
Abstract
The increase in prevalence of Salmonella 1,4,[5],12:i:- related infections over the last few years has been considered a public health issue in many European countries, especially as this serovar may be associated with tetraresistance to ampicillin, streptomycin, sulfonamides, and tetracyclines (R-type ASSuT). Salmonella 1,4,[5],12:i:- isolates (n = 187) obtained by the Portuguese National Laboratory from different sources, including human clinical cases (n = 170), veterinary (n = 10), environmental (n = 6), and food samples (n = 1), were collected from 15 districts between 2006 and 2011. All isolates were serotyped using the slide agglutination method and results were confirmed by multiplex PCR for the monophasic variant. From the confirmed Salmonella 1,4,[5],12:i:-, R-type ASSuT isolates were selected by disc diffusion and minimal inhibitory concentration (MIC) determination for further characterization by pulsed-field gel electrophoresis restriction with XbaI, virulence genes determination by PCR, additional antimicrobial resistance profiling by disc diffusion, and epidemiological distribution evaluation. Out of the 187 serotyped isolates, 133 were confirmed as Salmonella 1,4,[5],12:i:- with a R-type ASSuT occurrence of 61.7%. Distribution among Portuguese districts showed a higher percentage of reported cases in coastal areas, in particular, in Porto (24.8%), Setúbal (13.5%), and Aveiro (12.8%), probably due to the higher population density. Clonality analysis revealed a high diversity of pulsotypes with the majority of human salmonellosis cases being attributed to sporadic events. All isolates harbored 14 out of the 18 virulence genes evaluated and 87.8% of the isolates showed all the resistance genes frequently associated with the European clone, blaTEM+ sul2+ straA-straB+ tetB+. This study shows that Salmonella 1,4,[5],12:i:- resistant isolates are widely distributed in Portugal. This may be related to a selective advantage offered by R-type ASSuT profile, the presence of multiple virulent features, including the ability to form biofilms, which along with a high diversity of pulsotypes may be responsible for the dissemination through the country.
Introduction
S
Genes responsible for this resistance phenotype are present in a chromosomal resistance island that frequently includes the blaTEM, strA-strB, sul2, and tetB genes, with some isolates having additional resistances (Lucarelli et al., 2010). Also, the presence of multiple virulent determinants, along with biofilm formation, enables Salmonella to infect several host species and persist in the environment (Steenackers et al., 2012).
The aim of the present study was to characterize, in terms of distribution, molecular typing, antimicrobial resistance, and virulence factors, Salmonella 1,4,[5],12:i:- R-type ASSuT isolates obtained in Portugal from different sources, mainly of human origin and also from animal and environment. This study provides useful information for understanding the dissemination and epidemiology of this serovar in Portugal and within Europe, contributing to the future establishment of preventive and therapeutic strategies, as well as suitable regulatory measures regarding public health safety.
Materials and Methods
Bacterial isolates
A total of 187 Salmonella 1,4,[5],12:i:- isolates were included in the study. The samples were collected in Portugal between 2006 and the first quarter of 2011 from different sources, including human clinical cases (n = 170), veterinary (n = 10), environmental (n = 6), and food samples (n = 1).
Human salmonellosis is a mandatory notifiable disease in Portugal, respecting a passive surveillance system, and the isolates represent all reported cases of Salmonella 1,4,[5],12:i:- in Portugal during this period of time. All Salmonella were isolated at the Portuguese National Laboratory, the National Health Institute Doutor Ricardo Jorge, and serotyped using the slide agglutination method according to the Kauffmann–White scheme (Grimont and Weill, 2007). Identification of serotyped isolates was confirmed by multiplex PCR (mPCR), and confirmed Salmonella 1,4,[5],12:i:- isolates were selected for detection of R-type ASSuT. Salmonella 1,4,[5],12:i:- presenting the tetraresistance pattern were further submitted to phenotypic and molecular characterization.
Regarding the human samples, data, including year of sample collection, gender, age, and district location, were recorded.
DNA extraction and mPCR identification of Salmonella 1,4,[5],12:i:-
Salmonella isolates were plated onto Columbia agar with 5% sheep blood (BioMérieux) and incubated at 37°C for 24 h. DNA was extracted from a single colony grown overnight using the guanidine thiocyanate method (Pitcher et al., 1989). Identification of Salmonella 1,4,[5],12:i:- isolates was confirmed using mPCR (Tennant et al., 2010) as recommended by the European Food Safety Authority (EFSA, 2010). A total volume of 25 μL containing 2.5 U of Supreme NZYTaq 2 × Green Master Mix (NZYTech, Portugal), 0.4 μM of primers sense-59 and antisense-83, which amplify the phase 2 (fljB) flagellar gene, 0.8 μM of FFLIB and RFLIA, amplifying the fliB-fliA intergenic region (Stabvida, Portugal), and 3 μL of DNA template were submitted to the following conditions: initial denaturation at 95°C for 2 min, 25 cycles of denaturation at 95°C for 30 s, annealing at 64°C for 30 s, extension at 72°C for 1.5 min, and a final delay at 72°C for 5 min. Amplified products were resolved in a 1.5% agarose gel (Bioline, UK) stained with GelRed (Biotium) and visualized by transillumination under UV (Pharmacia Biotech, Thermal Imaging System FTI-500). NZYDNA ladder VI (NZYTech, Portugal) was used as molecular weight marker. Salmonella enterica subsp. enterica CECT 7162 and CECT 722 were used as monophasic and biphasic Salmonella controls, respectively, for PCR amplification.
Detection of R-type ASSuT pattern
Antimicrobial susceptibility testing
Salmonella 1,4,[5],12:i:- isolates, the identification of which was confirmed by mPCR, were evaluated to detect the R-type ASSuT pattern. Antimicrobial susceptibility testing (AST) was performed by disc diffusion method according to the Clinical and Laboratory Standards Institute (CLSI, 2012) guidelines using ampicillin (AMP, 30 μg), streptomycin (S, 25 μg), sulfamethoxazole (SXT, 10 μg), and tetracycline (TE, 30 μg) and confirmed by determination of minimal inhibitory concentration (MIC) using E-test (BioMérieux) as recommended by the WHO Global Salm-Surv (2003). Test performance was monitored using Escherichia coli ATCC 25922 and executed in duplicate.
Characterization of Salmonella 1,4,[5],12:i:- R-type ASSuT isolates
Evaluation of additional resistances
AST was performed by disc diffusion method according to CLSI (2012) guidelines using amoxicillin/clavulanic acid (AMC, 30 μg), cefotaxime (CTX, 30 μg), ceftazidime (CAZ, 30 μg), ciprofloxacin (CIP, 5 μg), chloramphenicol (C, 30 μg), gentamicin (CN, 10 μg), and nalidixic acid (NAC, 30 μg). Test performance was monitored using E. coli ATCC 25922 and executed in duplicate.
Detection of antimicrobial resistance determinants
Genes coding for resistance to amoxicillin (blaTEM), streptomycin (strA-strB), sulfamethoxazole (sul2), and tetracycline (tetB) were evaluated by PCR using primers and conditions previously described (Lucarelli et al., 2012). Salmonella Typhimurium monophasic variant 105/07/03 gently provided by Dr. Lucarelli's laboratory was used as a positive control for all PCR amplifications.
Genotyping of Salmonella
Genomic fingerprinting of Salmonella was achieved by pulsed-field gel electrophoresis (PFGE) using a CHEF-DRIII apparatus (Bio-Rad Laboratories, San Diego, CA) following DNA digestion with XbaI (Invitrogen) according to the PulseNet protocol (Ribot et al., 2006). BioNumerics software, version 7.5 (Applied Maths, Kortrijk, Belgium), was used to register macrorestriction patterns, clustering analysis was performed using DICE similarity coefficient, and the unweighted-pair group method with arithmetic mean with optimization and position tolerance set at 0.8% and 0.8%, respectively. Salmonella Braenderup strain H9812 was used as molecular weight marker.
Detection of virulence-associated genes
Genes coding for virulence factors such as virulence plasmid (spvC), invasion (invA, invH, sopB), enterotoxin (stn), genes associated with survival within macrophages (phoP, phoQ, slyA) and with the formation of fimbriae (agfA, sefA, safC, pefA) were evaluated by PCR using primers previously described (Huehn et al., 2009). The Quorum-sensing gene (sdiA) was also evaluated by PCR using the primers described (Halatsi et al., 2006). The presence of genes associated with adhesion to Peyer's patches (gipA, lpfD) was also assessed (Borriello et al., 2012). Genes associated with biofilm formation (adrA, csgD, gcpA) were included in the study as previous published (Seixas et al., 2014). Reproducibility was evaluated by selecting 10% replicates using Research Randomizer (Urbaniak and Plous, 2013). Salmonella enterica subsp. enterica CECT 443 and CECT 722 were used as positive controls for PCR amplifications.
Data analysis
Descriptive statistics (reported as proportions) were described. Incidence rates by district and by year were estimated dividing the number of reported cases by the district population or by the country population, respectively, according Census of 2011, conducted by the National Statistical Institute (Instituto Nacional de Estatística, 2011) and multiplied by 100,000.
Results
Out of the 187 serotyped isolates, 133 (71.1%) were confirmed by mPCR as monophasic. Confirmed Salmonella 1,4,[5],12:i:- isolates (n = 133) were obtained from human clinical cases (n = 125), environmental (n = 5) and veterinary (n = 3) samples (Table 1). The male/female ratio of the human clinical samples was 1.33 (69 male and 52 female). It was not possible to register the patient's gender for four samples, and for nine it was not possible to register their age. Distribution included 14 of the 15 districts reported. Three samples were from unknown locations.
Data include isolates, the total number of samples identified by serotyping (S), and confirmed isolates by mPCR (P). Fluids include blood, urine, and peritoneal fluid.
Salmonella 1,4,[5],12:i:- isolate (n = 133) distribution through Portugal, evaluated by district, showed that Porto had the higher number of cases (24.8%), followed by Setúbal (13.5%) and Aveiro (12.8%) (Fig. 1). The incidence rates per 100,000 inhabitants were 0.09 in 2006, 0.14 in 2007, 0.27 in 2008, 0.38 in 2009, 0.66 in 2010, and 0.23 in the first quarter of 2011.
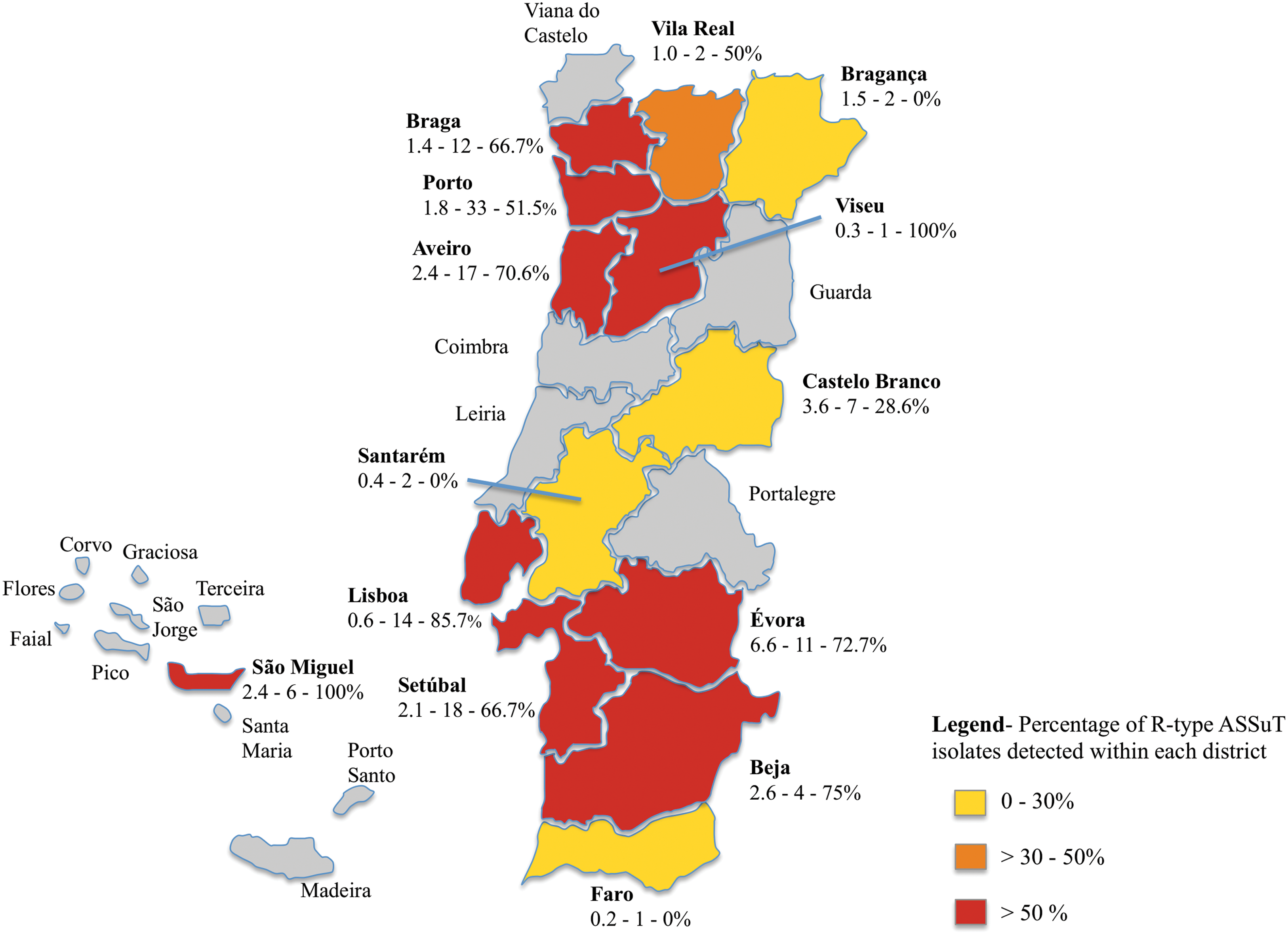
Schematic illustration of Portugal representing the distribution of Salmonella 1,4,[5],12:i:- R-type ASSuT by district from 2006 to the first quarter of 2011. Each district includes the following information: Number of Salmonella 1,4,[5],12:i:- confirmed isolates per 100,000 inhabitants of the corresponding district/total number of reported isolates/percentage of R-type ASSuT isolates detected within each district. For example, Lisboa presents 0.6 confirmed isolates per 100,000 inhabitants—a total of 14 reported isolates—85.7% of R-type ASSuT isolates detected. Color images available online at
Salmonella 1,4,[5],12:i:- isolates (n = 133) revealed an ASSuT profile occurrence of 61.6% (n = 82). The percentage of districts presenting R-type ASSuT isolates was 78.6%.
Forty out of the 82 isolates confirmed as R-type ASSuT presented additional resistances to at least one antimicrobial agent (48.8%), but none was resistant to all seven antimicrobials tested. The highest frequencies of additional resistances were observed for amoxicillin (28%, n = 23), followed by chloramphenicol (15.9%, n = 13), gentamicin (9.8%, n = 8), nalidixic acid (8.5%, n = 7), ceftazidime (4.9%, n = 4), cefotaxime (4.9%, n = 4), and ciprofloxacin (1.2%, n = 1).
Regarding the resistance profiles of the 82 Salmonella 1,4,[5],12:i:- isolates, 18 multiresistant profiles were observed. ASSuT (n = 42), AAMCSSuT (n = 16), and ACSSuT profiles (n = 4) were the three most prevalent profiles. All of the 82 ASSuT isolates showed the highest MIC for ampicillin (>256 μg/mL) and sulfamethoxazole (>1024 μg/mL). The MIC50 value for streptomycin was 512 μg/mL and the MIC90 value was 1024 μg/mL. Tetracycline MIC50 and MIC90 values were 64 and 96 μg/mL, respectively.
Almost all the isolates presented antimicrobial resistance genes, including blaTEM (93.9%), sul2 (95.1%), straA-straB (95.1%), and tetB (93.9%). The most common antimicrobial resistance gene profile was the presence of four determinants blaTEM+ sul2+ straA-straB+ tetB+ in 72 isolates (87.8%). Other resistance gene profiles detected were blaTEM+ sul2+ straA-straB+ in three isolates and tetB+ in a further three isolates (3.7%), followed by single isolates with one of the following profiles blaTEM+ sul2+ tetB+, blaTEM+ straA-straB+, sul2+ straA-straB+ tetB+, and sul2+ straA-straB+ (1.2%).
PFGE typing identified 48 different PFGE profiles among the 82 isolates examined (Fig. 2). Three predominant clones (STYM18, n = 7; STYM23, n = 7; STYM33, n = 6) were identified and correspond to 24.4% of Salmonella 1,4,[5],12:i:- R-type ASSuT isolates. Cluster analysis allowed grouping of the isolates into seven clusters at ∼80% similarity, with four clusters including only one isolate.

Dendrogram based on XbaI-PFGE patterns of the Portuguese Salmonella 1,4,[5],12:i:- R-type ASSuT isolates. AMC, amoxicillin/clavulanic acid; AMP, ampicillin; C, chloramphenicol; CAZ, ceftazidime; CIP, ciprofloxacin; CN, gentamicin; CTX, cefotaxime; NAC, nalidixic acid; NA, not applicable; NR, not registered; PFGE, pulsed-field gel electrophoresis; S, streptomycin; SXT, sulfamethoxazole; TE, tetracycline.
Virulence gene detection revealed that phoP, phoQ invA, invH, sopB stn, slyA, agfA, safC, sdiA, gipA, ipfD, adrA, and csgD genes were present in 100% (82/82) of the isolates. The remaining virulence genes had different frequencies. However, independent of the sample's year, district, and source of isolation, gcpA and spvC genes were present in 95.1% (78/82) and 67.1% (55/82), respectively. sefA and pefA genes were not present in any isolate (0/82). Overall, four different virulence profiles were identified (gcpA+ spvC−, gcpA− spvC+, gcpA+ spvC+, gcpA− spvC−), with all isolates showing at least 14 virulence-associated genes out of the 18 genes tested (Table 2).
AMC, amoxicillin/clavulanic acid; C, chloramphenicol; CAZ, ceftazidime; CIP, ciprofloxacin; CN, gentamicin; CTX, cefotaxime; NAC, nalidixic acid.
Discussion
A rapid increase in Salmonella 1,4,5,[12]:i:- was observed on an international scale, becoming the third most common serovar in humans in 2013 in the European Union (EFSA and ECDC, 2015). Accurate identification of serovar 1,4,5,[12]:i:- is important, since misclassification could result in unnecessary regulatory actions. In fact, serotyping alone can lead to misidentifications and should be combined with mPCR, as recommend by EFSA for identification of Salmonella Typhimurium and 1,4,[5],12:i:-, two serovars included in European regulations aiming the reduction of Salmonella prevalence in poultry (EFSA, 2010).
According to our results, 71.1% of isolates serotyped as 1,4,5,[12]:i:- were confirmed as monophasic variants by PCR assays. Several studies reported similar or higher percentages: 71.4% (Tavechio et al., 2004), 82% (Mandilara et al., 2013), 90.3% (Barco et al., 2011), 91.7% (Wasyl and Hoszowski, 2012), 94.2% (Lailler et al., 2013), and 95.1% (Hopkins et al., 2010). Almost all Salmonella 1,4,[5],12:i:- isolates (28.3%) that were not confirmed as monophasic by mPCR, represent isolates in which the second flagellar phase was not detected by serotyping but was detected by PCR. This can be explained by deletions in the fljB gene that do not compromise the primer binding sites but impair the phase inversion, leading to misidentifications in serotyping (Barco et al., 2011). One remaining isolate (0.6%) did not produce the 1 kb amplicon, which could be attributed to the complete loss of the IS200 fragment (Barco et al., 2011).
The rapid dissemination of this serovar is allied with an escalating incidence of antimicrobial resistance in 1,4,[5],12:i:- strains (Antunes et al., 2011). The ASSuT tetraresistance pattern, already identified in many European countries (Lucarelli et al., 2010), was detected in 61.6% of the isolates included in this study, which was identical to another study where the occurrence of ASSuT isolates was 60% (Hopkins et al., 2012). In this study, similar to other reports (Wasyl and Hoszowski, 2012; Mulvey et al., 2013), ASSuT was the most frequent R-type detected, and resistance to amoxicillin and chloramphenicol was also an additional resistance frequently identified. For many years, ampicillin or trimethoprim–sulfamethoxazole was recommended as antimicrobial agents for the treatment of severe Salmonella human infections, which has contributed for the increased resistance levels and reduced efficacy of these agents (Winokur et al., 2000). In this study, resistance to ampicillin and trimethoprim–sulfamethoxazole was also observed, and tetracycline was the most active of the four antimicrobial compounds tested.
Distribution for Portuguese districts showed a higher percentage of Salmonella 1,4,[5],12:i:- isolates in coastal areas, namely Porto, Setúbal, and Aveiro. Most districts have reports of Salmonella 1,4,[5],12:i:-, including the Portuguese islands Azores, which suggests a wide distribution from the north to south of the country with a higher prevalence in the west, which may be explained by the higher human population density in these areas. Although in the period of this study an increasing annual incidence was observed, the number of reported isolates is low, which may be attributed to a decrease in the number of salmonellosis cases reported in the European Union in this period, including Portugal (EFSA, 2010). However, it is of concern that more than half of the reported cases in most districts are promoted by isolates presenting the ASSuT pattern. This is a critical issue for public health safety since ASSuT isolates are most frequently collected from young people (72.1%), who are more susceptible, and may lead to severe infections (EFSA, 2010; Ke et al., 2014).
In our study, quinolone and third-generation cephalosporin resistance was low. Nevertheless, resistance to ciprofloxacin, cefotaxime, and ceftazidime was mostly detected in isolates from young people, as already observed for the R-type ASSuT. Although most of Salmonella gastrointestinal infections are self-limiting and may not require antimicrobial therapy, treatment of complicated infections may be difficult due to high levels of resistance to frequently used antimicrobial compounds. Therefore, resistance to fluoroquinolones and third-generation cephalosporins can pose a major challenge to clinicians for the effective control of these infections, particularly relevant in young people (Ke et al., 2014).
Taking together with previous published results regarding biofilm formation (Seixas et al., 2014), it is particularly important to point out that almost all isolates resistant to fluoroquinolones and third-generation cephalosporins were also moderate biofilm producers, harboring the gcpA gene responsible to form biofilm under low nutrient conditions.
The molecular analysis of antibiotic resistance in R-type ASSuT isolates demonstrated that almost all these isolates, regardless of their origin, harbored the same resistance genes, blaTEM, strA-strB, sul2, and tetB, commonly found in several studies (Hopkins et al., 2010; Lucarelli et al., 2010; Antunes et al., 2011). These genes are present in a chromosomal resistance island and are typically associated to the European clone (Lucarelli et al., 2010), suggesting that the Portuguese clones may be related to those present in other European countries (Hopkins et al., 2010). Recently, a study demonstrated the high prevalence of metal tolerance, especially to silver and copper, associated with antimicrobial-resistant determinants in Salmonella 1,4,[5],12:i:- from Portugal, which may facilitate the dissemination of these isolates (Mourão et al., 2015).
The dissemination in Portugal may be also due to a relatively high diversity of clones during the study period, also detected in other reports, even among isolates from a single country (Soyer et al., 2009; Mandilara et al., 2013). PFGE identified 48 profiles with no relation to the epidemiological data, and this indicates that human cases were most likely sporadic and that the associated strains were not responsible for outbreaks. The unrelated epidemiological background of all cases supports this hypothesis. Interestingly, cluster analysis of PFGE profiles showed that all isolates belonging to cluster V were resistant to chloramphenicol, also including isolates with the highest rates of antimicrobial resistance with most isolates displaying additional resistances to at least two or three antimicrobials. BioNumerics cluster analysis showed that ASSuT and ACSSuT-resistant isolates were mainly included in two different clusters with a similarity level of 70% each, which may suggest that ASSuT-resistant isolates belong to the same clonal lineage, different from the ACSSuT-resistant isolates' lineage.
The high genetic diversity does not relate to the low variability of the virulence gene repertoire of the Salmonella 1,4,[5],12:i:- isolates, since only four different virulotypes were observed, also demonstrated in other reports (Capuano et al., 2013). This can be explained by the fact that most isolates were collected from humans and therefore, to promote invasive salmonellosis, it is required a common core of virulence genes (Suez et al., 2013). In addition, the spvC gene, frequently identified in isolates that cause systemic infections in immunocompromised patients (Guiney and Fierer, 2011), was detected in some isolates expressing resistance to fluoroquinolones and third-generation cephalosporins, which is particularly relevant in the clinical context.
Conclusions
Salmonella 1,4,[5],12:i:- is an emerging pathogen worldwide. In Portugal, although the number of reports was low, there was an increasing incidence of Salmonella 1,4,[5],12:i:- cases during this 5-year study. It was observed a wide distribution throughout the country, with reports of the R-type ASSuT in the majority of districts.
This rapid spread of Salmonella 1,4,[5],12:i:- R-type ASSuT in Portugal might be related with the diversity of pulsotypes, the advantage offered by R-type ASSuT, and the presence of a core of virulence genes, also along with biofilm formation.
Close surveillance of Salmonella 1,4,[5],12:i:- and its resistance patterns, especially important for young people, as revealed in this report, may help to prevent outbreaks and dissemination to nonproblematic districts, to track potential transmission pathways, and to rationalize salmonellosis antimicrobial therapeutics.
Footnotes
Acknowledgments
This study was conducted with the financial support of the “Centro de Investigação Interdisciplinar em Sanidade Animal, Faculdade de Medicina Veterinária da Universidade de Lisboa”—Project UID/CVT/00276/2013 (funded by Foundation for Science and Technology—FCT). R.S. and T.R.S. hold a PhD fellowship (SFRH/BD/75836/2011 and SFRH/BD/100571/2014, respectively) from FCT, Portugal.
Disclosure Statement
No competing financial interests exist.