
Abstract
Antimicrobial drug use in food animals is associated with an elevation in relative abundance of bacteria resistant to the drug among the animal enteric bacteria. Some of these bacteria are potential foodborne pathogens. Evidence suggests that at least in the enteric nontype-specific Escherichia coli, after treatment the resistance abundance reverts to the background pre-treatment levels, without further interventions. We hypothesize that it is possible to define the distribution of the time period after treatment within which resistance to the administered drug, and possibly other drugs in case of coselection, in fecal bacteria of the treated animals returns to the background pre-treatment levels. Furthermore, it is possible that a novel resistance mitigation strategy for microbiological food safety could be developed based on this resistance reversion phenomenon. The strategy would be conceptually similar to existing antimicrobial drug withdrawal periods, which is a well-established and accepted mitigation strategy for avoiding violative drug residues in the edible products from the treated animals. For developing resistance-relevant withdrawals, a mathematical framework can be used to join the necessary pharmacological, microbiological, and animal production components to project the distributions of the post-treatment resistance reversion periods in the production animal populations for major antimicrobial drug classes in use. The framework can also help guide design of empirical studies into the resistance-relevant withdrawal periods and development of mitigation approaches to reduce the treatment-associated elevation of resistance in animal enteric bacteria. We outline this framework, schematically and through exemplar equations, and how its components could be formulated.
Introduction
A
Field data show that the treatment-associated elevation in resistance may be transient, with the resistance abundance reverting to background levels without further interventions. Relevant data in cattle include when the cephalosporin ceftiofur is administered parenterally in therapeutic dosages, the abundance of cephalosporin-resistant fecal bacteria reverts to the pre-treatment levels in ∼5–18 d after the treatment period (Jiang et al., 2006; Lowrance et al., 2007; Singer et al., 2008). Other studies report a more transient (Boyer and Singer, 2012) or no resistance elevation (Mann et al., 2011) during ceftiofur treatments. The temporality of resistance elevation is also reported for chlortetracycline treatments in cattle (Platt et al., 2008). In cattle pens sampled biweekly through the feedlot period, no association can be detected between pen-level exposure to the fluoroquinolone enrofloxacin or florfenicol and prevalence of resistance to those drug classes in the fecal bacteria (cattle were treated in hospital pens and returned to the sampled pens most often in 3 d post-treatment) (Morley et al., 2011). No association can be detected between treatments by ceftiofur, the macrolide tilmicosin, or oxytetracycline early in the feedlot period and corresponding resistances in the cattle fecal bacteria at slaughter (Checkley et al., 2008). These studies used nontype-specific Escherichia coli as an indicator fecal bacteria, because of their ubiquity, to study the resistance dynamics. When there is a coselection for resistance to other antimicrobial classes, the multiresistance in the cattle fecal E. coli appears to follow the temporal reversion dynamics of resistance to the administered drug (Lowrance et al., 2007). Overall, we termed this phenomenon “post-treatment resistance reversion.”
We hypothesize that a novel mitigation strategy for microbiological food safety can be developed based on the post-treatment resistance reversion phenomenon, similar to existing antimicrobial withdrawal period for avoiding violative drug residues in edible animal products. It should be possible to define the distribution of the time period after an antimicrobial treatment in food animals within which resistance in the animal fecal bacteria to the administered drug, and possibly other drugs in case of coselection, will return to the background pre-treatment levels. Since resistance cannot be eliminated from enteric bacteria, this strategy would facilitate the best practically achievable level of microbiological food safety with respect to resistance for animals that required antimicrobial treatments.
Proposed Approach
The post-treatment resistance reversion period can be measured in empirical studies for each antimicrobial and its specific application. However, having a modeling framework for projecting the period distribution offers three advantages. First, the models, once developed, could guide the empirical study design, by suggesting time points and influential variables to measure, helping to reduce and improve animal use. Second, the models parameterized for the antimicrobial for one of the animal's categories could be adaptable to project the expected intestinal antimicrobial concentrations and impact in other categories if the drug is repurposed for other applications. Third, relative contributions of the component processes (e.g., drug pharmacology and bacterial ecology) and their specific variables to resistance could be estimated, providing ideas for mitigating the duration and extent of treatment-associated resistance elevation.
A possible mathematical framework joining necessary pharmacological, microbiological, and animal production components to project the distribution of the post-treatment resistance reversion period in the animals' enteric bacteria is outlined hereunder. The framework links (sub-) models of the antimicrobial drug pharmacokinetics related to its intestinal concentrations (PK-for-intestine), drug pharmacodynamics (PD) against enteric bacteria, bacterial ingestion by the animals and intestinal bacterial ecology, and resistance dynamics (e.g., mutations and resistance transmission) in the enteric bacteria. The framework is presented in a generalized form, including the relevant processes across antimicrobial drug classes and enteric bacteria. For a given antimicrobial or bacteria with given resistance transmission dynamics, the necessary parameters will need to be selected from the framework.
Pharmacokinetics
Exposure of animal enteric bacteria to antimicrobial drugs or their antimicrobially active metabolites occurs within the animal intestine where these substances are excreted or passed with digesta. There is a variation in the metabolism and excretion of different antimicrobials, and in processes these substances undergo in the intestinal environment (e.g., degradation). The drug/metabolite intestinal concentrations do not scale linearly with the treatment schedule, dose and duration, at the animal level. This may explain inconsistency in results of analyses of the impact of antimicrobial use on resistance in enteric bacteria operating with the use estimates of different antimicrobials at the animal or farm level (Bager et al., 1997; Kruse et al., 1999; Harada et al., 2008; Varga et al., 2009a, 2009b; Deckert et al., 2010; Garcia-Migura et al., 2014; Benedict et al., 2015; Noyes et al., 2016). The selective pressure on the treated animals' enteric bacteria can be estimated using a new type of the PK-for-intestine models that project concentrations of the drug and its antimicrobially active metabolites in the intestine during the treatment schedule and the antimicrobial's intestinal elimination period. Such models have been developed for cephalosporin ceftiofur used parenterally (Volkova et al., 2012) and chlortetracycline used per os (Cazer et al., 2014) in cattle. Component processes of PK-for-intestine include passage of the antimicrobial with digesta, biliary excretion, absorption to central circulation, presence of enterohepatic circulation, whether the intestinal drug/metabolites are antimicrobially active, and, if so, their degradation and binding to the digesta. Schematics of these models in Figure 1 illustrate the body compartments and drug/metabolite flows; Supplementary Data provide exemplar generalized equations for a PK-for-intestine model (Supplementary Data are available online at
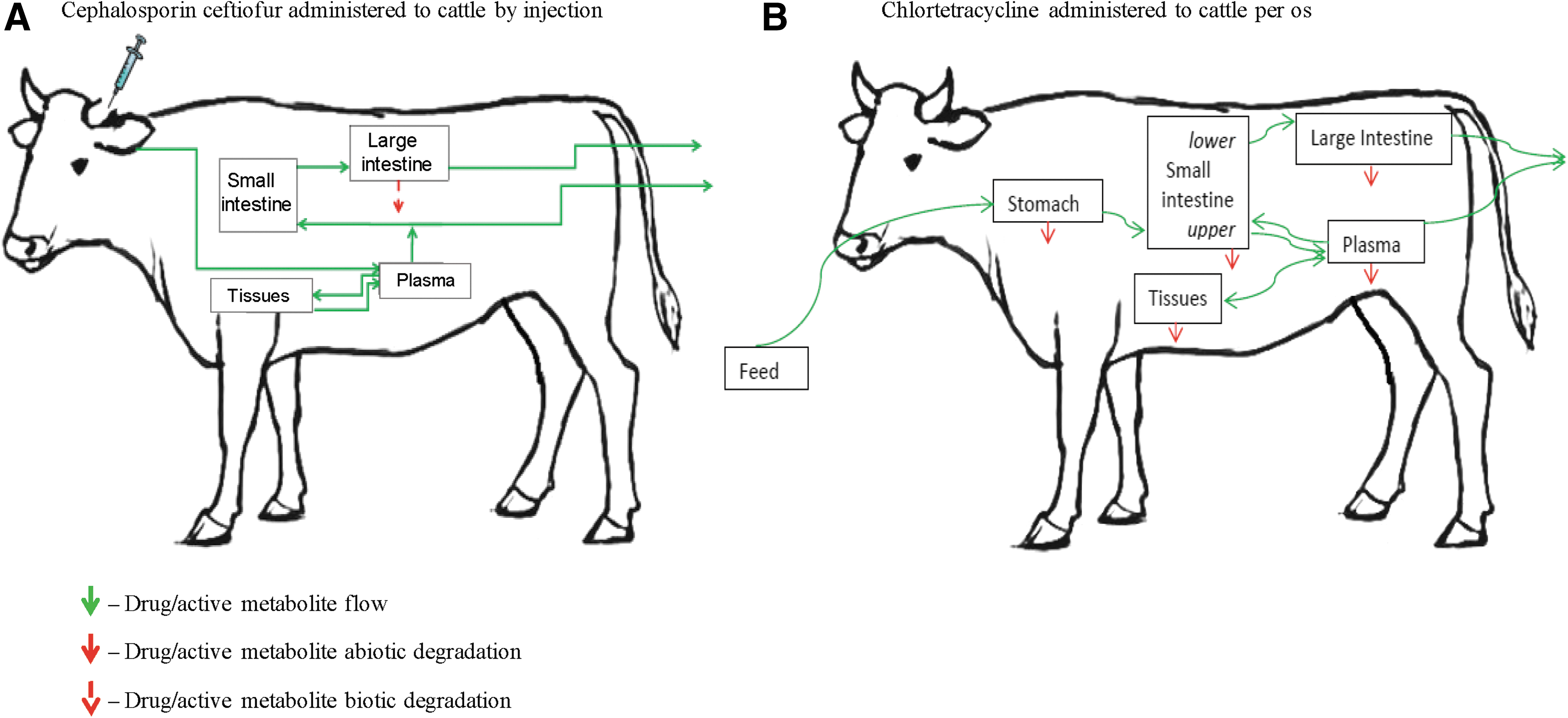
Schematics of two PK-for-intestine models for projecting concentrations of antimicrobial drugs and their active metabolites in the treated animals' intestines (Volkova et al., 2012; Cazer et al., 2014).
Allometric relationships between the animal's body size and the fraction of the administered antimicrobial drug occurring in the intestine may not exist among food animal species. This is because, first, the rates of antimicrobial drug absorption from the intestine, metabolism, biliary excretion, and enterohepatic circulation differ among food animal species (Toutain et al., 2010; Huang et al., 2015). Second, those species differ in gastrointestinal anatomy, physiology, and microbiome that determine further processes antimicrobials undergo in the intestinal environment. For example, food animal species differ in the intestinal transit time, which influences the degree of antimicrobial's abiotic or biotic degradation in the intestine. The microbiome's taxonomic composition may differ among food animal species (Shanks et al., 2011; Isaacson and Kim, 2012; Kim and Isaacson, 2015). Genera and species of enteric bacteria vary in their production of β-lactamases, enzymes degrading β-lactam antimicrobials (Welling et al., 1992; Livermore, 1995; Jones et al., 1997; Rafii et al., 2009; Wagner et al., 2011). Hence, a microbiome composition difference may imply a different intensity of the intestinal biodegradation of β-lactam antimicrobials, unless the same intensity is achieved by variable microbiome compositions in different animal species. Such differences would not be captured by an allometric scaling of the PK-for-intestine projections based on the animals' body sizes. Hence, the PK-for-intestine models would need to be fitted for each species individually.
Pharmacodynamics
Projecting the antimicrobial effects of the drug/metabolite concentrations occurring in the animal intestine against the enteric bacteria requires new PD models that capture the effects against these dense bacterial populations composed of subpopulations with a variable degree of susceptibility to antimicrobials. The models should reflect the functional dependencies of the antimicrobial effects on the bacterial density (Udekwu et al., 2009) and susceptibility (Gehring and Riviere, 2013; Ahmad et al., 2015), type of action expected given the drug/metabolite and their concentrations occurring (bactericidal or bacteriostatic), connection to the intestinal bacterial population growth (Volkova et al., 2012), and prominence of postantibiotic effect (Hanberger, 1992; Cars and Odenholt-Tornqvist, 1993). Supplementary Data provide an exemplar generalized equation for such a PD model. Consideration should be given to the impact of pH and anaerobic conditions of the intestine on the drug/metabolite antimicrobial activity and the bacterial susceptibility (Bryan et al., 1979; Schlessinger, 1988; Cid et al., 1994; DeMars et al., 2016). Furthermore, there may be feedbacks in the effects, for example, the enzymatic antimicrobial resistance may increase with the bacterial exposure to higher concentrations of some antimicrobials (Livermore, 1987; Jacobs et al., 1997; Zeng and Lin, 2013), unless the enzyme-producing species are decimated by the antimicrobial (Welling et al., 1991), or refractory phenotypic bacterial resistance (not genetically encoded) may manifest over the time of antimicrobial exposure (Levin et al., 2014).
Dynamics of bacterial ecology and resistance in the intestine
Bacteria in the intestine exist as either luminal or adhered to the endothelium and mucosa. The data suggest that adhering E. coli types are a smaller fraction of the total fecal E. coli (Gordon, 2013). Thus, the model presented focuses on the luminal E. coli. The luminal enteric bacterial ecology includes the bacterial movement into the lumen with digesta, through and out due to peristalsis and defecation, and population growth in the lumen. Bacterial functions that act as the mechanisms of antimicrobial resistance differ depending on the bacterial species and antimicrobial (Walsh, 2000; Beceiro et al., 2013). The genes encoding those functions spread in bacteria through several processes: vertically through replication (a.k.a. clonal expansion), horizontally through conjugative transfer of plasmids and other extrachromosomal genetic elements, and from dead to alive bacteria through transformation or transduction by bacteriophages (Boerlin and Reid-Smith, 2008). For modeling purposes, the resistance transmission dynamics are commonly categorized into the vertical vs. horizontal (D'Agata et al., 2008; Gehring et al., 2010; Andersson and Hughes, 2011; Tazzyman and Bonhoeffer, 2014). Schematic and formulas of the model of ecology of a bacterial species and its antimicrobial resistance in the intestinal lumen are shown in Figure 2; the model's exemplar generalized equations are provided in Supplementary Data.
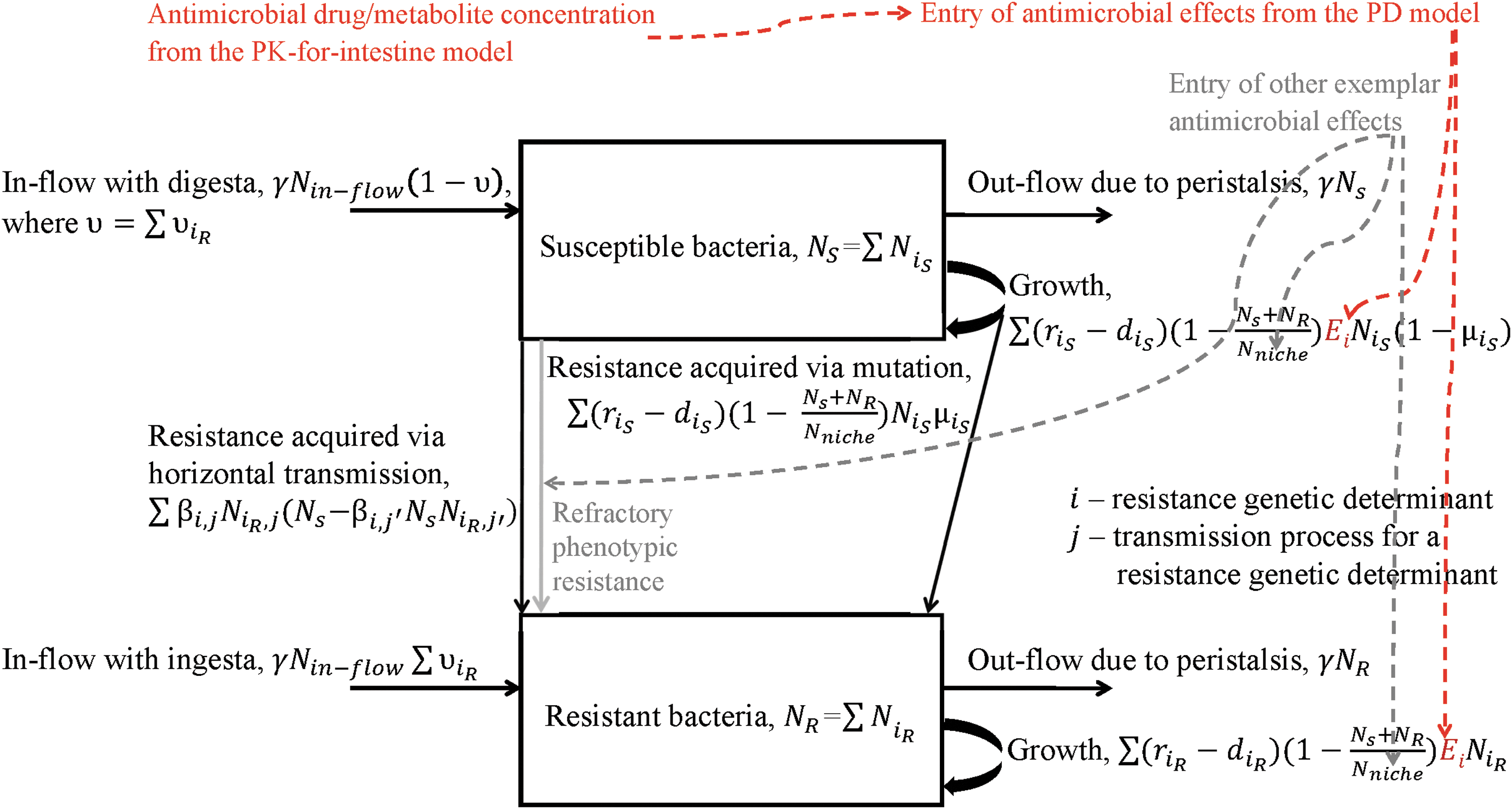
Schematic of the model of ecology of a bacterial species and its antimicrobial resistance dynamics in the intestinal lumen, and joining of the bacterial ecology and PK-PD models. Black: Bacterial ecology and resistance transmission dynamics in the bacteria. Red: Joining PK-PD models to the bacterial ecology. Gray: Adding other effects of the antimicrobial on the bacterial species. PK, pharmacokinetics; PD, pharmacodynamics.
Framework application: a simple example
An earlier model of cephalosporin resistance dynamics in fecal E. coli of cattle during and after parenteral treatment with ceftiofur includes several elements of the proposed framework (Volkova et al., 2012). The system modeled was simpler than for other antimicrobials: the resistance transmission was primarily clonal and by plasmids; the distribution of E. coli ceftiofur susceptibility could be assumed to be dichotomous with a large fraction of highly susceptible and a smaller fraction of resistant subpopulations; and possible feedbacks between the antimicrobial concentration and bacterial population resistance in the intestine were not included. The model's temporal projections of the resistant E. coli relative abundance agree well with field data for different drug formulations and treatment schedules in beef and dairy cattle (Jiang et al., 2006; Lowrance et al., 2007; Singer et al., 2008; Volkova et al., 2012). We reanalyzed the simulations to illustrate a possible distribution of the resistance reversion period in 100 beef cattle after ceftiofur treatment against bovine respiratory disease (BRD). We defined the outcome in the animal as the period after the 10-d treatment (when antimicrobially active ceftiofur metabolites are present in the animal's central circulation) until the relative abundance of cephalosporin-resistant fecal E. coli reverts within 10% of the average background pre-treatment level of 100 animals. The analysis demonstrated that in 70% of the animals, this outcome may occur in 19 d post-treatment.
Production practices
The animal production characteristics and practices that significantly influence the active concentrations of antimicrobial drug/metabolite in the animal intestines can be identified by the population PK methods (Mason et al., 2012; Li et al., 2014, 2015); a tentative factor list is given in Table 1. The influential animal characteristics likely include age, category that determines the animal size and body composition (e.g., finishers vs. breeders), and production (e.g., lactation) and health (e.g., mastitis) status. These affect the absorption, metabolism, and excretion of antimicrobial (Brown et al., 1996; Rule et al., 1996; Green and Duffull, 2004; Kissell et al., 2015; Gorden et al., 2016). The influential production practices likely include the feed composition, roughage, intake, and feeding frequency. These affect the intestinal transit, binding, and degradation of antimicrobial (Shaver et al., 1986; Zebeli et al., 2007; Shanks et al., 2011; Kim and Isaacson, 2015). The production practices also affect the abundance of total and resistant bacteria in feed, water, and other matters animals ingest. These enter the framework as the bacterial and resistance inflows in the enteric bacterial ecology and resistance model (Fig. 2). The inflows' definitions provide means to link the within-animal resistance dynamics to epidemiology of resistant bacteria in the animal production systems.
Impact of disease pathophysiology
Pathophysiology of the treated and concurrent disease can affect the PK-for-intestine. There can be changes in the drug/metabolite concentration in the central circulation and hence its biliary excretion (Sarwari and Mackowiak, 1996; Marier et al., 2001). Intestinal motility and thus the drug/metabolite intestinal transit time can change due to the inflammatory responses (Winter et al., 2010). Decline in appetite resulting in a reduced feed intake, for example, with BRD (Galyean and Hubbert, 1995), increased water consumption, and changes in enteric microbiome composition in disease (Kostic et al., 2013; Kim and Isaacson, 2015) may affect the intestinal transit, binding, and degradation of antimicrobial.
Accounting for variability
Variability in resistance reversion period in food animal populations can arise from contributions of a variety of factors. Some factors are better understood; their expected variability can be specified explicitly, for example, for the PK parameters by the population PK methods (Martin-Jimenez and Riviere, 1998; Buur et al., 2006; Mason et al., 2012; Wu et al., 2013). Others are less understood, but can be assumed to vary stochastically over time, for example, background resistance levels in enteric bacteria (Wagner et al., 2003) or resistance horizontal transmission rate among the bacteria (Suchard, 2005; Minoia et al., 2008). Elaborating the modeling framework into a stochastic form could provide the means, once parameterized for a given treatment scenario, to incorporate contributions of both factor types, predictable and stochastic, to simulate the total possible distribution of the reversion period among treated animals [see for a simplified example Volkova et al. (2013b)]. Values of factors most difficult to define could be estimated through dynamic optimization (Gelb, 1974; Anderson and Moore, 1979) between the model and data on the resistance dynamics during the treatment and the antimicrobial intestinal elimination period.
Once fully elaborated for a treatment scenario, the framework in a stochastic form reflecting total variation in the reversion period can be used to investigate which individual processes or factors, across the submodels, make the largest relative contribution to the reversion period length in the production animal populations. This could be done through the outcome's global sensitivity analysis (Mckay et al., 2000; Marino et al., 2008), which is conceptually similar to partitioning the variation to the contributions of individual factors. The contributions' strength and significance could be evaluated using statistical methods (Lanzas et al., 2008; Volkova et al., 2013a). For some of the factors, further identifying the main level of their variability may provide insights into intervention design; for example, whether the intestinal microbiome composition and biotic degradation of antimicrobial varies primarily at the animal species, animal-group feeding, or individual level (Welling et al., 1992; Shanks et al., 2011; Isaacson and Kim, 2012; Hanning and Diaz-Sanchez, 2015; Kim and Isaacson, 2015).
Discussion
We hypothesize that elevation of antimicrobial resistance in enteric bacteria of food animals during antimicrobial treatment is temporal; after treatment the resistance abundance may revert to background levels without further interventions. A mitigation strategy for microbiological food safety can be developed based on this phenomenon. The supportive evidence comes from field studies with antimicrobial treatments for animal disease, with comparably high doses of drugs administered for short periods. Antimicrobial schedules for other purposes are designed differently, for example, for animal growth promotion, low antimicrobial doses are given for extended periods, and may induce less reversible changes in the enteric bacteria (Danzeisen et al., 2011; Shanks et al., 2011; Kim et al., 2012; Looft et al., 2012). Antimicrobial use for animal disease metaphylaxis in certain cases lies in-between, with a lower dose administered for a longer period compared with the treatment (e.g., chlortetracycline per os at 22 mg/kg body weight for 5 d for BRD treatment vs. at 350 mg/head for 28 d for BRD metaphylaxis). In other cases, the metaphylactic and treatment schedules are similar (e.g., florfenicol parenterally at 40 mg/kg for BRD control or treatment). When the treatment and metaphylactic schedules differ, the resistance reversion period may differ correspondingly. Our hypothesis focuses on the antimicrobial resistance abundance and does not include other possible effects of antimicrobials on enteric bacteria.
Evidence for the resistance reversion comes from studies with nontype-specific fecal E. coli. These E. coli shared by human and nonhuman animals are considered to be a substantial reservoir of genetic determinants of resistance (Hammerum and Heuer, 2009). The presence of the reversion phenomenon in foodborne pathogens of other taxonomies, for example, fecal vibrio bacteria, needs to be investigated. Furthermore, the overall enteric resistome dynamics may differ from specific resistance dynamics in individual component bacterial species.
Acceptance of the hypothesized mitigation strategy based on resistance reversion by veterinarians and producers may be facilitated by its conceptual and practical similarity to existing drug withdrawal for toxicological food safety (avoiding violative antimicrobial residues in animal products), especially for therapeutic antimicrobial use. In the long run, this new mitigation strategy may support a more harmonized public view of antimicrobial use for animal health.
There is evidence of an increased use of antimicrobial drugs in food animals in an extra-label manner since older label dosages may be ineffective; the necessary extended withdrawal is estimated using PK modeling (Li et al., 2014). The withdrawal period for violative residue prevention under per- or extra-label use may be compared with the resistance reversion period and the longer interval applied, harmonizing mitigation of the toxicological and microbiological risks. This could be implemented through the Food Animal Residue Avoidance and Depletion program (
Conclusions
The mitigation strategy based on the post-treatment resistance reversion phenomenon could facilitate the best practically achievable level of microbiological food safety with respect to antimicrobial resistance when antimicrobial treatment is necessary for animal disease.
Footnotes
Acknowledgment
This work was funded by Kansas Bioscience Authority.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.