02% NaOCl (Table 2). A 15-min treatment of oxidative stress by 3% H2O2 resulted in no statistical difference between the three strains. The average log reductions of WT, ΔnmlRlm , and cnmlRlm strains after a 60-min treatment of H2O2 showed significant differences (p = 0.002) (Table 2). The ΔnmlRlm mutant survived significantly less than the WT (p = 0.003). No significant difference was observed between the WT and the complemented strain. To investigate the role of NmlRlm in nitrosative stress response, the GSNO survival assay was performed on all three strains. After 60 min of treatment, the ΔnmlRlm mutant survived significantly less than the WT (p = 0.049) (Table 2). However, survival was not restored to the WT level in the complemented strain.
Log reduction = log CFU/mL of untreated sample − log CFU/mL of treated sample.
Statistical groups with p < 0.05 as a cutoff for significant difference.
CHP, cumene hydroperoxide; H2O2, hydrogen peroxide; NaOCl, sodium hypochlorite; GSNO, S-nitrosoglutathione; SD, standard deviation; CFU, colony-forming unit.
HCl and NaOH were used to investigate the role of NmlRlm in pH stress response. Average log reductions of WT, ΔnmlRlm , and cnmlRlm strains after the 60-min treatment with HCl are shown in Table 2. Log reductions were significantly different among the three strains (p = 0.015). The ΔnmlRlm mutant survived significantly less than the WT at pH 2.5 (p = 0.013). No significant difference was observed between WT and the complemented strain. Under alkali stress (pH 12), no significant difference was observed among tested strains (Table 2).
To investigate the role of NmlRlm in intracellular growth, WT, ΔnmlRlm , and cnmlRlm strains were inoculated into differentiated U937 cell cultures. Log CFU/mL values of all strains at different time points are shown in Figure 3A. After 6 h of incubation, ΔnmlRlm strains grew at higher counts than the WT strain. At 8 and 10 h after inoculation, the ΔnmlRlm mutant grew significantly at higher counts than the WT (p = 0.012 and 0.016, respectively). However, at 12 h, growth of the WT strain caught up with the ΔnmlRlm mutant and there was no significant difference. Complementation could slightly restore the intracellular growth characteristics during entry to stationary phase.
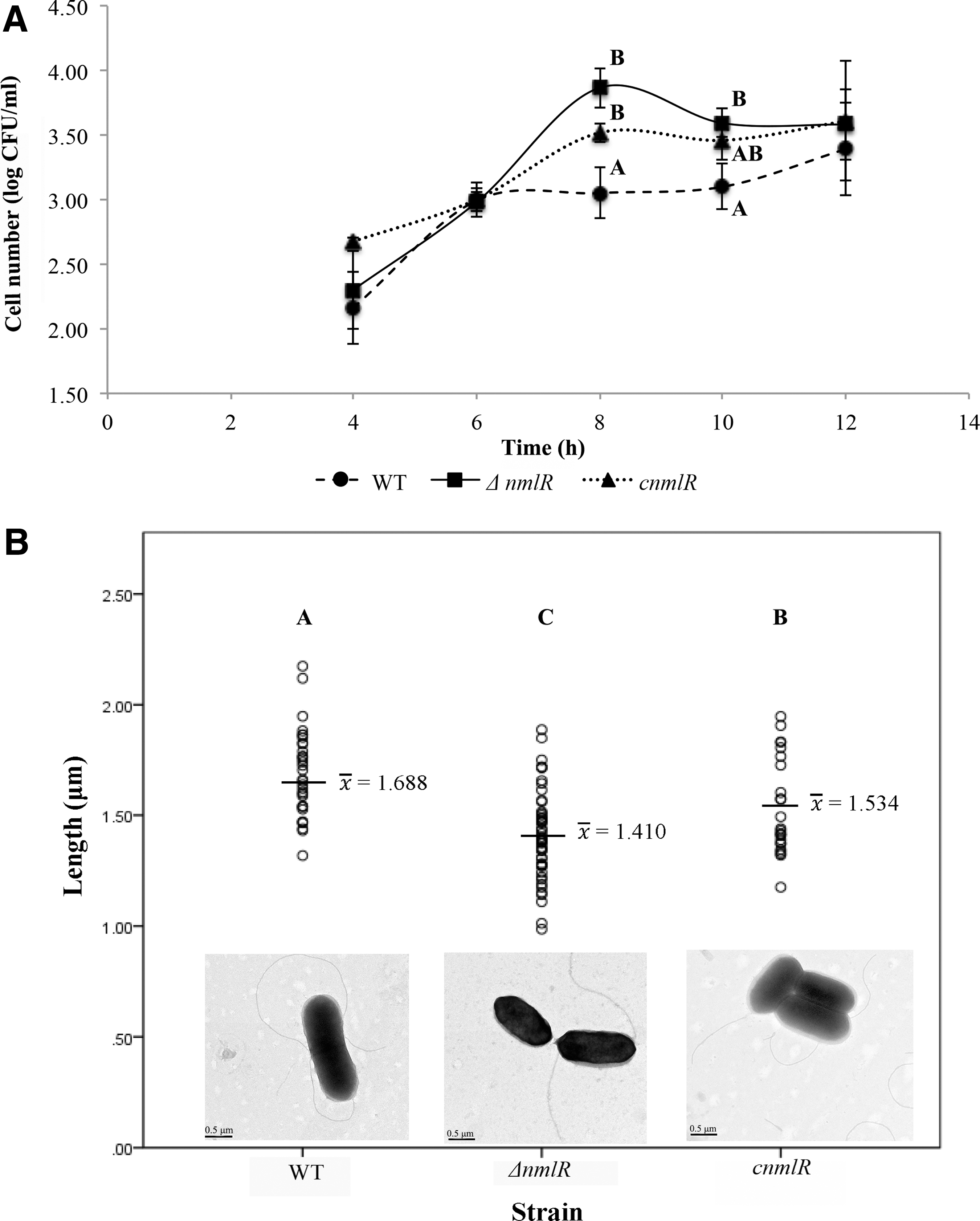
Finally, to assess the role of nmlRlm on the cell morphology, all strains were observed under TEM. Each bacterial cell was measured in length and in width (Table 2). Figure 3B shows example pictures from TEM. The ΔnmlRlm strain was significantly shorter than the WT strain (p < 0.001) and complementation could partially restore the length. The widths are not statistically different among strains.
Discussion
Homologs of the NmlR regulator were found in at least eight species of bacteria: N. gonorrhoeae, N. meningitidis, H. influenzae, Lactobacillus plantarum, Oceanobacillus iheyensis, Desulfitobacterium hafniense, B. subtilis, and L. monocytogenes (Kidd et al., 2005). Previous studies on the NmlR homolog in N. gonorrhoeae found NmlRng to be involved in oxidative stress response as the nmlRng mutant exhibited a decrease in survival under oxidative stress condition created by cumene hydroperoxide and diamide (Kidd et al., 2005). In H. influenzae, disruption of nmlRhi resulted in impaired growth under nitrosative stress condition (0.5 mM GSNO) (Kidd et al., 2007). In this study, the aim was to characterize the role and the regulon of NmlRlm in the foodborne and facultative intracellular pathogen L. monocytogenes 10403S. Physiological stresses related to food and gastrointestinal infection such as acid stress and intracellular growth were included. Phenotypic characterization showed that the nmlRlm mutant survived significantly less than the WT under acid, oxidative, and nitrosative stress conditions. Deletion of nmlRlm also affected cell length and intracellular growth. These phenotypes suggest the roles for NmlRlm in stress response and also in host infection.
Previous studies in other organisms have shown that NmlR directly regulates the gene divergently upstream of the nmlR gene (Kidd et al., 2005, 2007; Huyen et al., 2009). The promoter overlap and dyad symmetry sequences of NmlR binding sites are shared (Kidd et al., 2005, 2007; Huyen et al., 2009). Promoter overlap and dyad symmetry are found between lmo1477 and nmlRlm gene as shown in Figure 1A. Unlike previous studies, this work shows that NmlRlm positively regulates lmo1477 (or LMRG_00930). Based on qRT-PCR, expression levels of lmo1477 were higher in the WT than in the ΔnmlRlm mutant during the stationary phase. This could be due to the fact that NmlR can act, on the same promoter, both as a repressor under nonstressed conditions and as an activator under stressed conditions (Kidd et al., 2005, 2007). In L. monocytogenes, the stationary-phase condition could be enough to impose stress and induce NmlRlm activity. However, it is important to note that lmo1477 was not identified as NmlRlm dependent by RNAseq.
This study also supports earlier findings that NmlRlm is another stationary-phase regulator, among other key regulators such as σB, σH, and HrcA (Hu et al., 2007; Oliver et al., 2009; Chaturongakul et al., 2011). RNAseq showed that NmlRlm and σH regulate chemotaxis and motility through negative regulation of a 28-membered cheY operon, purine ribonucleotide biosynthesis (purE operon), and protein synthesis (rpsJ operon). This suggests that NmlRlm, like σH, is involved in the transition from the exponential phase to the stationary phase, and a network of gene regulation between NmlRlm and other regulators is complex.
In comparison to an oxidative stress regulon characterized in L. monocytogenes 10403S after exposure to chlorine dioxide (Pleitner et al., 2014), 13 genes from NmlRlm regulon overlap (Fig. 2A). Specifically, grpE and groS, which are regulated by NmlRlm, HrcA, and σH, are upregulated under ClO2 exposure. In contrast, spxA, regulated by NmlRlm and σH, is downregulated under ClO2 exposure. These overlaps indicate again a network of gene regulation in stress response (Fig. 2B).
RNAseq also revealed that NmlRlm regulates key regulators involved in L. monocytogenes stress response. In this study, NmlRlm negatively regulated expression of HrcA. A hrcA operon is in close proximity to the nmlRlm gene as shown in Figure 1B. The putative dyad symmetry for NmlRlm binding is between nmlRlm and lmo1477. The NCBI database shows absence of a terminator sequence between lmo1477 and hrcA (Fig. 1B). Therefore, it is possible that NmlRlm directly and negatively regulates hrcA and genes in the hrcA operon. Other regulatory proteins regulated by NmlRlm are CggR, a repressor of the gap operon and glycolysis downregulated by HrcA (Hu et al., 2007). Expression levels of CggR, HrcA, GroS, and NmlRlm have been shown to increase while L. monocytogenes takes on an intracellular life cycle (Gahan et al., 2001; Chatterjee et al., 2006; Joseph et al., 2006; Typas et al., 2012). These regulatory cascades among CggR, HrcA, GroS, and NmlRlm again suggest a complex regulatory network in stress response and during host infection.
Based on the identified NmlRlm regulon, two types of feed forward loop regulatory cascades (Fig. 4) could be demonstrated, that is, incoherent and coherent (Mangan and Alon, 2003). Type 2 incoherent feed forward loops (I2-FFL) were identified as NmlRlm negatively regulates HrcA, CggR, and GroS, and subsequently HrcA negatively regulates CggR and GroS (Hu et al., 2007). I2-FFL can speed up stress response because, without a stimulation factor, NmlRlm can directly repress expression of CggR and GroS by bypassing the HrcA function. Type 2 coherent feed forward loop was identified here as NmlRlm exerted direct and indirect negative regulation on expression of protein synthesis genes (e.g., infC and ribosomal protein encoding genes). The indirect negative regulation was through the negative regulation of NmlRlm on HrcA, a positive regulator of these protein synthesis genes (Fig. 4).
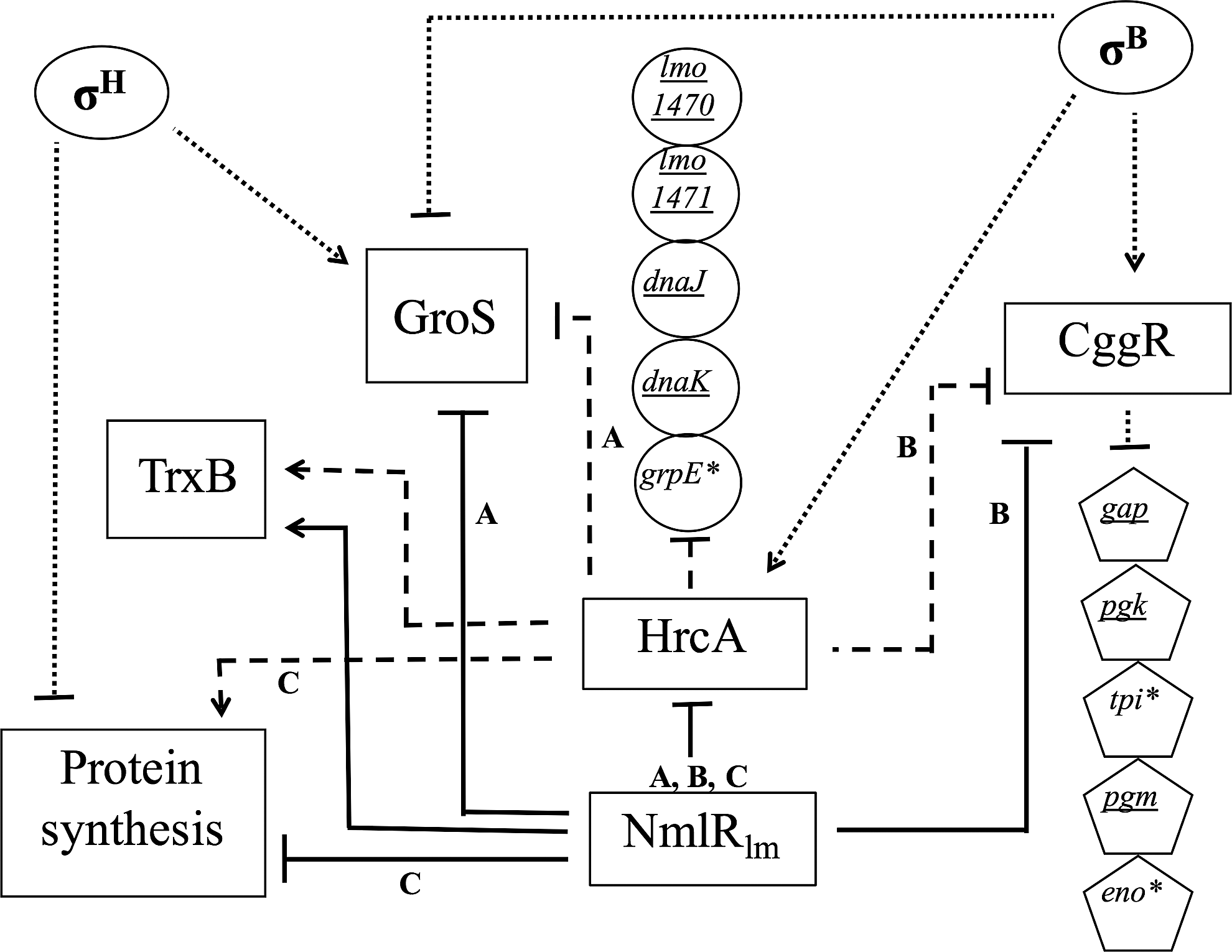
A proposed sub-network of NmlRlm-regulated genes. Target arrows (↓) indicate positive regulation, target stops (⊥) indicate negative regulation. Black solid lines indicate NmlRlm-dependent genes/proteins. Circular shapes indicate genes in hrcA operon. Dashed lines (----) indicate HrcA-dependent genes/proteins (Hu et al., 2007). Pentagon shapes indicate CggR-dependent genes. Dotted lines (….) indicate genes/proteins dependent on σB or σH. Asterisks indicate genes that are member of HrcA-NmlRlm regulons or CggR-NmlRlm regulons. Underlined letters indicate genes indirectly regulated by NmlRlm (via either HrcA or CggR). “A” and “B” indicate type 2 incoherent feed forward loop regulatory cascades while “C” indicates type 2 coherent feed forward loop regulatory cascade.
Conclusions
Most NmlR-dependent genes in previous studies were involved in oxidative, nitrosative, and carbonyl stresses. However, in this study, RNAseq and phenotypic characterization revealed that NmlRlm was also involved in other activities such as acid stress and intracellular growth. Findings from this study strongly suggest that NmlRlm regulates other important stress response and virulence regulators in L. monocytogenes, particularly HrcA, σH, CggR, and GroS, adding to the complexity of L. monocytogenes stress response network.
Footnotes
Acknowledgments
We thank Dr. Skorn Mongkolsuk, Dr. Benjarath Pupacdi, and Dr. Renato H. Orsi for their valuable comments and discussions. We are grateful for Dr. Pongsak Utaisincharoen for providing us the U937 cell line. This work was partially supported (to S.C.) by the Thailand Research Fund and the Office of the Higher Education Commission (MRG5280077).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.