
Abstract
Contamination of food and water with pathogenic bacteria is of concern. Although culture-independent detection and quantification of pathogens is useful, isolation of pathogenic bacteria is still important when identifying the sources of pathogens. Here, we report the use of flow cytometry (FCM) and fluorescence-activated cell sorting (FACS) to specifically detect and isolate individual Escherichia coli O157:H7 cells from water samples. When present at >10 cells/mL water, target pathogen was specifically detected and isolated. The FACS-sorted E. coli O157:H7 population reflected the original population diversity, in contrast to the populations obtained by immunomagnetic separation. Relative abundance of multiple pathogenic strains is important when performing source-tracking studies; therefore, single-cell isolation with FCM–FACS can be a useful tool to obtain pathogenic bacteria for source tracking purpose.
Introduction
C
Although the advancement of molecular tools such as quantitative polymerase chain reaction (qPCR) is useful to detect and quantify pathogens of interest (Quilliam et al., 2011; Ishii et al., 2013; Ramírez-Castillo et al., 2015), isolation of target pathogens is necessary to identify the sources because current source identification protocols for pathogens (e.g., pulsed-field gel electrophotesis and multilocus sequence typing) require pure culture strains (Swaminathan et al., 2001; Pérez-Losada et al., 2013). Historically, isolation of target pathogens has been done using selective media. However, some nontarget bacteria can also grow in selective media and negatively influence the growth of target pathogens (LeJeune et al., 2001). To further increase the specificity of the media, growth inhibitors (e.g., antibiotics) and selective growth conditions (e.g., elevated temperature) can be used to inhibit the growth of nontarget bacteria; however, growth of some target pathogens can also be inhibited. For example, cefixime and tellurite that are frequently used for selective isolation of E. coli O157:H7 can inhibit the growth of some E. coli O157 strains (Bielaszewska et al., 2005). Antibiotics and elevated temperature conditions can also suppress the growth of starved E. coli O157 strains (Sata et al., 1999).
Another approach to separate target pathogens from background population is to use pathogen-specific antibodies conjugated with magnetic beads (i.e., immunomagnetic separation [IMS]). This approach has been successfully used to isolate pathogens from food and water samples (Quilliam et al., 2011), although their low sensitivity may require an enrichment step before IMS. Multiple genotypes of pathogenic bacteria can be present in water and other environmental samples (Jay et al., 2007), and their relative abundance (i.e., population diversity) is important to consider when identifying the sources of pathogens (Ishii et al., 2007). Relative abundance of bacteria strains can be altered after culture enrichment in broth media. Therefore, the current IMS procedure, which requires an enrichment step, is not suitable to isolate target pathogens for source identification purposes.
Single-cell isolation and cultivation techniques separate individual cells before cultivation; therefore, theoretically it should reflect original population diversity (Ishii et al., 2010). Among various single-cell isolation procedures, flow cytometry (FCM) with fluorescence-activated cell sorting (FACS) function is promising because it is high throughput and can analyze multiple parameters (Davey and Kell, 1996; Müller and Nebe-von-Caron, 2010). FCM has been used to detect pathogens in food and water samples (Tortorello et al., 1997; Vives-Rego et al., 2000; Yamaguchi et al., 2003; Hahn et al., 2008). However, it is still challenging to apply FCM–FACS to the isolation of pathogens from environmental samples because of the low concentrations of target pathogens, large background populations, and high autofluorescent signals present in environmental samples. Also, it is not well examined if the sorted population reflects original population diversity.
The objectives of this study were (1) to optimize FCM–FACS to specifically detect and isolate E. coli O157 cells, (2) to compare the strain compositions of the E. coli O157 populations before and after FCM–FACS, and (3) to apply the FCM–FACS approach to isolate the pathogens from environmental water samples.
Materials and Methods
Bacterial strains
Non-Shigatoxigenic E. coli O157:H7 strain LMG 21756 was obtained from Belgian Coordinated Collections of Microorganisms (BCCM). Kanamycin-resistant mutant of strain LMG 21756 (named as strain LMG 21756 KmR) was prepared by transposon mutagenesis as described previously (Kim et al., 2014). E. coli K12 strain MG1655 was used as non-O157 control.
E. coli strains were grown in brilliant green bile broth (BGBB; Oxoid, Hampshire, United Kingdom) at 37°C. Cell densities of the E. coli strains were determined by measuring the optical density at 600 nm (OD600) and direct cell counting after 4′,6-diamidino-2-phenylindole (DAPI) staining.
Specific staining of E. coli O157 cells
Fluorescein isothiocyanate (FITC)-labeled anti-O157 monoclonal antibody (#1063) was purchased from ViroStat (Portland, ME). Allophycocyanin (APC)-labeled anti-O157 monoclonal antibody was prepared by labeling anti-O157 antibody (#1031 from ViroStat) using allophycocyanin labeling kit-SH (Dojindo, Mashiki, Kumamoto, Japan) according to the manufacturer's instruction.
The FITC- and APC-labeled antibodies were added to 1 mL of bacteria cell suspension at final concentrations of >3 × 10−6 and >3 × 10−7 μg/cell, respectively, and incubated for 20 min on ice. These concentrations of antibodies were proven sufficient to stain most of the E. coli O157 cells (Supplementary Fig. S1; Supplementary Data are available online at
Single-cell isolation of E. coli O157 by FCM–FACS
Stained E. coli cells were used for FCM–FACS analyses done with a JSAN desktop cell sorter (Bay Bioscience, Kobe, Japan). FCM data were analyzed using AppSan software (Bay Bioscience), and the O157-specific area was set based on the forward scatter, side scatter, as well as FITC and APC fluorescent signals obtained by the pure culture analysis of strain LMG 21756.
Cells appeared in the O157-specific area by FCM analysis and were individually sorted into 100 μL BGBB medium in 96-well plates. After overnight incubation at 37°C, cells were stamp inoculated onto Sorbitol MacConkey (SMAC) agar (Difco, Detroit, MI) and CHROMagar O157 (CHROMagar, Paris, France) plates. Transparent and pink colonies on SMAC agar and CHROMagar O157, respectively, were considered positive for E. coli O157.
Isolation of E. coli O157 by using IMS
E. coli O157 cells were also isolated using Dynabeads anti-E. coli O157 (Applied Biosystems, Waltham, MA) according to the manufacturer's instruction. For this experiment, we prepared E. coli cell mixtures (104 cells/mL) composed of strains LMG 21756-KanR and LMG 21756 wild type with a known ratio (10:0, 9:1, 5:5, 1:9, and 0:10, respectively). The same cell mixtures were used for both FCM–FACS and IMS analyses to see whether the ratios of the two strains remain similar after isolation.
In brief, mixtures of E. coli O157 cells were preincubated in tryptic soy broth (TSB; Difco) at 37°C for 8 h. After IMS by the Dynabeads kit, captured cells were suspended and diluted 1000-fold with phosphate-buffered saline (PBS). Aliquots (20 μL) of the cell suspension were spread onto SMAC agar. Colonies grown on SMAC agar were randomly picked and transferred onto SMAC agar supplemented with 100 mg/L kanamycin to verify strain identity.
Water samples
Environmental water samples were collected from Atsubetsu River (Sapporo, Japan; lat 43.091114, long 141.469679) on November 13, 2012, by using surface-disinfected plastic bucket. The physicochemical characteristics of the water samples were as follows: temperature = 8.8°C, pH 7.5, electrical conductivity = 21.8 mS/m, dissolved oxygen = 6.60 mg/L, biological oxygen demand = 0.9 mg/L, suspended solids = 7.1 mg/L, total N = 1.18 mg/L, total P = 0.14 mg/L, total coliform = 2400 MPN/100 mL, and total E. coli = 11 MPN/100 mL.
Soon after water sampling (<1 week), E. coli O157 strain LMG 21756 was inoculated to the water at concentrations of 1000, 100, or 10 cells/mL. The water samples (1 L) were filtered through 0.22-μm-pore polyethersulfone membrane filters (Millipore), and bacterial cells were detached from the membrane by vigorously shaking in PBS (pH 7.2) with 0.1% gelatin (Hamilton et al., 2010). The cells were centrifuged at 10,000 × g for 15 min and resuspended in 1 mL of PBS as described previously (Ishii et al., 2013; Kobayashi et al., 2013).
To separate cells from mineral particles, 500 μL of 60% (w/v) Nycodenz (Axis-Shield, Dundee, United Kingdom) was gently poured onto 1 mL of the concentrated samples. After centrifugation at 10,000 × g for 60 min, low to middle density solution (1 mL) was collected in a new tube and mixed with 9 mL of PBS. After centrifugation at 10,000 × g for 60 min, pellets were resuspended with 1 mL of PBS.
Statistical analysis
All experiments were repeatedly performed (at least two to three runs) to see the reproducibility of the results. Fisher's exact test was performed to test whether the expected and observed values were significantly different or not by using R version 3.2.3.
Results and Discussion
E. coli O157:H7 was specifically detected by FCM similar to the previous studies (Tortorello et al., 1997; Vives-Rego et al., 2000; Yamaguchi et al., 2003; Hahn et al., 2008). We used two antibodies each labeled with FITC and APC to increase the specificity of target detection. Preliminary experiments were performed to optimize the antibody-staining procedures (e.g., antibody concentrations and reaction time) (Supplementary Fig. S1). As a result, stained E. coli O157:H7 cells were clearly distinguished from nonstained O157 cells and E. coli K12 cells both by microscopy (Supplementary Fig. S2) and FCM analysis (Fig. 1). E. coli O157 strains other than strain LMG 21756 could also be detected by this approach (Ozawa, 2013).
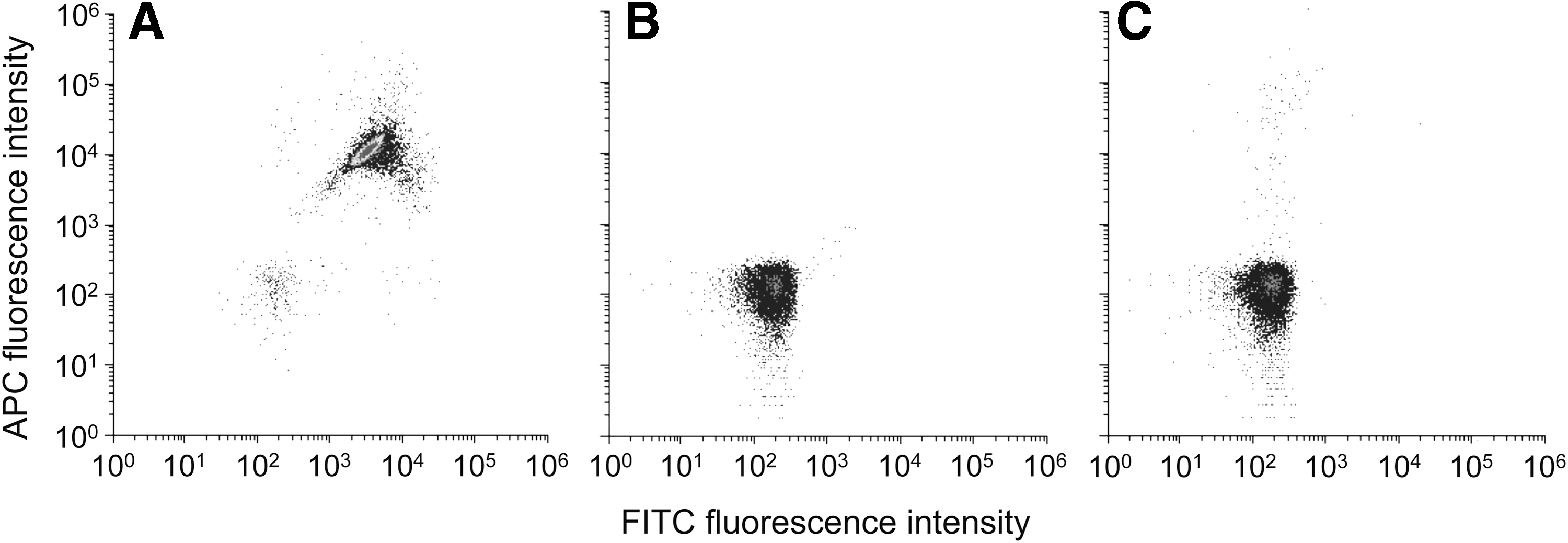
Flow cytometric analysis of Escherichia coli cells.
One advantage of FCM is its capability of analyzing multiple parameters (Davey and Kell, 1996; Müller and Nebe-von-Caron, 2010). For example, Yamaguchi et al. (2003) used FITC-labeled anti-O157 antibody and 5-cyano-2,3-ditolyl tetrazolium chloride (CTC) to detect respiring E. coli O157:H7 cells. In our preliminary experiments, we also stained live E. coli O157:H7 cells with FITC-labeled anti-O157 antibody and 7-isobutyloxycarbonyloxy-3H-phenoxazin-3-one (CytoRed). This dual staining worked fine, although specificity of E. coli O157:H7 was relatively low due most likely to the nontarget fluorescence present in environmental water samples. We therefore used two antibodies that recognize different sites of O157 antigen to increase specificity of detection.
A linear relationship was observed between quantities of E. coli O157:H7 used and those counted by FCM when concentrations were >103 cells/mL (Fig. 2), suggesting that the target cells were quantitatively detected by FCM. Background fluorescent signals could cause false positives when E. coli O157:H7 concentrations were below 103 cells/mL similar to the previous study (Tanaka et al., 2000).
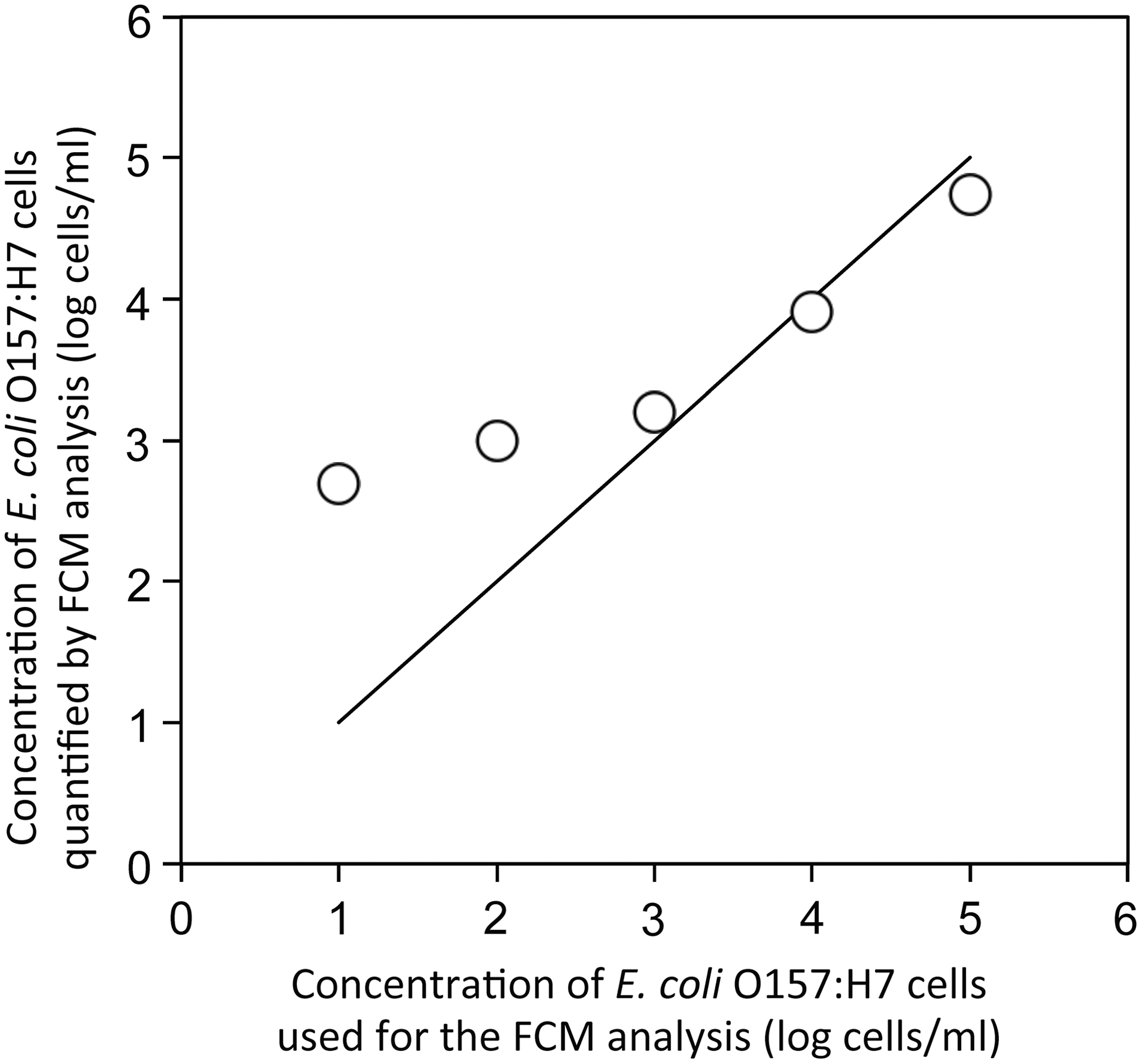
Relationship between concentrations of Escherichia coli O157 strain LMG 21756 (○) used for the FCM analysis and those quantified by FCM analysis. Solid line indicates the ideal relationship (y = x). FCM, flow cytometry.
By using FCM–FACS approach, we could specifically isolate E. coli O157:H7 from the mixture of E. coli O157:H7 strain LMG 21756 and K12 strain MG1655 cells. When the proportions of E. coli O157:H7 were 50%, 9.1%, 1.0%, 0.1%, and 0.01% in the original E. coli cell mixture (106 cells/mL), those in the sorted populations were 97.2%, 91.7%, 61.1%, 39.9%, and 13.3%, respectively (Fig. 3A). This indicated that we could specifically detect target pathogens even when they are present at relatively low concentrations (0.01% of the total population [106 cells/mL] = 100 cells/mL). This detection limit is similar to those reported previously (Yamaguchi et al., 2003).
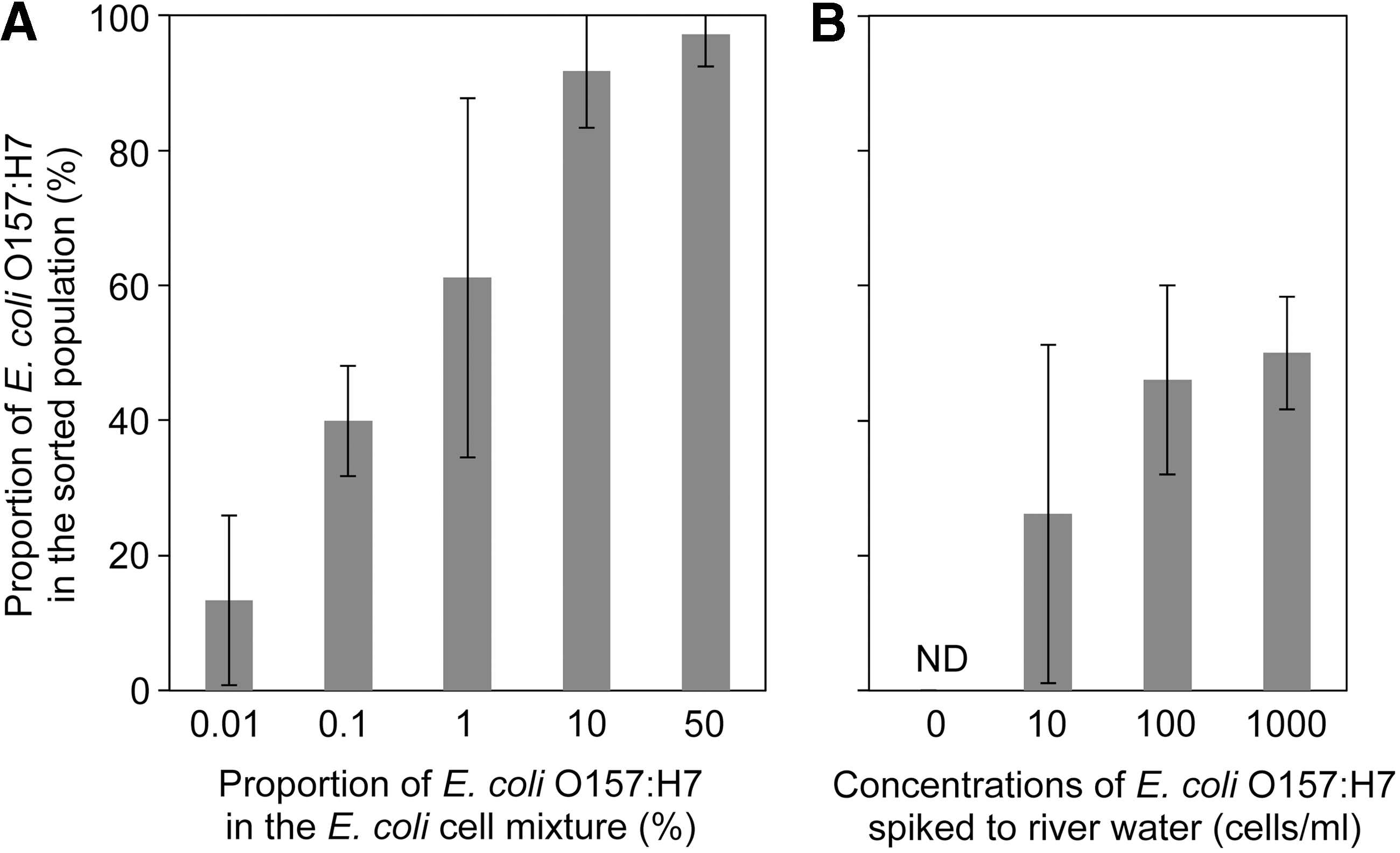
Proportion of Escherichia coli O157:H7 in the sorted populations in
Specific isolation was also possible from environmental water samples. Proportions of E. coli O157:H7 in the sorted populations were 50%, 46%, and 26% when E. coli O157:H7 strain LMG 21756 was spiked to river water samples at 1000, 100, and 10 cells/mL, respectively (Fig. 3B). These values are considerably higher than the stochastic single-cell isolation, given the high levels of background bacteria population in the river water samples (e.g., Okabe et al., 2007). E. coli O157:H7 was not isolated from nonspiked samples (i.e., original river water samples). Thus, the detection limit of the target pathogen was 10 cells/mL water. The pathogen concentrations of >10 cells/mL water can be naturally observed in freshwater environments (Tanaka et al., 2000; Ishii et al., 2014). In the river water samples collected from the urban area in Sapporo, the average concentration of pathogenic E. coli was, on average, 1.9 cells/mL with the highest concentration of 33 cells/mL (Ozawa, 2013). The detection limit can be further lowered by concentrating more water samples (Mull and Hill, 2009). Therefore, although the detection limit was higher than qPCR-based methods (Ishii et al., 2013; Ramírez-Castillo et al., 2015), our FCM–FACS approach is sensitive enough to detect pathogens in water samples without an enrichment step.
The advantages of FCM–FACS over qPCR-based approach are its capability to sort and isolate target cells for further analysis. In this study, the detected pathogen cells were individually sorted and grown in culture medium for further analysis. Because FACS can inoculate target cells individually to culture medium in 96-well format, potential growth of nontarget bacteria is minimum (Ishii et al., 2010). Therefore, we do not have to add selective agents such as antibiotics to the media, which may allow us to recover selective agent-susceptive target cells (Sata et al., 1999; Bielaszewska et al., 2005).
Single-cell isolation by FCM–FACS is also of advantage because the isolated population reflects the original population diversity. In this study, the proportions of the original E. coli O157:H7 populations (i.e., ratio of strains LMG21756 KanR and LMG21756 wild-type strain) were well preserved when FCM–FACS was used as the isolation method (Fig. 4A). Proportions of the two strains were not significantly different between observed and expected values at all ratios (p = 0.40 by Fisher's exact test). In contrast, the proportions of the two strains were changed by IMS with Dynabeads kit (Fig. 4B; p < 0.01) due probably to the enrichment step done before the IMS. Similar results were repeatedly observed (Supplementary Fig. S3). Dynabeads method requires an enrichment step in TSB or other media for 6–18 h according to the manufacturer's instruction. Although enrichment increased the recovery of E. coli O157 in various samples (LeJeune et al., 2001; Verstraete et al., 2010), different strains may compete for the resources in the broth media (Duffy et al., 1999).
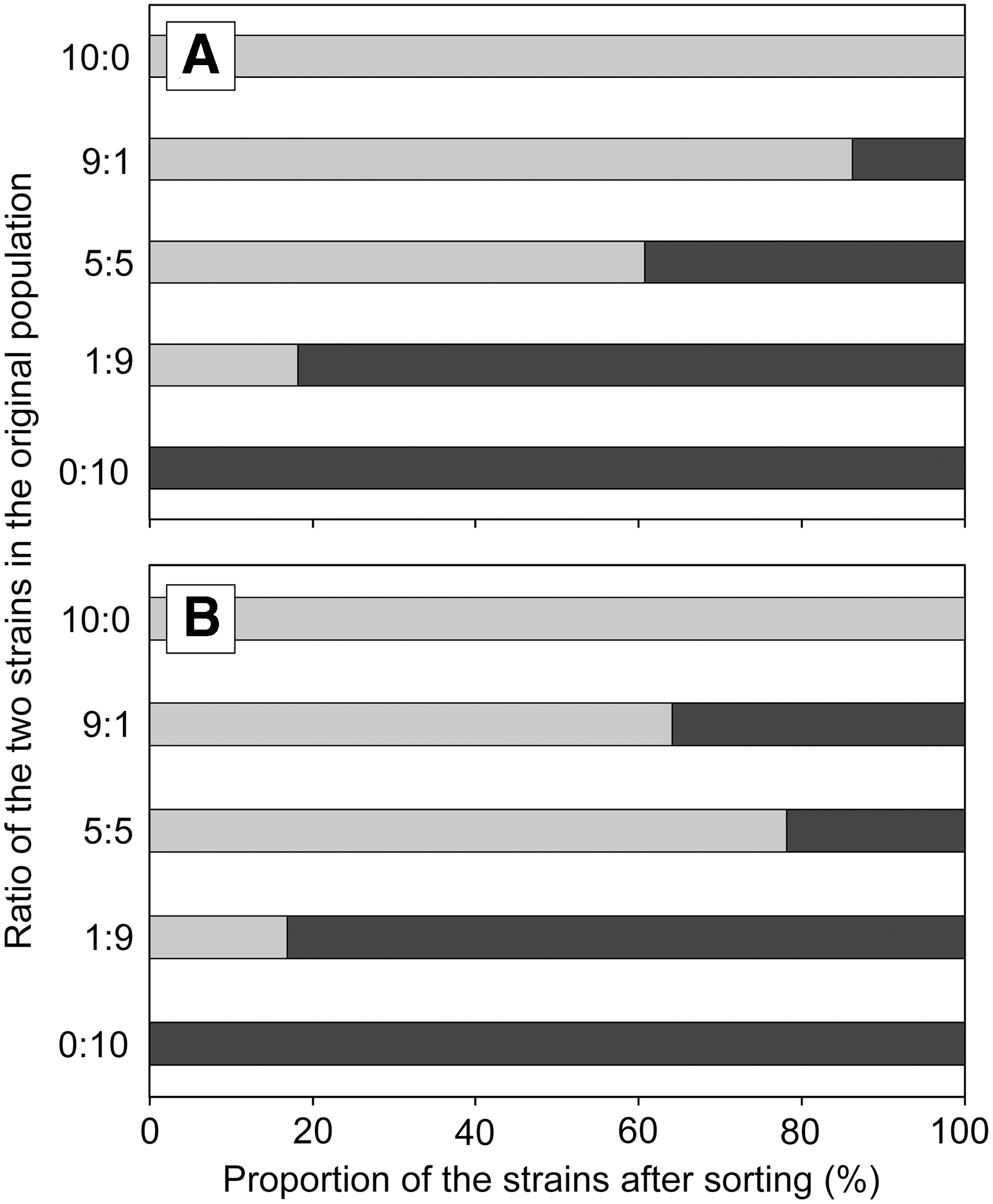
Proportion of the strains after sorting by
Conclusions
Although FACS-based pathogen isolation has been previously performed (Tortorello et al., 1997), they used culture enrichment before FACS. Our study demonstrated that culture enrichment should be avoided when original population diversity needs to be preserved. This is important when identifying the sources of pathogens that most significantly contribute to the pollution. Therefore, single-cell isolation by FACS without culture enrichment is a promising approach for the source tracking of pathogens in the future.
Footnotes
Acknowledgments
We thank Drs. Daisuke Sano and Ayano Kobayashi (Hokkaido University) and Dr. Nobuyuki Kijima (NARO Institute of Vegetable and Tea Science) for valuable comments, and Nora Powers (University of Minnesota) for scientific editing. This study was supported by the Grants-in-Aid for Scientific Research (23860003) from Japan Society for the Promotion of Science (JSPS), the CREST program from Japan Science and Technology Agency (JST), Kurita Water and Environment Foundation Grant, Japan, and the MnDRIVE Initiative of the University of Minnesota.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.