
Abstract
Cantaloupes, marketed as “Rocky Ford,” were implicated in the U.S. multistate outbreak of listeriosis in 2011, which caused multiple fatalities. Listeria monocytogenes can survive on whole cantaloupes and can be transferred to the flesh of melons. The growth of L. monocytogenes on fresh-cut “Athena” and “Rocky Ford” cantaloupe cultivars during refrigerated storage was evaluated. Fresh-cut cubes (16.4 cm3) from field-grown cantaloupes were each inoculated with 5 log10 CFU/mL of a multi-strain mixture of L. monocytogenes and stored at 4°C or 10°C. Inoculated fresh-cut cubes were also: (1) continuously stored at 4°C for 3 days; (2) temperature-abused (TA: 25°C for 4 h) on day 0; or (3) stored at 4°C for 24 h, exposed to TA on day 1, and subsequently stored at 4°C until day 3. L. monocytogenes populations on fresh-cut melons continuously stored at 4°C or 10°C were enumerated on selected days for up to 15 days and after each TA event. Brix values for each cantaloupe variety were determined. L. monocytogenes populations on fresh-cut cantaloupe cubes stored at 4°C increased by 1.0 and 3.0 log10 CFU/cube by day 7 and 15, respectively, whereas those stored at 10°C increased by 3.0 log10 CFU/cube by day 7. Populations of L. monocytogenes on fresh-cut cantaloupes stored at 10°C were significantly (p < 0.05) greater than those stored at 4°C during the study. L. monocytogenes showed similar growth on fresh-cut “Athena” and “Rocky Ford” cubes, even though “Athena” cubes had significantly higher Brix values than the “Rocky Ford” fruit. L. monocytogenes populations on fresh-cut cantaloupes exposed to TA on day 1 and then refrigerated were significantly greater (0.74 log10 CFU) than those stored continuously at 4°C for 3 days. Storage at 10°C or exposure to TA events promoted growth of L. monocytogenes on fresh-cut cantaloupe during refrigerated storage.
Introduction
T
Contaminated melons have been a major vehicle of foodborne illness in the United States and have been responsible for 34 foodborne disease outbreaks between 1973 and 2011 (Walsh et al., 2014). There are numerous routes of contamination of melons with bacterial foodborne pathogens in agricultural fields and in food distribution chains. In 2011, the consumption of whole cantaloupes (Cucumis melo L) from Jensen Farms in the state of Colorado led to a large outbreak of listeriosis, which resulted in 147 cases of illness in 28 states, 33 fatalities, and 1 miscarriage (McCollum et al., 2013). This was the deadliest foodborne disease outbreak in nearly 90 years in the United States (CDC, 2011a). Melons were thought to be contaminated through transfer of Listeria monocytogenes from equipment surfaces in the packing shed, where melons were cleaned and boxed (FDA, 2015). L. monocytogenes is the etiologic agent of listeriosis, an opportunistic invasive illness, which occurs in immunocompromised individuals, older adults, infants, and pregnant women. It is associated with more severe symptoms, such as meningoencephalitis, bacteremia, neonatal sepsis, and premature abortion (Lund and O'Brien, 2011; Silk et al., 2012). Listeriosis is associated with a high hospitalization rate (94%), and it is the third leading cause of fatalities due to foodborne infections in the United States (19%) behind salmonellosis (28%) and toxoplasmosis (24%) (Scallan et al., 2011).
The cantaloupe rind provides a physical barrier that precludes entry of pathogens into the flesh of melons, but could serve as site of pathogen attachment (Ukuku et al., 2012). The surface topography of the cantaloupe rind, which consists of netting comprising rough bumps and grooves, harbors soil, bacteria, and fungi. These rind microsites shield adherent microbes and soil from cleaning and sanitizing agents, resulting in the incomplete inactivation of the microorganisms attached on the rind (Fan et al., 2009). Tissues that have been mechanically wounded and colonized by phytopathogens may facilitate entry of L. monocytogenes or Salmonella spp. into the mesocarp of cantaloupes (Richards and Beuchat, 2004). L. monocytogenes can survive on whole cantaloupes and can potentially be transferred to the mesocarp as melons are sliced. This study evaluated the effects of different cantaloupe cultivars, different storage temperatures, and temperature-abuse (TA) events on the growth of L. monocytogenes on fresh-cut cantaloupe cubes during storage.
Materials and Methods
Bacterial strains and cultures
The L. monocytogenes strains LI0072, LIS0077, and LIS0094 used in this study represent three distinct pulsed-field gel electrophoresis subtypes implicated in the U.S. multistate outbreak of listeriosis in 2011 associated with cantaloupes (Lomonaco et al., 2013). Frozen stock cultures were prepared for each bacterial strain on trypticase soy broth (TSB; BD, Franklin Lakes, NJ) in sterile vials containing 15% glycerol and stored at −80°C. Bacterial cultures were activated by streaking the stock cultures onto separate TSAYE (trypticase soy agar plus 0.6% yeast extract; Neogen, Lansing, MI) plates, which were incubated at 37°C for 20–24 h, and stored at 4°C. Before each experiment, active cultures of each bacterial strain were cultured on TSAYE plates incubated at 37°C for 20–24 h, followed by subculture from a single colony of each individual strain in 10 mL TSBYE (TSB plus 0.6% yeast extract) incubated at 37°C for 20–24 h. Cultures were centrifuged (Allegra 25R Centrifuge, Beckman Coulter, Danvers, MA) at 5000 g for 10 min and the pelleted cells resuspended in 10 mL of 0.1% peptone water (PW; BD). Cell suspensions were thoroughly vortexed and a multi-strain inoculum of L. monocytogenes was prepared by mixing aliquots (1 mL) of the cell suspensions for the individual bacterial strains in a 1:1:1 ratio. Bacterial populations of individual strain cultures were enumerated by performing separate serial dilutions of aliquots (1 mL) of cell suspensions and also the multi-strain inoculum in PW (0.1% PW); each dilution (100 μL) was plated in duplicate onto RAPID'L.mono medium (Bio-Rad, Hercules, CA) using a spiral plater (Microbiology International, Frederick, MD). Plates were incubated at 37°C for 24 h, and populations were determined by manual colony counts.
Cantaloupes
Two cantaloupe varieties, “Athena” and “Rocky Ford,” were included in this study to ascertain differences by cultivar on the growth of L. monocytogenes on fresh-cut pieces of cantaloupes during storage. The seeds of both “Athena” and “Rocky Ford” cantaloupe cultivars were obtained from Southern States (Fort Collins, CO); both cantaloupe cultivars were separately planted in soil in fields or high tunnels at the U.S. Department of Agriculture, Agricultural Research Service, Beltsville Agricultural Research Center (Beltsville, MD) and the University of Delaware Carvel Research and Education Center (Georgetown, DE). Melons were harvested when pedicels showed half-to full-slip/separation from melons, the harvested melons were field packed into fresh produce boxes (Uline, Pleasant Prairie, WI), and stored at 4°C within 2 h.
Effect of different temperatures (4°C, 10°C) on the proliferation of L. monocytogenes on fresh-cut cantaloupes during storage
Athena and Rocky Ford melons were analyzed separately; melons were washed with tap water to remove soil and debris and allowed to air dry. The rinds of Athena and Rocky Ford melons were removed using separate sterile knives. Cubes (16.4 cm3) of fresh-cut pieces of cantaloupes were prepared using sterile knives, and three cubes were packed into each 8 oz (235 cm3) square recycled polyethylene terephthalate (PET) deli containers (
Microbial analysis involved adding each cantaloupe cube to 25 mL of PW in separate Whirl-Pak filter bags and homogenizing (BagMixer MiniMix Stomacher, Interscience, Woburn, MA) for 2 min. Aliquots (1 mL) from each Whirl-Pak bag were serially diluted in PW, and each dilution was plated (100 μL) in duplicate onto RAPID'L.mono medium using a spiral plater and incubated at 37°C for 24 h to obtain L. monocytogenes populations.
The number of generations of L. monocytogenes on TA cantaloupe cubes was calculated by subtracting the initial populations inoculated onto the fresh-cut cantaloupes from the final populations after each TA event using the formula below:
where A was the final population at the end of time interval t2, and B was the initial population at the start of time interval t1.
The soluble sugar content (Brix values) was determined by placing cubes of fresh-cut cantaloupes into Whirl-Pak filter bags (Nasco, Fort Atkinson, WI), and gentle pressure was applied to separate fruit juice through the filter that was transferred into sterile vials. The pH of fruit juice of Athena and Rocky Ford cantaloupes was determined using an Orion benchtop pH meter with an Orion 915BNH Triode probe (Thermo Electron Corporation, Madison, WI) at 25°C. In addition, the Brix values of fruit juice of Athena and that of Rocky Ford cantaloupes were determined using a digital refractometer (Hanna Instruments, Inc., Woonsocket, RI).
Statistical analysis
Data were analyzed using JMP Pro 11 statistical software (SAS Institute, Inc., Cary, NC). Three replicates of each experiment were conducted and reported values are means of populations of L. monocytogenes and the standard errors of means (SEM). Data were analyzed using the fit model (linear model) for an analysis of variance (ANOVA) with cantaloupe cultivar (Athena, Rocky Ford) and storage temperature (4°C, 10°C) as independent variables and log-transformed means of bacterial populations (log10 CFU/cube) at various time points as the dependent variable. A factorial mode was selected to ascertain the effects (α = 0.05) of cultivar and temperature on populations of L. monocytogenes over storage time. The Tukey's HSD (honestly significant difference) test was used for pairwise comparisons to ascertain which mean values were significantly different (p < 0.05).
Results
L. monocytogenes populations increased on fresh-cut pieces of cantaloupes during storage
L. monocytogenes populations increased on fresh-cut pieces of both Athena and Rocky Ford cantaloupes during storage at 4°C and 10°C (Fig. 1). At 4°C, L. monocytogenes populations on fresh-cut Athena cantaloupes increased from 4.93 log10 CFU/cube on day 0 to 5.63 ± 0.07 log10 CFU/cube by day 7, while populations on fresh-cut Rocky Ford cantaloupes increased from 4.8 log10 CFU/cube on day 0 to 5.95 ± 0.44 log10 CFU/cube after 7 days of storage (Fig. 1). After 15 days of storage at 4°C, L. monocytogenes populations increased to 8.16 ± 0.16 and 7.84 ± 0.53 log10 CFU/cube on fresh-cut pieces of Athena and Rocky Ford cantaloupes, respectively (Fig. 1). At 10°C, L. monocytogenes populations on fresh-cut pieces of Athena cantaloupes increased by 3.6 log10 CFU/cube after 7 days of storage, while populations on Rocky Ford increased by 2.2 log10 CFU/cube, in the same time period (Fig. 1). The increase in populations of L. monocytogenes over time on fresh-cut cantaloupes stored at 10°C was significantly (p < 0.05) greater than on fresh-cut cantaloupes stored at 4°C. Storage temperature significantly affected growth of L. monocytogenes on fresh-cut cantaloupe. L. monocytogenes populations were not significantly different on fresh-cut Athena or Rocky Ford cantaloupes even though Athena cantaloupes had a mean Brix value of 9.93% ± 0.30%, which was significantly higher (p < 0.05) compared to that of Rocky Ford cantaloupes (6.04% ± 0.26%). The pH of juice from the flesh of Athena cantaloupes was 6.74, while that of juice from Rocky Ford flesh was 6.34 at 25°C.
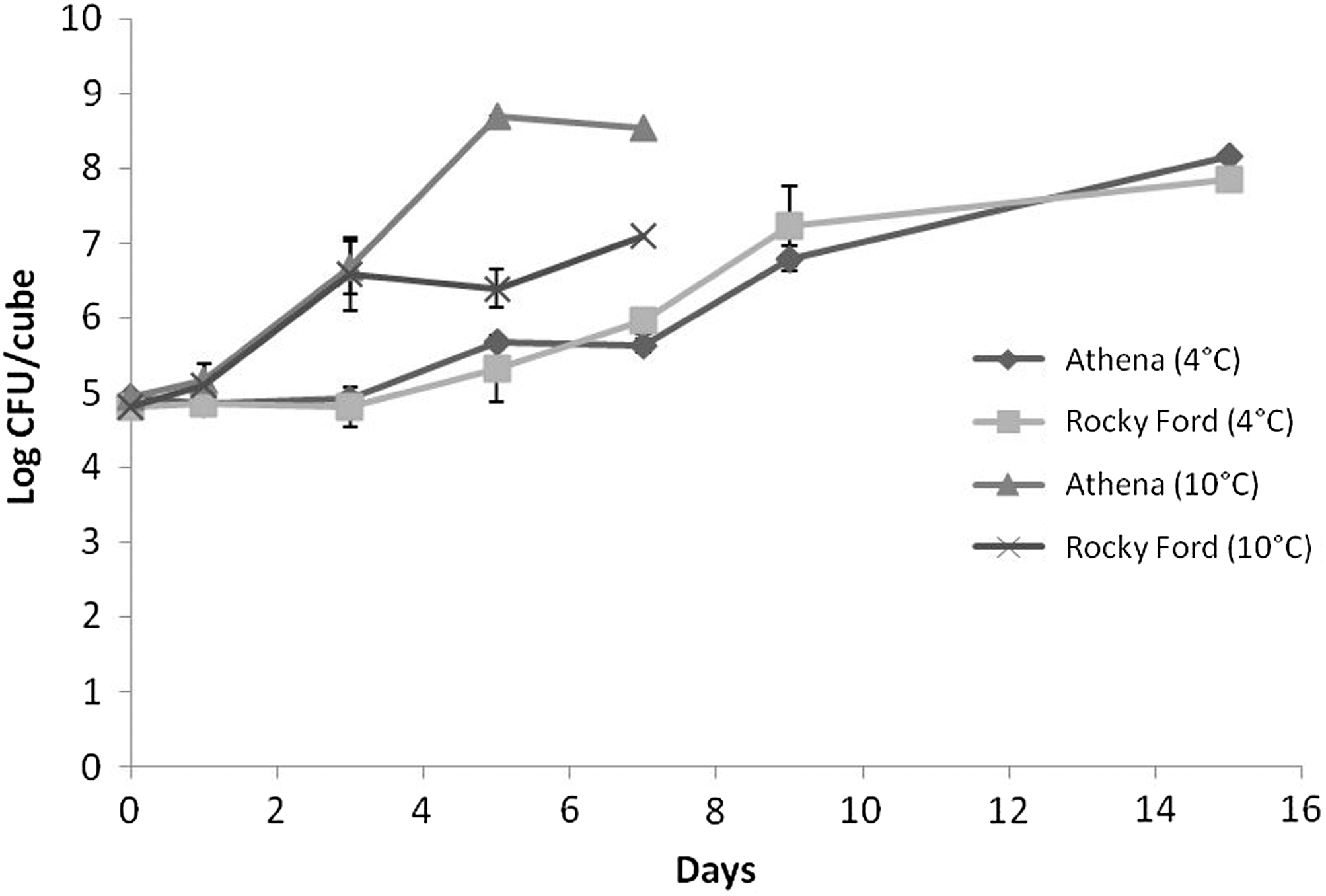
Change in populations of Listeria monocytogenes on fresh-cut pieces of Athena and Rocky Ford cantaloupe cultivars during storage at 4°C and 10°C. L. monocytogenes populations were significantly greater (p < 0.05) on fresh-cut melons stored at 10°C than at 4°C. Populations were not significantly affected by cultivar (p > 0.05). Values shown are means (n = 3) plus standard errors (error bars).
TA (25°C; 4 h) during storage of fresh-cut pieces of cantaloupes contaminated with L. monocytogenes
L. monocytogenes populations on fresh-cut pieces of cantaloupes continuously stored at 4°C did not change over 3 days (Table 1). Populations of L. monocytogenes on fresh-cut pieces of cantaloupes exposed to TA on day 0 were not significantly (p > 0.05) different compared to those on fresh-cut cantaloupes continuously stored at 4°C for 4 h. Similarly, populations of L. monocytogenes on fresh-cut pieces of cantaloupes, which were stored at 4°C for 24 h and then exposed to TA on day 1, were not significantly different from those on fresh-cut cantaloupes, which were continuously refrigerated through day 1. On day 3, populations of L. monocytogenes on fresh-cut cantaloupes exposed to TA on day 1 and then stored at 4°C until day 3 were significantly greater (0.63–0.84 log CFU/cube) than populations on melon cubes stored continuously at 4°C. The populations of L. monocytogenes on fresh-cut pieces of cantaloupe stored at 4°C for 24 h and exposed to TA on day 1, then refrigerated until day 3, increased by approximately two bacterial generations (n).
L. monocytogenes populations were determined immediately after exposure to TA on day 0.
L. monocytogenes populations were determined immediately after TA on day 1 and after 4°C storage until day 3.
Within columns (days), mean populations followed by a different superscript lowercase letter (a,b) indicate that values are significantly (p < 0.05) different.
TA, temperature-abuse.
Discussion
The implicated source of the L. monocytogenes strains that caused the U.S. multi-state outbreak of listeriosis in 2011 was the cantaloupe packing shed (FDA, 2015). The transfer of pathogens to the stem-scar areas of melons or infiltration of wash water contaminated with pathogens into melons can cause contamination of the flesh (Richards and Beuchat, 2004). In addition, bacteria can survive on cantaloupe rind and be transferred to the flesh during cutting (Ukuku et al., 2012). Recent work has also shown that L. monocytogenes can proliferate on the stem-scars of cantaloupes when stored at 25°C (Nyarko et al., 2016).
The storage temperatures used in our study, 4°C and 10°C, were selected to contrast refrigerated and improper storage of fresh-cut pieces of cantaloupes. L. monocytogenes populations increased by ∼1 log10 CFU/cube on fresh-cut pieces of cantaloupes stored at 4°C by day 7, but the populations also displayed a long lag phase duration, suggesting that refrigerated storage of fresh-cut pieces of cantaloupe cubes slowed growth of the pathogen. L. monocytogenes populations increased by 3.0–3.5 log10 CFU/cube when stored at 4°C for 15 days, but the quality of the fresh-cut cantaloupes severely deteriorated after day 7. Our results are mostly in agreement with a study that modeled the growth of L. monocytogenes on fresh-cut cantaloupe, honeydew, and watermelon, which predicted ∼4 log10 CFU increase by day 15 of storage at 5°C and ∼1 log10 CFU increase by day 6 of storage at 4°C (Danyluk et al., 2014). Unlike Escherichia coli O157:H7, which did not grow when stored at 5°C for 34 h on fresh-cut melon fruit tissue (Del Rosario and Beuchat, 1995), L. monocytogenes grew at 4°C on fresh-cut cantaloupe in our study.
Fresh-cut cantaloupe cubes stored at 10°C resulted in significantly greater increases of L. monocytogenes populations (∼3.5 log10 CFU/cube) compared to those that were stored at 4°C (∼1 log10 CFU/cube) over 7 days. In a previous study, populations of L. monocytogenes on fresh-cut pieces of cantaloupes stored continuously for 7 days at 12°C were greater than on those stored at 8°C, which were also greater than on those stored at 4°C (Huang et al., 2015). Previous work has shown that Salmonella spp. populations on fresh-cut cantaloupes, honeydew, and watermelon pieces increased by at least 1 log10 CFU/g when stored at 10°C for up to 10 days (Ukuku and Sapers, 2007). In our study, there were no significant differences in the growth of L. monocytogenes on Athena or Rocky Ford cultivars. However, at 10°C, L. monocytogenes populations on Athena cubes were 2.29 and 1.43 log10 CFU/cube greater than on Rocky Ford melons on day 5 and 7, respectively. The significantly greater Brix value of the Athena melons compared to the Rocky Ford melon indicates that maturity or at least sugar content at the time of cubing may have more influence on L. monocytogenes growth than cultivar. Future work will evaluate if stage of maturity can promote greater growth of L. monocytogenes in fresh-cut melons. Post-harvest storage has been found to cause biochemical changes in pH, Brix values, organic acids, texture, appearance, and sugars in freshly cut cantaloupes (Lamikanra et al., 2000). In addition, lower storage temperatures (4°C) were found to slow down the rate of biochemical changes in cantaloupes compared to higher storage temperatures.
In a picnic or home party-type setting, melons may be cut and held at temperatures above refrigeration for several hours and the remaining portions may be refrigerated for consumption the next day. In our study, exposure of fresh-cut melons to TA conditions (25°C, 4 h) on day 0 did not result in a significant increase in L. monocytogenes populations compared to those on melons stored continuously at 4°C. TA of fresh-cut cantaloupes interrupted by refrigerated storage led to higher populations of L. monocytogenes populations compared to those continuously refrigerated fresh-cut cantaloupes on day 3. Other authors found that Salmonella spp. on fresh-cut cantaloupe increased by 0.3 and 1 log10 CFU/g after 3 and 5 h of storage, respectively, at 22°C compared to inoculated fresh-cut cantaloupes continuously stored at 4°C (Ukuku and Sapers, 2007). Populations of L. monocytogenes and Salmonella enterica increased by 0.51 and 0.6 log10 CFU, respectively, on fresh-cut cantaloupes after exposure to TA (25°C for 4 h) events on day 2 of refrigerated storage (Huang et al., 2015). In our study, TA of fresh-cut cantaloupe cubes that interrupted refrigerated storage on day 1 led to higher populations of L. monocytogenes than to the continuously refrigerated fresh-cut cantaloupe cubes on day 3.
Conclusion
Growth of L. monocytogenes on fresh-cut cantaloupe occurs during storage at 4°C, but it is enhanced at higher storage temperatures (10°C). Refrigerated storage of inoculated fresh-cut cantaloupe interrupted by TA events promoted growth of L. monocytogenes. Cultivar did not influence growth of L. monocytogenes, and further work is needed to investigate the effect of cantaloupe maturity and sugar content on growth and survival of L. monocytogenes. Regardless, consistent storage at 4°C is the best method to limit growth of L. monocytogenes on fresh-cut cantaloupes.
Footnotes
Acknowledgments
The authors are grateful to Patrick Spanninger and Dr. Gordon Johnson of the University of Delaware, Newark and Georgetown, DE; Kathryn White, Dave Clark, Bin Zhou, and Richard Stonebraker of the Environmental Microbial and Food Safety Laboratory, U.S. Department of Agriculture, Agricultural Research Service, Beltsville, MD; and Yi Chen, Dumitru Macarisin, Michael Mahovic, Annemarie Buchholz, and Mickey Parish of U.S. Food and Drug Administration for their assistance with this study. This study was funded by Inter-Agency Agreement 224-11-2046 between FDA and USDA ARS, Specific Cooperative Agreement 58-1245-4-111 between USDA ARS and University of Delaware, and USDA ARS Project Number 8042-32420-005-00 “Microbial Ecology and Safety of Fresh Produce.”
Disclosure Statement
No competing financial interests exist.