
Abstract
The Pennsylvania Egg Quality Assurance Program (EQAP) provided the framework for Salmonella Enteritidis (SE) control programs, including the Food and Drug Administration (FDA) mandated Final Egg Rule, for commercial layer facilities throughout the United States. Although flocks with ≥3000 birds must comply with the FDA Final Egg Rule, smaller flocks are exempted from the rule. As a result, eggs produced by small layer flocks may pose a greater public health risk than those from larger flocks. It is also unknown if the EQAPs developed with large flocks in mind are suitable for small- and medium-sized flocks. Therefore, a study was performed to evaluate the effectiveness of best management practices included in EQAPs in reducing SE contamination of small- and medium-sized flocks by longitudinal monitoring of their environment and eggs. A total of 59 medium-sized (3000 to 50,000 birds) and small-sized (<3000 birds) flocks from two major layer production states of the United States were enrolled and monitored for SE by culturing different types of environmental samples and shell eggs for two consecutive flock cycles. Isolated SE was characterized by phage typing, pulsed-field gel electrophoresis (PFGE), and clustered regularly interspaced short palindromic repeats-multi-virulence-locus sequence typing (CRISPR-MVLST). Fifty-four Salmonella isolates belonging to 17 serovars, 22 of which were SE, were isolated from multiple sample types. Typing revealed that SE isolates belonged to three phage types (PTs), three PFGE fingerprint patterns, and three CRISPR-MVLST SE Sequence Types (ESTs). The PT8 and JEGX01.0004 PFGE pattern, the most predominant SE types associated with foodborne illness in the United States, were represented by a majority (91%) of SE. Of the three ESTs observed, 85% SE were typed as EST4. The proportion of SE-positive hen house environment during flock cycle 2 was significantly less than the flock cycle 1, demonstrating that current EQAP practices were effective in reducing SE contamination of medium and small layer flocks.
Introduction
N
SE can be transmitted to the egg vertically during its formation and passage through the reproductive tract of the hen (Thiagarajan et al., 1994) or horizontally postlay through the shell due to external contamination (Shivaprasad et al., 1990; Henzler et al., 1998). Therefore, many programs have been developed by state and federal agencies to minimize SE contamination of shell eggs. The National Poultry Improvement Plan developed guidelines under the “SE clean program” to keep breeder flocks free of SE (Federal Register, 2009). Due to the possibility of SE contamination of eggs through horizontal transmission, guidelines were also developed to minimize SE contamination of eggs during production, storage, and transportation (Food and Drug Administration [FDA], 2014b).
The Pennsylvania Egg Quality Assurance Program (PEQAP) was the first Egg Quality Assurance Program (EQAP) developed by any U.S. state (PennState Extension, 2016). This program that relied on Hazard Analysis Critical Control Point (HACCP)-based management practices to reduce the risk of SE contamination of layers and shell eggs has greatly reduced the SE prevalence in commercial layer flocks in Pennsylvania. The PEQAP provided the framework for HACCP-based SE control programs for commercial layer facilities throughout the United States, including the EQAPs developed by the states, such as California (CEQAP), New York (NYSEQAP), Maryland (MDA-EQAP), and Ohio (OEQAP) (Maryland Department of Agriculture, 2004; New York Agriculture and Markets, 2004; Pacific Egg and Poultry Association, 2006; Ohio Poultry Association, 2010).
Despite the efforts put forth by state governments and the table egg industry, the incidence of Salmonella infection in humans continues to stay well above the national health objective target. Therefore, to address this important public health issue, the FDA implemented the Final Egg Rule entitled “Prevention of SE in Shell Eggs During Production, Storage, and Transportation” (FDA, 2014b). Unlike the state EQAPs, the Final Egg Rule is a mandatory program for flocks with more than 3000 birds. This program was implemented in two stages; large flocks with more than 50,000 birds were required to enroll in the program before July 2010 and medium-sized flocks having 3000 to 50,000 birds were required to comply with the rule by July 2012. All these EQAPs have been developed based on the management practices adopted by large production facilities and therefore may not be suitable for medium- and small-sized flocks. Nevertheless, small flocks with less than 3000 birds are exempted from all these programs highlighting the importance of monitoring medium- and small-scale layer farms for SE presence.
The objectives of the present study were (1) to identify the sources of SE contamination of the poultry house environment from medium (3000 to 50,000 birds) and small (<3000 birds) layer flocks, (2) to characterize SE isolated from the poultry house environment and shell eggs using phage typing and pulsed-field gel electrophoresis (PFGE), and (3) to evaluate the effectiveness of best management practices included in EQAPs in reducing SE contamination of layer flocks. This study describes longitudinal monitoring of SE in layer environment and eggs conducted for two consecutive flock cycles in the two major layer production states of Iowa and Pennsylvania.
Materials and Methods
Description of the study flocks
Producers were selected for the study on the basis of voluntary participation from two major egg producing states. A single flock was selected from each producer and the samples were collected after obtaining written permission for sampling at multiple time points over two flock cycles. Forty-three flocks were enrolled for the first laying cycle. Seventeen flocks were withdrawn by the participating producers at the end of the first laying cycle, which necessitated enrolling 17 new flocks for the second laying cycle. Of the selected flocks, 27 had 3000 to 50,000 birds (medium flocks) and 32 had less than 3000 birds (small flocks). To trap the rodents, 12 tin cat mousetraps were provided to each producer to be placed in the layer house. An honorarium was also given to the producers in one state to encourage their participation in the study.
Sample collection
This study was carried out from April 2012 to June 2014. The sampling schedule consisted of collecting chick box papers at day 1 and multiple samples taken at the age of weeks 15, 30, 45, and postmolting. In addition to chick box papers, sample types included hen house drag swabs, feed, water, rodent, fly, and other environmental samples of interest. At the time of sampling during the first laying cycle, none of the farms had an EQAP in place. After cleaning and disinfection, the same sampling schedule was followed for the successive flock. The number and type of samples taken were determined by the size and design of the poultry house and presence or absence of other animal species on the farm. If trapped rodents were not available, rodent feces or swabs from tin cat mousetraps were collected.
The drag swabs were premoistened using pasteurized condensed milk. All samples were collected into Whirl-Pak bags (Fort Atkinson, WI) and transported to the laboratory on the same day or shipped overnight in coolers containing ice packs. If SE was isolated from environmental drag swabs or rodent samples, eggs were collected systematically from the same flock every 2 weeks, four times. The number of eggs to be collected from the farms was defined by the poultry flock size (Supplementary Table S1; Supplementary Data are available online at
Bacteriological culture procedure
Environmental samples
Hen house drag swabs and paddock swabs were processed and cultured according to the Bacteriological Analytical Manual (BAM) Method of FDA (FDA, 2014a).
Rodents
Rodents were taken from Whirl-Pak bags and placed in a quaternary ammonium disinfectant, Roccal II (National Laboratories, Montvale, NJ), to disinfect surface fur. The mouse carcasses were eviscerated aseptically by opening along the ventral midline with sterile instruments. Viscera from up to six mice were pooled together and ground in a sterile blender system. The contents were transferred into a sterile Whirl-Pak bag containing tetrathionate (TT) broth and incubated at 35°C ± 2°C for 18 to 24 h. A loopful (10 μL) of pre-enriched broth was inoculated onto brilliant green with novobiocin (BGN) and xylose lysine tergitol 4 (XLT4) plates and incubated at 35°C for 24 ± 2 h.
Feed
A 100- to 125-mL volume of buffered peptone water was added to a Whirl-Pak bag containing 50 g of feed. The contents in the bag were mixed well and incubated at 35°C ± 2°C for 2 h. A 500-mL volume of TT broth was added to the bag and incubated at 35°C ± 2°C for 18 to 24 h. A 10-μL aliquot from the bag was inoculated onto BGN and XLT4 plates and incubated at 35°C for 24 ± 2 h.
Water
A 10-mg tablet of sodium thiosulfate (Brim Technologies, Randolph, NJ) was added to 200–300 mL of water to neutralize any preexisting treatment with chlorine. The sample was filtered through 250-mL Acro Vac filter units with 0.45-μm pore size (Pall Life Sciences, Ann Arbor, MI). The membrane filter was then removed aseptically from the filter assembly, placed in 100 mL of TT broth, and incubated at 35°C for 24 ± 2 h. A 10-μL aliquot of pre-enriched sample was inoculated onto BGN and XLT4 plates and incubated at 35°C for 24 ± 2 h.
Flies and fly cards or tapes
Flies, fly tapes, and fly cards collected from a single poultry house were pooled and enriched in 100 mL of TT broth as indicated above for other sample types.
Shell eggs
Eggs were tested for Salmonella in pools of 20 eggs according to the FDA-BAM method.
Serotyping and phage typing
Presumptive Salmonella isolates were serogrouped with D1 antiserum, and the isolates that gave a positive reaction were sent to the National Veterinary Services Laboratories (Ames, IA) for serotyping. Isolates that were serotyped as SE were subjected to phage typing.
Pulsed-field gel electrophoresis
PFGE was performed according to the protocol developed by the CDC (2013). Salmonella Braenderup H9812 was used as the molecular marker.
Clustered regularly interspaced short palindromic repeats-multi-virulence-locus sequence typing
SE isolates were assigned a clustered regularly interspaced short palindromic repeats-multi-virulence-locus sequence typing (CRISPR-MVLST) sequence type (ST) according to a method described previously (Liu et al., 2011a, b; Shariat et al., 2013). Briefly, the CRISPR1, CRISPR2, fimH, and sseL loci were amplified with polymerase chain reaction (PCR) using the primers listed in Supplementary Table S2. Purified PCR products were sequenced in both forward and reverse directions using the primers listed in Supplementary Table S2 at the Genomics Core Facility at The Pennsylvania State University. Sequences were assembled and aligned using SeqMan and MegAlign (Lasergene 9; DNA Star, Madison, WI). For all four loci, fimH, sseL, CRISPR1, and CRISPR2, sequences were aligned against previously identified alleles (Shariat et al., 2013). Each isolate was assigned an SE Sequence Type (EST) based on the combination of ST at all four loci.
Statistical analysis
The test on two independent proportions was used to compare the level of SE contamination before and after implementation of EQAP practices in flocks with less than 3000 hens and flocks with more than 3000 hens. The PFGE results were analyzed using BioNumerics software (Applied-Maths, Inc., Austin, TX) by the Unweighted Pair Group Method with Arithmetic Mean method based on the Dice similarity index with a 1.5% band position tolerance. The XbaI restriction patterns were compared with the patterns in the PulseNet database of the CDC.
Results
Study flocks and isolation of SE
The sizes of layer flocks enrolled for the study ranged from 250 to 31,000 birds. All flocks were reared under the cage-free production system and a majority of flocks were vaccinated against SE with live ST and killed SE vaccines. A total of 54 Salmonella isolates belonging to 17 serovars were isolated from multiple sample types. Of these 54 isolates of Salmonella, 22 (40.74%) SE were isolated from hen house drag swabs (n = 10), rodents (n = 6), hen paddock swabs (n = 1), and pooled eggs (n = 5; Table 1). The five SE-positive egg pools originated from four different flocks. Only the environmental swabs and rodents collected at 15 to 45 weeks of flock age were positive for SE. Chick box papers, feed, water, and fly samples were negative for SE throughout the study period.
The farms that were positive for SE during the first flock cycle were free of SE during the second cycle following cleaning and disinfection (Table 2). The hen house environment and eggs were determined to be SE positive, if SE was isolated from at least one hen house swab and one egg pool, respectively. Accordingly, SE occurrence in the small layer flock environment was16.7% and 7.4% during flock cycle 1 and flock cycle 2, respectively, whereas the occurrence in medium flocks were 16% during flock cycle 1 and 0% during flock cycle 2. Occurrence of SE in eggs from small flocks was16.6% during flock cycle 1 and 6.7% during flock cycle 2. Eggs from medium flocks were negative for SE throughout the study (Table 3).
CRISPR-MVLST, clustered regularly interspaced short palindromic repeats-multi-virulence-locus sequence typing; IA, Iowa; Medium, flocks with 3000–50,000 hens; PA, Pennsylvania; PFGE, pulsed-field gel electrophoresis; Small, flocks with <3000 hens; ST, sequence type.
In flocks with less than 3000 hens, SE was isolated from the environmental samples collected during both flock cycles 1 and 2. However, the proportion of SE-positive hen house environment during year 2 was significantly less than during year 1 (flock cycle 1: 3/18, flock cycle 2: 2/15, p < 0.005). Environmental samples collected from medium-sized flocks were positive in flock cycle 1 (4/25) and negative in flock cycle 2 (0/25), respectively. Similarly, the proportion of small flocks producing SE-positive eggs in flock cycle 2 was significantly less than in flock cycle 1 (flock cycle 1: 3/18, flock cycle 2: 1/15, p < 0.005). Eggs from medium-sized flocks were negative for SE during both flock cycle 1 and flock cycle 2 (0/25; Table 3).
Phage types
Three phage types (PTs) of SE were detected in hen house environmental samples and eggs. The PT8 was the major PT that was represented by 19 (91%) isolates of SE originated from nine different layer flocks. SE belonging to PT3 and PT23 were isolated from rodent and hen paddock samples, respectively. All egg-originated SE belonged to PT8. SE isolated from hen house drag swabs from farms that produced SE-positive eggs were also typed as PT8. However, SE isolated from hen paddock environment of one medium flock producing SE-positive eggs was typed as PT23 (Table 2).
PFGE fingerprint patterns (pulsotypes)
The XbaI digested genomic DNA of SE isolated from this study yielded three distinct PFGE pulsotypes. The most common PFGE pattern identified was JEGX01.0004, which was represented by 20 (90%) of the isolates tested. The PFGE patterns of two SE isolates originating from rodents and the hen paddock were different from the rest of the isolates. An SE isolate from the rodent sample (Flock 7) was typed as JEGX01.0021, whereas the SE isolated from the hen paddock swab (Flock 1) was untypeable (Table 2 and Fig. 1). All SE isolates except the SE that originated from the hen paddock swab collected from Flock 1 shared an identical BlnI restriction pattern (Fig. 2).
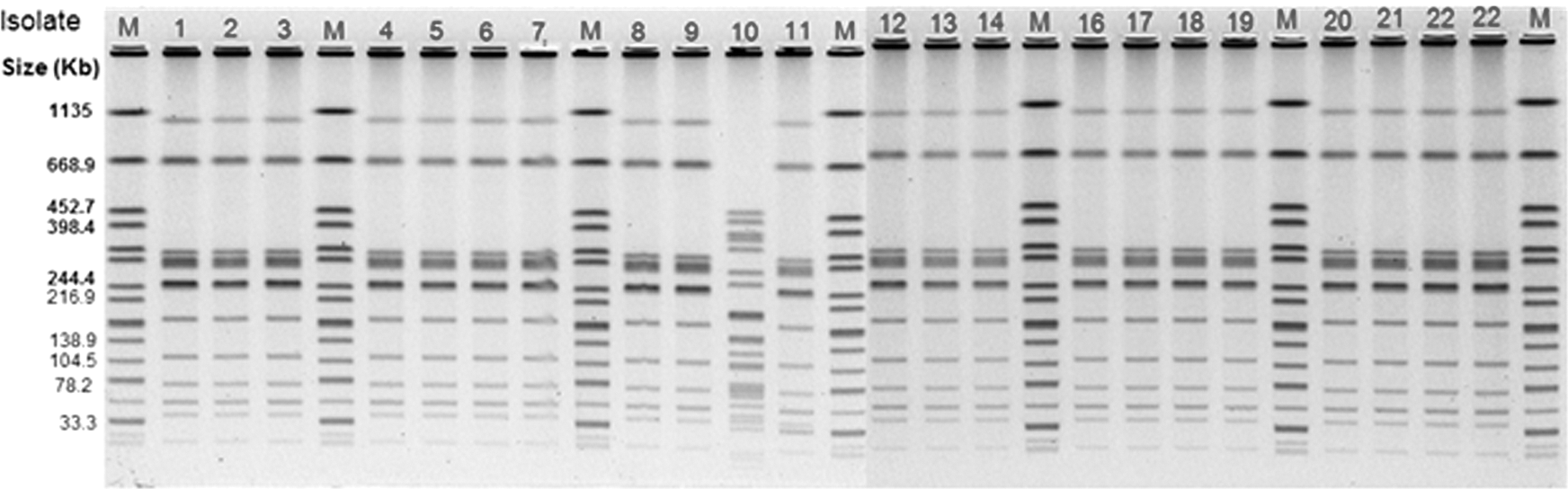
Pulsed-field gel electrophoresis patterns for XbaI-digested genomic DNA of 22 Salmonella Enteritidis strains isolated from hen house swabs, rodents, pooled eggs, and hen paddock swabs from nine contaminated flocks together with XbaI-digested Salmonella Braenderup used as the marker (M).

Pulsed-field gel electrophoresis patterns for BlnI-digested genomic DNA of 22 Salmonella Enteritidis strains isolated from hen house swabs, rodents, pooled eggs, and hen paddock swabs from nine contaminated flocks together with BlnI-digested Salmonella Braenderup used as the M.
CRISPR-MVLST types
The CRISPR-MVLST yielded three ESTs. A majority of isolates (85%) belonged to EST4. One SE isolated from a rodent (Flock 7) and another isolated from a hen house drag swab (Flock 9) were typed as EST8 and EST42, respectively. Another SE isolated from hen house drag swabs (Flock 5) was untypeable by CRISPR-MVLST (Table 2).
Discussion
This study monitored SE contamination of hen house environments and eggs for two consecutive flock cycles in small (<3000 hens) and medium (3000–50,000 hens) layer flocks in Pennsylvania and Iowa. SE was the predominant serovar (40.74%) isolated from poultry environments, rodents, and pooled eggs (Table 1). Although five out of nine SE-positive farms had SE-positive rodents, none of the farms that produced SE-positive eggs had SE-positive rodents. Except for one isolate of SE, all other SE isolated from rodents belonged to PT8 and PFGE pattern JEGX01.0004, indicating the presence of SE-infected rodents as a possible risk factor for SE contamination of layer flocks. Association of other sources for SE contamination of layer flocks could not be established since SE was not isolated from feed, water, and fly samples or other environmental samples examined in the study. Similarly, chick box papers were negative for SE, suggesting that SE-positive flocks have acquired SE through horizontal transmission at the layer farm but not through vertical transmission. As rodents were detected positive for SE, one likely source is the contamination of poultry house environment by SE-infected rodents.
The JEGX01.0004 pattern was the most common PFGE pattern reported to the national PulseNet database (CDC, 2010). It was also the most common PFGE pattern implicated in human foodborne outbreaks of SE in many states, including Pennsylvania and New York. According to a recent study, this PFGE pattern represented more than 99% of SE isolated from chicken carcass rinsates (Sandt et al., 2013). In our study of the 22 SE isolates recovered from poultry house environments, rodents, and eggs, 20 (91%) isolates belonged to the JEGX01.0004 pattern indicating that SE of this PFGE pattern may be enzootic in both commercial broiler and layer facilities posing a major risk to public health. Also, 20 (90%) isolates of SE were typed as PT8, the most common PT associated with human foodborne SE outbreaks in the United States. Out of all SE isolates examined in the present study, 19 (86%) demonstrated both the JEGX01.0004 PFGE pattern and PT8.
In PFGE, the use of the secondary restriction enzyme, BlnI, is recommended if PFGE patterns obtained with the primary enzyme are indistinguishable. However, all isolates except the SE isolate recovered from a single hen paddock swab belonging to PT23 produced an identical banding pattern also after digestion with BlnI, confirming that all 18 are identical based on PFGE and phage typing. The CRISPR-MVLST has emerged as a new typing technique for nontyphoidal Salmonella serovars with a higher discriminatory power than PFGE (Liu et al., 2011a, b; Shariat et al., 2013). In our study, 21 isolates of SE yielded three EST patterns and one isolate was untypeable. Of the 21 isolates that could be typed by CRISPR-MVLST, 19 belonged to EST4, whereas one each was typed as EST8 and EST42. The isolate belonging to EST8, originated from a rodent (Flock 7), was different from the rest of the isolates by all three typing techniques. However, the isolate that belonged to EST42 by CRISPR-MLVST was typed along with the majority of isolates (PT8 and pulsotype JEGX01.0004) by the other two typing methods. It is widely accepted that PFGE, which reflects genetic changes that occurred during a short evolutionary time scale, is more useful in epidemiologic investigations than the typing techniques that reflect events occurring over a long evolutionary time scale and useful for phylogenetic classifications (Riley, 2004).
Based on the typing methods used in the study of the nine flocks that were positive, SE isolated from six flocks were indistinguishable regardless of the sample type and the flock age. In Flock 1, except the SE isolate recovered at 41 weeks of age from the hen paddock swab, which yielded a different PT and untypeable by PFGE XbaI digestion, all other SE recovered from the eggs and hen house swabs at different time points were indistinguishable suggesting a common origin. In Flock 9, SE isolated from a hen house drag swab at 15 weeks of flock age (PT8, JEGX01.0004, and EST42) was different from an SE isolated from a rodent at the same time point (PT8, JEGX01.0004, and EST4) only with the CRISPR-MVLST analysis. It is interesting to note that the difference between EST42 and EST4 lies in fimH but not in any of the CRISPR loci. Altogether, SE isolated from layer house environment and eggs in the study demonstrated a clonal spread of SE belonging to PT8 and pulsotype JEGX01.0004 among these layer facilities (Table 2 and Fig. 1).
SE-infected rodents are considered a major source of SE contamination of layer poultry environment (Henzler et al., 1998; Denagamage et al., 2015). In this study, among various samples types collected from different premises, SE was isolated more frequently from rodents (five premises) and hen house swabs (five premises) followed by eggs (four premises) than any other sample types. Although the layer house environment and rodents from the same flock were not positive for SE, it is likely that rodents act as a reservoir of SE in these flocks but were not positive for SE at the time of sample collection due to the intermittent shedding of SE by infected hens (Henzler et al., 1998, 1999). This is further supported by the observation that SE isolated from the rodents and other environmental sources shared the same PFGE fingerprint pattern (JEGX01.0004) and phage type (PT8).
This study also examined the effectiveness of egg quality assurance practices, such as rodent control, vaccination, hygiene practices, and disinfection procedures to reduce infection levels, and periodic monitoring for Salmonella. The first phase of sampling (first flock cycle) was initiated before these practices were introduced to the flock. However, the flocks with more than 3000 were required to comply with the FDA Final Egg Rule in July 2012. Therefore, some medium-sized flocks have already started EQAP practices before conclusion of the flock cycle 1 sample collection hindering the opportunity to assess the effectiveness of EQAP practices in these flocks. This could be the reason why medium-sized flocks were detected positive at the age of 15 weeks, but not afterward. The medium-sized premises that were SE-positive during the first sample collection period were negative during the subsequent sample collection period and this could be attributed to the EQAP practices implemented by the producers.
In contrast to medium-sized flocks, small flocks (both hen house swabs and eggs) were still positive for SE but there was a significant reduction of frequency of SE isolation in flock cycle 2 than in flock cycle 1, which again highlighted the effectiveness of EQAP practices in minimizing SE contamination of layer flocks. Since there was no requirement for small-sized flocks to comply with the FDA Final Egg Rule, these flocks did not have satisfactory EQAP practices on the farms during the first sample collection period except some flocks received live and/or killed vaccines to control SE. Although SE was not recovered from vaccinated flocks, this study was unable to determine the effect of vaccination on SE shedding by hens and SE contamination of shell eggs due to limited number of unvaccinated flocks enrolled in the study compared to the number of vaccinated flocks. Also, withdrawal of 17 out of 32 producers with less than 3000 birds from the study after the first sample collection period necessitated recruiting new producers for the second phase (second flock cycle) of the study. Due to this loss, we were unable to monitor the same 32 small-sized premises throughout the study.
Although this study was able to make several important observations, it has some limitations. First, the participants selected for the study were based on voluntary participation. In one state, an honorarium had to be provided to participating producers, which also contributed to selection bias. Second, medium-sized flocks with 3000 to 50,000 birds started implementing EQAP practices during the first sampling phase to comply with the FDA Final Egg Rule. Third, due to withdrawal of some producers of small flock category, there was a reduced follow-up during the second phase of the study. Fourth, in some instances, the most suitable samples were not available for testing. This happened, in particular, with the small farm category as they were exempted from the FDA Final Egg Rule. For example, on a few occasions, sample collectors noticed the premises had been cleaned by the producer just before their arrival to collect environmental samples. Finally, the number of eggs tested from each environmental positive flock was based on the flock size where the greater the number of birds in a flock, the lower the percentage of eggs tested, although more eggs. A 1000 egg sample is required by the FDA safety rule for all flocks greater than 30,000 hens even if the flock has more than 300,000 hens. Scientific evidence suggest that only a small number of hens in an infected flock shed SE at any given time and infected hens may lay many uncontaminated eggs (Humphrey, 1994). Despite the 2–100 × fold greater egg sampling rate utilized herein compared to the FDA egg safety rule, this study may have missed the opportunity to detect SE-contaminated eggs produced by some of the SE-infected flocks during the study period. Similarly, the SE-positive eggs might have been detected at a higher frequency in SE-infected small flocks than in medium flocks (∼10 × greater sampling rate or less).
Conclusions
This study demonstrated that the most common SE subtypes associated with human foodborne outbreaks (PT8 and JEGX01.0004 pulsotype) are present in laying hen environments, re-emphasizing the importance of enforcing EQAPs to minimize SE contamination of medium and small layer flocks and their eggs. Isolation of the same subtypes of SE also from the rodents highlights the importance of giving priority to rodent control measures in EQAPs.
Although EQAP practices, such as biosecurity, control of rodents, flies, and other pests, cleaning and disinfection of hen houses in between flocks, vaccination, monitoring hen house environment and eggs for SE, record keeping, and refrigeration of eggs during storage and transport recommended for large flocks can be applied to small flocks, it is imperative to refine the sampling protocol to make it economically viable and realistic to the small producer considering the limited number of birds on small layer farms, in particular, in backyard flocks. Furthermore, financial constraints associated with testing, diverse farm management practices, and lack of knowledge on food safety practices may hinder implementation of EQAPs for small egg producers.
Footnotes
Acknowledgments
This study was funded by the USDA-National Integrated Food Safety Initiative (Grant No. 2011-51110-30993) to S.K. and the United States Army Research Office grant to E.G.D. (W911NF-11-1-0442). The authors thank the poultry producers who volunteered to participate in the study.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.