
Abstract
Poultry is a major source of Campylobacter, which can cause foodborne bacterial gastroenteritis in humans. Additionally, poultry-associated Campylobacter can develop resistance to important antimicrobials, which increases the risk to public health. While broiler chickens have been the focus of many studies, the emergence of antimicrobial-resistant Campylobacter on layer farms has not received equal attention. However, the growing popularity of cage-free and organic layer farming necessitates a closer assessment of (1) the impact of these farming practices on the emergence of antimicrobial-resistant Campylobacter and (2) layers as a potential source for the transmission of these pathogens. Here, we showed that the prevalence of Campylobacter on organic and conventional layer farms was statistically similar (p > 0.05). However, the average number of Campylobacter jejuni-positive organically grown hens was lower (p < 0.05) in comparison to conventionally grown hens. Campylobacter isolated from both production systems carried antimicrobial resistance genes. The tet(O) and cmeB were the most frequently detected genes, while the occurrence of aph-3-1 and blaOXA-61 was significantly lower (p < 0.05). Farming practices appeared to have an effect on the antimicrobial resistance phenotype, because the isolates from organically grown hens on two farms (OF-2 and OF-3) exhibited significantly lower resistance (p < 0.05) to ciprofloxacin, erythromycin, and tylosin. However, on one of the sampled organic farms (OF-1), a relatively high number of antimicrobial-resistant Campylobacter were isolated. We conclude that organic farming can potentially impact the emergence of antimicrobial-resistant Campylobacter. Nevertheless, this impact should be regularly monitored to avoid potential relapses.
Introduction
T
The aforementioned observations along with consumer demand have driven the rise in organic farming as an alternative to conventional poultry production, which includes the use of antimicrobials in treating and preventing (prophylaxis) diseases and as growth promoters (Wegener, 2003; Sapkota et al., 2007; Holtcamp, 2011). Since organic farming reduces the reliance on antimicrobials, it is predicted to significantly decrease the prevalence of antimicrobial-resistant bacteria, including Campylobacter, in the food chain (Fanatico et al., 2009; Sapkota et al., 2011, 2014). In fact, Luangtongkum et al. (2006) showed that the prevalence of antimicrobial-resistant Campylobacter was lower in organically farmed poultry. For example, only 2% of Campylobacter isolated from organically raised birds were resistant to fluoroquinolones in comparison to up to 67% of isolates retrieved from conventionally raised birds. However, a relatively large number of isolates from organically raised birds were still found to be resistant to certain antimicrobials such as tetracycline (Luangtongkum et al., 2006). The latter raises questions in regard to factors that might drive the persistence of antimicrobial-resistant isolates under organic farming practices and the contribution of these isolates to the pool of resistant bacteria that might affect public health.
Most of the epidemiological studies to date have focused on antimicrobial-resistant Campylobacter in meat-producing birds such as broilers, because they pose the highest risk for consumers (Sahin et al., 2015). In comparison, laying hens (layers) have not received equal attention. However, due to the increase in the transition to the consumer-preferred organic and cage-free farming in shell egg production, there might be a need to assess layers as potential reservoirs for antimicrobial-resistant Campylobacter. This is important, because layer farming is a massive operation. In the United States, it is estimated that the size of the table egg producing layers was 288 million in 2016. The cage-free layers comprised 23.6 million, of which 11.4 million hens were certified as organic by the U.S. Department of Agriculture (USDA, 2016). These hens, by regulation, should have access to outside the farming houses. Therefore, antimicrobial-resistant Campylobacter associated with these hens can pose a potential risk of environmental contamination and subsequent transmission via wildlife and other farm animals, workers and run-off (Ahmed et al., 2013). Notably, Campylobacter can survive in feces of layers and in poultry litter for several days, which suggested that the land application of these byproducts as fertilizers might also contribute to the dissemination of the pathogens (Kassem et al., 2010; Ahmed et al., 2013). Furthermore, organically raised layers are consumed as roasting chickens at the end of their production cycle, while conventional layers are processed as protein supplements for pet food or as stewing hens. Hence, layers can also pose a food safety risk, although significantly lesser than broilers. In this study, we investigated the occurrence of antimicrobial-resistant Campylobacter on organic and conventional cage-free layer farms to assess the impact of organic farming on the prevalence of antimicrobial-resistant isolates and assess potential public health risks associated with layer farming in general.
Materials and Methods
Collection of fecal samples from layer flocks
Between November and January (2014–2015), 360 fresh fecal samples (n = 60/farm) were collected from three conventional (CF) and three organic (OF) cage-free layer farms that were located in North-Western Ohio (USA). The farms housed flocks (10,000 cage-free hens each) that were 24–27 (CF-1 and OF-1), 41–44 (CF-2 and OF-2), and 62–63 (CF-3 and OF-3) weeks old, respectively (Supplementary Table S1; Supplementary Data are available online at
Isolation and identification of Campylobacter spp.
To isolate Campylobacter, fecal samples (1 g) were suspended in 9 mL of Preston enrichment broth with Campylobacter growth supplements (SR0117 and SR0232; Oxoid) (Kashoma et al., 2016). The suspensions were incubated under microaerobic conditions (5% O2, 10% CO2, and 85% N2) at 42°C for 48 h. After enrichment, putative Campylobacter were isolated on modified charcoal cefoperazone deoxycholate agar (mCCDA) that contained a selective supplement, SR0155 (Oxoid). The isolates were purified on Mueller–Hinton (MH) agar containing a selective supplement (SR0117; Oxoid) as described elsewhere (Kashoma et al., 2015).
DNA templates from each purified colony were extracted using the boiling method (Kashoma et al., 2016). The isolates were then confirmed as Campylobacter and their species was identified using a previously described multiplex polymerase chain reaction (PCR) (Sanad et al., 2011). If only the genus-specific PCR fragment was detected, the isolates were designated as other Campylobacter. Isolates were identified as either Campylobacter coli or Campylobacter jejuni if they harbored both the genus- and the species-specific PCR fragments. C. jejuni 81–176 and C. coli (ATCC 33559) were used as positive controls, while sterile water constituted a no-template (negative) control.
Detection of antimicrobial resistance genes
A previously described multiplex-PCR assay was used to detect the occurrence of four antimicrobial resistance genes: blaOXA-61 (ampicillin resistance), aph-3-1 (aminoglycoside resistance), tet(O) (tetracycline resistance), and cmeB (encoding the multidrug efflux pump) in the Campylobacter isolates (Obeng et al., 2012). DNA templates were prepared as described above, and Campylobacter isolates that were previously confirmed to harbor these genes (Kashoma et al., 2016) were used as positive controls.
Evaluating antimicrobial susceptibility by using the broth microdilution method
All the Campylobacter isolates were tested for susceptibility to five antimicrobial agents, ciprofloxacin, erythromycin, tylosin, spectinomycin, and tetracycline (Sigma-Aldrich Co.), using the broth microdilution method (Ge et al., 2013). Briefly, 96-well plates containing twofold serial dilutions of each antimicrobial agent were prepared. The isolates were then suspended in MH broth to achieve an optical density (OD600) of 0.05, and aliquoted (100 μL) to each well of the antimicrobial-containing plates. After incubation under microaerobic conditions at 42°C for 24 h, the plates were assessed by both visual examination and using a microplate spectrophotmeter to determine growth inhibition (Ge et al., 2013; Kashoma et al., 2016). Control wells contained Campylobacter suspensions without antimicrobials, and sterile MH broth containing each of the antimicrobials, respectively. C. jejuni (81–176) and C. coli (ATCC 33559) were used as control strains in this experiment. Minimum inhibitory concentrations and resistance breakpoints were determined as described previously (Ge et al., 2013). Isolates that were resistant to three or more antimicrobial classes were considered as multidrug resistant (MDR) (Hakanen et al., 2003).
Statistical analysis
Statistical analysis was performed using one-way analysis of variance followed by Tukey's posttest. Alternatively, Student's t-test and the chi-square test were used for comparisons between two groups or to determine the independence of the data, respectively. A p value of <0.05 was considered statistically significant.
Results and Discussion
A total of 248 (69%) Campylobacter isolates were identified from the 360 fecal samples collected from all the farms. Coincidently, the Campylobacter isolates were equally divided between the production types, each yielding 124 isolates. A previous study reported that the number of Campylobacter-positive laying hens between November and April was similar (Doyle, 1984). In addition, our results showed that the age of the flock did not significantly affect (p > 0.05) the prevalence of total Campylobacter between the farms that belonged to different production types. Therefore, we averaged the prevalence of Campylobacter across the organic farms and compared it to that in the conventional farms to focus on the effect of production type. The average prevalence of Campylobacter in organic layers (69% ± 21%) was not statistically different (p > 0.05) in comparison to conventional layers (69% ± 8%) (Fig. 1). However, the occurrence of Campylobacter (93%) in samples collected from organic farm 1 (OF-1) was higher than OF-2 (55%) and OF-3 (58%), respectively. In comparison, the occurrence of Campylobacter on conventional farms (CF-1, 2, and 3) was less variable and ranged between 60% and 72%. The prevalence of C. coli was also not statistically different between organic (58% ± 22%) and conventional layers (40% ± 6%) (p > 0.05) (Fig. 1). The occurrence of C. coli (83%) in samples collected from OF-1 was higher than OF-2 (40%) and OF-3 (52%), respectively. The occurrence of C. coli in CF-1, 2, and 3 was less variable and ranged between 36% and 47%. Notably, the average prevalence of C. jejuni in organic layers (4% ± 2.5%) was significantly lower (p < 0.05) in comparison to conventional layers (25% ± 6.5%) (Fig. 1). C. jejuni was the lowest in OF-3, where the pathogen occurred in only 1 out of 60 fecal samples. Studies on the prevalence of Campylobacter in layer and organic flocks are scarce. However, in one report, approximately up to 25% of individually caged laying hens were positive for C. jejuni at peak isolation rates, while 8.1% of the hens were found to chronically excrete the pathogen (Doyle, 1984). Furthermore, it has been shown that the prevalence of Campylobacter was 89% and 66% in organic and conventional broilers, respectively, and C. jejuni was the predominant species in both types of broilers (Luangtongkum et al., 2006). Therefore, while the prevalence of Campylobacter in our study was comparable to other reports, we also show that the distribution of the Campylobacter species might be affected by the production system. This observation can have an impact on the management and risk assessment of the production system and the transmission of the pathogens.
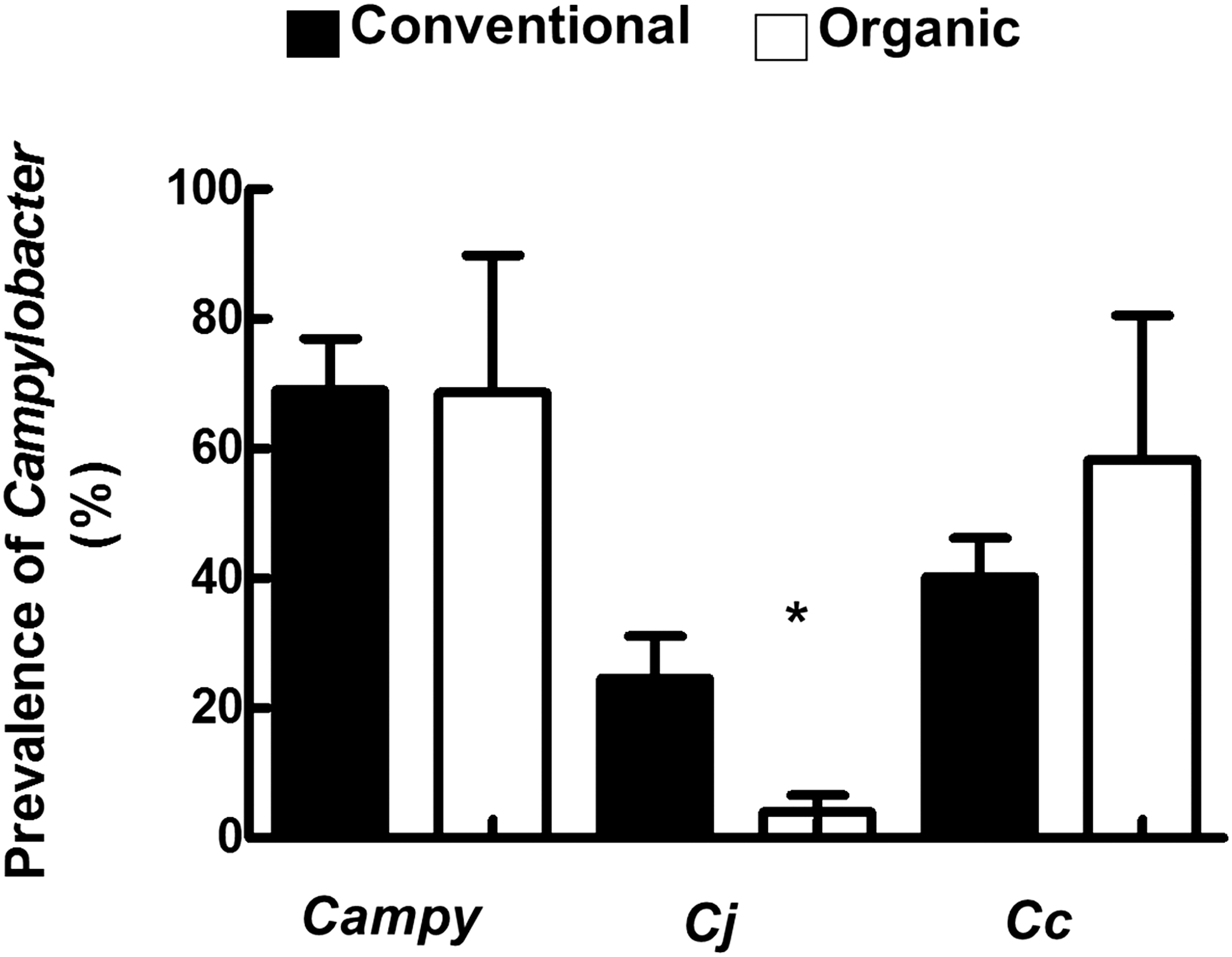
The average prevalence of Campylobacter across production type. Campy: all Campylobacter species; Cj, Campylobacter jejuni; Cc, Campylobacter coli. Campylobacter was isolated from fresh fecal samples (n = 360; 60/farm). Confirmation and identification of Campylobacter species were accomplished using a multiplex PCR. *Denotes statistically significant difference (p < 0.05) between Cj conventional and Cj organic. Black and white bars indicate conventional and organic farms, respectively. PCR, polymerase chain reaction.
We screened the Campylobacter isolates for potential antimicrobial resistance genes, which included blaOXA-61 (ampicillin), aph-3-1 (aminoglycoside), tet(O) (tetracycline), and cmeB (multidrug efflux pump). The occurrence of these genes in Campylobacter isolated from organically raised hens was comparable (p > 0.05) to those from conventionally raised hens (Fig. 2A). The average number of isolates that were positive for multiple genes was also statistically comparable (p > 0.05) in organic and conventional samples (Fig. 2A). Regardless of the production system, the most frequently detected genes were tet(O) and cmeB, while the detection of aph-3-1 and blaOXA-61 was significantly lower (p < 0.05) in comparison to the aforementioned genes (Fig. 2A). The tet(O) (87%) and cmeB (98%) were most frequently detected in isolates from OF-1, while aph-3-1 and blaOXA-61 were not detected on that farm. Taken together, our data suggest that the Campylobacter isolates occurring in both systems have the genetic potential that is necessary for exhibiting resistance to antimicrobials. However, the latter requires further investigation to determine (1) the expression and functionality of the detected genes and (2) mutations that might be associated with antimicrobial resistance. For example, the resistance of Campylobacter to macrolides (erythromycin) and fluoroquinolones (ciprofloxacin) has been associated with distinct point mutations in the 23S rRNA gene and gyrA (DNA gyrase subunit A), respectively (Piddock et al., 2003; Gibreel et al., 2005). Regardless, the number of C. jejuni in organic samples was low and most of the antimicrobial resistance genes occurred in C. coli. This suggested that C. coli shed from organic layers might serve as a reservoir for antimicrobial resistance genes. Since organic layers have access to outside the farming houses, shedding of pathogens that carry antimicrobial resistance markers should be considered in terms of potential environmental contamination and cognate ramifications such as the transmission of pathogen and the acquisition of the markers by other bacterial pathogens.
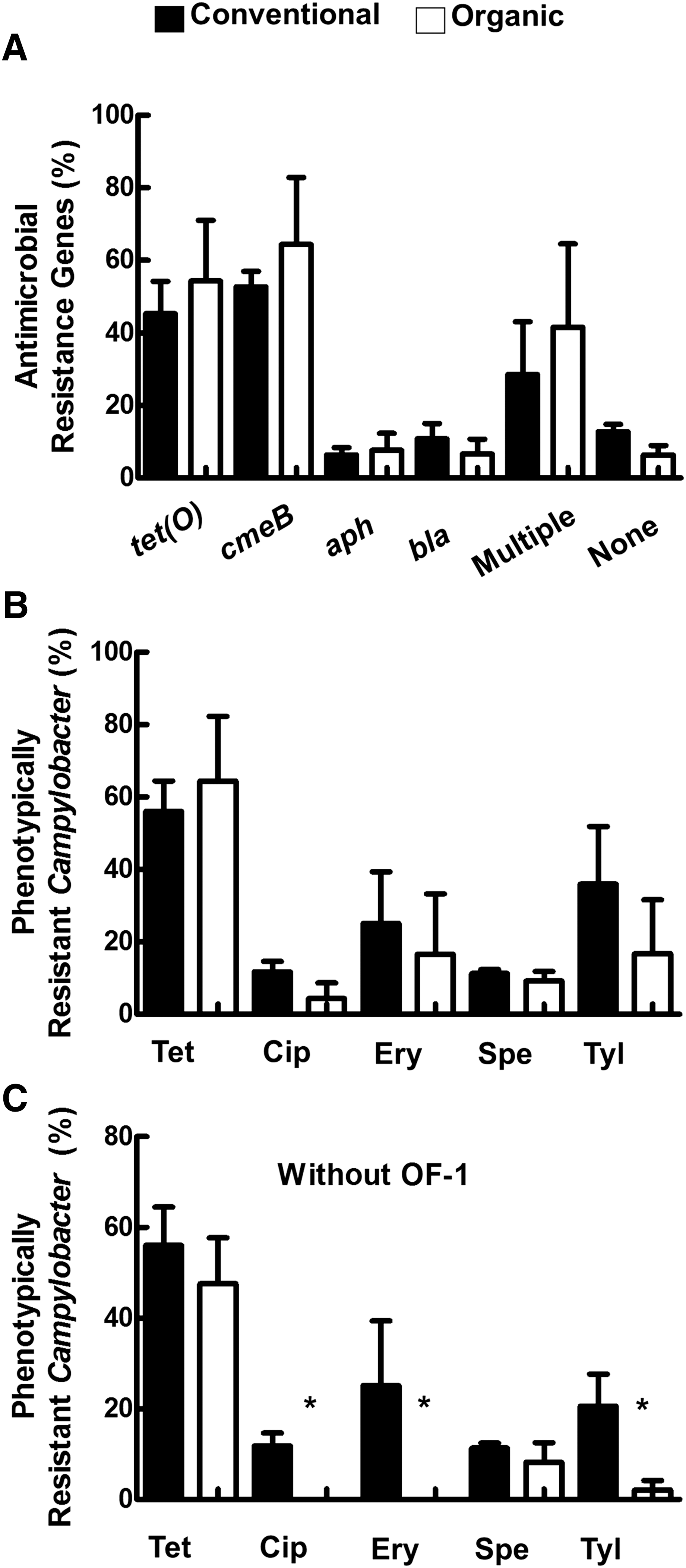
The carriage of antimicrobial resistance markers does not always clearly reflect the antimicrobial resistance phenotype (Kashoma et al., 2015, 2016). Therefore, we determined the susceptibility of the Campylobacter isolates, focusing on C. jejuni and C. coli only, to five different antimicrobials. When considering the average number of antimicrobial-resistant Campylobacter, there was no significant difference between organic and conventional farms (Fig. 2B). The most frequent resistance was observed against tetracycline under both operations, while the lowest was observed against ciprofloxacin in isolates from organically grown hens (Fig. 2B). In a previous study, >80% and 50% to 60% of Campylobacter isolated from conventionally and organically raised broilers were found to be resistant to tetracycline, respectively (Luangtongkum et al., 2006). Furthermore, C. jejuni from conventional layers in South Africa exhibited resistance to tetracycline (100% of isolates), ciprofloxacin (76%), gentamicin (81%), and erythromycin (57%) (Bester and Essack, 2008). The resistance reported in the latter study is considerably higher than our observations. This might be due to differences in farming practices or to the antimicrobial use guidelines.
A total of 11 C. jejuni and 7 C. coli from conventionally grown hens (CF-1, 2, and 3) were MDR (data not shown). In comparison, 2 C. jejuni and 25 C. coli from organically grown hens (only in OF-1) were MDR (data not shown). Subsequently, a closer inspection of our data revealed that resistance to ciprofloxacin and erythromycin was only detected in isolates from OF-1 (Supplementary Table S1). Furthermore, 98% and 46% of the Campylobacter isolates from OF-1 were resistant to tetracycline and tylosin, which was higher than OF-2 (37.5% and 4.2%) and OF-3 (57.7% and 0%), respectively (Supplementary Table S1). Due to these discrepancies and previous literature that showed a mitigating effect of organic farming on antimicrobial resistance (Luangtongkum et al., 2006; Sapkota et al., 2011, 2014), we reanalyzed our data without the inclusion of OF-1. We found that Campylobacter from the other two organic farms (OF-2 and OF-3) exhibited significantly lower resistance (p < 0.05) to ciprofloxacin, erythromycin, and tylosin (Fig. 2C). Furthermore, none of the C. jejuni from OF-2 was resistant to any of the antimicrobials, while one C. jejuni was resistant to tetracycline in OF-3. Taken together, our reanalysis suggested that organic farming can reduce the emergence of antimicrobial resistance.
Organic layer farming appeared to have an impact on the population of Campylobacter as C. coli was the predominantly isolated species. The latter requires further investigation and might be attributed to other farming practices. In addition to the merits of organic layer farming on animal welfare, it also appears to mitigate the emergence of antimicrobial resistance. Although the exact reason behind the relatively high number of antimicrobial-resistant Campylobacter in OF-1 is not known, lapses in management practices might have been a potential factor that affected the emergence of antimicrobial resistance on this farm. For example, we observed that the biosecurity and hygiene controls on farm OF-1 were less stringent in comparison to the other farms. Therefore, strict guidelines and periodic surveillance should be maintained to facilitate interventions and curb lapses that might reduce the impact of organic farming on antimicrobial resistance.
Footnotes
Acknowledgments
Research in the Rajashekara laboratory is supported by funds from the Agriculture and Food Research Initiative (AFRI, US Department of Agriculture), the Ohio Agricultural Research and Development Center, and the Ohio State University.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.