
Abstract
Numerous studies have assessed the efficacy of phage-based methods to inhibit Salmonella contamination in food products. As with most antibacterials, bacteria can develop resistance to phage in vitro. Here, we applied a single broad-spectrum Salmonella phage, vB_SalS_SJ_2 (SJ2; 108 PFU; MOI = 10), to Salmonella-contaminated meat and eggs to quantify the development of resistance in actual food matrices. Treatment with a single phage significantly reduced Salmonella Typhimurium contamination in both ground pork and liquid egg at various time points. Similarly, the same phage significantly reduced Salmonella Enteritidis in both food matrices. Efficacy was temperature dependent as larger reductions were seen at higher temperatures (21°C) versus lower temperatures (4°C) at 24 h. Following phage treatment, over 10,000 Salmonella isolates were examined for resistance to the treatment phage. The percentages of phage-resistant Salmonella (either serovar) recovered from phage-treated versus untreated pork did not differ. Conversely, significantly (p < 0.05) higher percentages of phage-resistant Salmonella Typhimurium (92.50% vs. 0.56% of control) and Salmonella Enteritidis (50.83% vs. 0.56% of control) isolates were observed in phage-treated versus untreated egg samples after incubation at room temperature for 48 h. Taken together, these data indicate that the food matrix may influence the emergence of phage resistance with resistance developing more rapidly in foods with less complex microbial communities. Future studies will focus on the impact the development of resistance in production and processing settings may have on the efficacy of phage treatments for longer term biocontrol of pathogens.
Introduction
S
The majority of the aforementioned studies used phage cocktails or mixtures of several distinct phages. As phages can be highly specific, mixtures of unrelated phages can increase the spectrum of the treatment. This would be especially beneficial in food processing settings where little diagnostic information is available regarding the contaminating bacteria outside of species.
As with most antibacterials, targeted bacteria can develop resistance to phages in vitro. The potential for phage resistance in the targeted host is also another justification for use in polyphage preparations as the simultaneous development of resistance to multiple, unrelated phages is less likely. There are remarkably few studies, however, that examine the extent to which phage resistance actually occurs in different food matrices and the impact it may have on efficacy. The present study was designed to measure the efficacy of a single broad-spectrum phage (vB_SalS_SJ_2 [SJ2]; Zhang et al., 2015) in reducing Salmonella contamination in ground pork and liquid egg, and concurrently determine, on a larger scale, how often and how soon targeted bacteria become phage resistant within a food matrix.
Materials and Methods
Bacteria and bacteriophage
Salmonella Typhimurium γ4232 (nalidixic acid resistant) was previously isolated from a diseased pig (Ebner and Mathew, 2000) and Salmonella Enteritidis was kindly provided by Dr. Arun K. Bhunia (Purdue University, West Lafayette, IN). Phage vB_SalS_SJ_2 (SJ2) was originally isolated from wastewater using Salmonella Typhimurium as a host (Wall et al., 2010). It is a siphovirus with a genome size of 152,460 bp and contains 197 predicted genes and showed the broadest spectrum among our library of Salmonella phages, with high lytic capacity against Salmonella serovars Typhimurium, Enteritidis, Indiana, and Kentucky (Zhang et al., 2015).
Phage treatment of Salmonella-contaminated food products
Ground pork and shell eggs were purchased from local retail stores. All samples were tested by culturing for Salmonella before any further phage testing to confirm that they were Salmonella free. To test phage efficacy in ground pork, 15 g samples were placed in a sterile stomacher bag (VWR International, SCLO7012-VW1) and inoculated with 107 CFU of either Salmonella Typhimurium (n = 100) or Salmonella Enteritidis (n = 100) in 100 μL of Luria-Bertani broth, and massaged by hand thoroughly for 30 s to ensure even contamination throughout the sample.
Samples assigned for phage treatment (n = 50) were then inoculated with 108 PFU of phage SJ2 suspended in 100 μL of phosphate buffered saline (PBS). Control samples (n = 50) received 100 μL of PBS only. All samples were once again massaged by hand for 30 s after inoculation and incubated under one of five conditions (n = 10 for both control and phage treatment per condition): 21°C (RT) for 24 h, RT for 48 h, 4°C for 24 h, 4°C for 1 week, and 4°C for 2 weeks.
After incubation, meat samples were resuspended in 100 mL of buffered peptone water (Becton, Dickinson and Company, 212367), the suspension was serially diluted and concentrations of Salmonella were determined by plating on XLT4 agar plates (Becton, Dickinson and Company, 223420) containing 50 μg/mL of nalidixic acid (Alfa Aesar, 3374-05-8) for Salmonella Typhimurium or XLD agar plates (Becton, Dickinson and Company, 221284) for Salmonella Enteritidis, as XLT4 plates greatly inhibited the growth of the Salmonella Enteritidis strain used in this study (data not shown). The entire process was repeated independently three times for a total of 300 samples per incubation condition per Salmonella serovar.
To test the efficacy of phage treatment in liquid egg, egg shells were wiped with 70% alcohol and air-dried before breaking to avoid unintended contamination from outer surfaces. Eggs were broken and homogenized for 2 min with a Proctor Silex Easy Mix 5-Speed Hand Mixer (Hamilton Beach Brands, Inc., Glen Allen, VA) at speed 1. Liquid egg samples (15 mL; n = 10 per treatment group per incubation condition per Salmonella serovar) were inoculated with 107 CFU of Salmonella Typhimurium or Salmonella Enteritidis in 100 μL of PBS and vortexed.
Samples assigned for phage treatment were treated with 108 PFU of SJ2 in 100 μL of PBS and control samples received 100 μL of PBS only. Samples were once again vortexed after inoculation and incubated under one of the same five conditions previously described for pork experiments. After incubation, Salmonella concentrations were quantified as previously described. Each experiment was repeated independently three times.
Phage susceptibility of Salmonella after phage treatment
To test the development of phage resistance in the challenge Salmonella after treatment with phage SJ2, 24 Salmonella isolates from each phage-treated sample (3 replicates of 10 samples per treatment [time and incubation period] across two challenge Salmonella serovars and two food matrices) and 12 Salmonella isolates from each control sample (samples not treated with phage) were collected. This model produced more than 10,000 Salmonella isolates for resistance testing. A small amount (∼size of toothpick tip) of each colony was picked from agar plates using sterilized toothpicks and resuspended in individual tubes containing 900 μL of PBS.
To determine an appropriate amount of a bacterial biomass for suspension, at the beginning of the study, different sizes (size of toothpick tip, an entire 1-mm-diameter colony, an entire 2-mm-diameter colony, etc.) of phage-susceptible colonies were grown on overlay both with and without phage. This was done to ensure that the biomass in suspension was large enough to form bacterial lawns on overlay with no growth inhibitor (phage), but not too large to result in a false positive due to overgrowth. Following a brief vortex, 5 μL of each suspension was dropped onto an overlay containing a concentration of SJ2 (1.4 × 109 PFU/mL) previously determined to completely inhibit growth of the challenge (parent) Salmonella strains and the plates were incubated at 37°C overnight. Salmonella isolates that formed complete bacterial lawns at dropped spots were considered resistant to phage SJ2.
Statistical analysis
A randomized block design with replications was used to compare the concentrations of Salmonella and the percentages of phage-resistant Salmonella colonies in both ground pork and liquid egg under different treatments. The experiment was blocked on replicate. Treatments were fixed effects, and experimental triplicates were random effects. Analysis was done using a mixed model analysis of variance (SAS 9.4) and ANOVA F-tests were used to compare differences between the treatments. In addition, percentages of phage-resistant colonies were transformed into ranks. p-Values less than 0.05 were considered as significantly different.
Results
Phage treatment of Salmonella-contaminated ground pork
In Salmonella Typhimurium-contaminated ground pork, samples treated with phage SJ2 had significantly lower bacterial concentrations than untreated samples by 1.65 Log10 CFU/mL (Fig. 1A, p = 0.002) and 1.39 Log10 CFU/mL (Fig. 1B, p = 0.02) after incubation at room temperature for 24 and 48 h, respectively. While differences were numerically smaller compared to samples incubated at room temperature, the concentrations of Salmonella Typhimurium in SJ2-treated ground pork were also significantly lower than those in untreated samples by 0.29 Log10 CFU/mL (Fig. 1C, p = 0.03) and 0.43 Log10 CFU/mL (Fig. 1D, p = 0.04) after incubation at 4°C for 24 h and 1 week, respectively. There were no differences in the concentrations of Salmonella Typhimurium in ground pork recovered after incubation at 4°C for 2 weeks between phage-treated and untreated samples (Fig. 1E, p = 0.84).
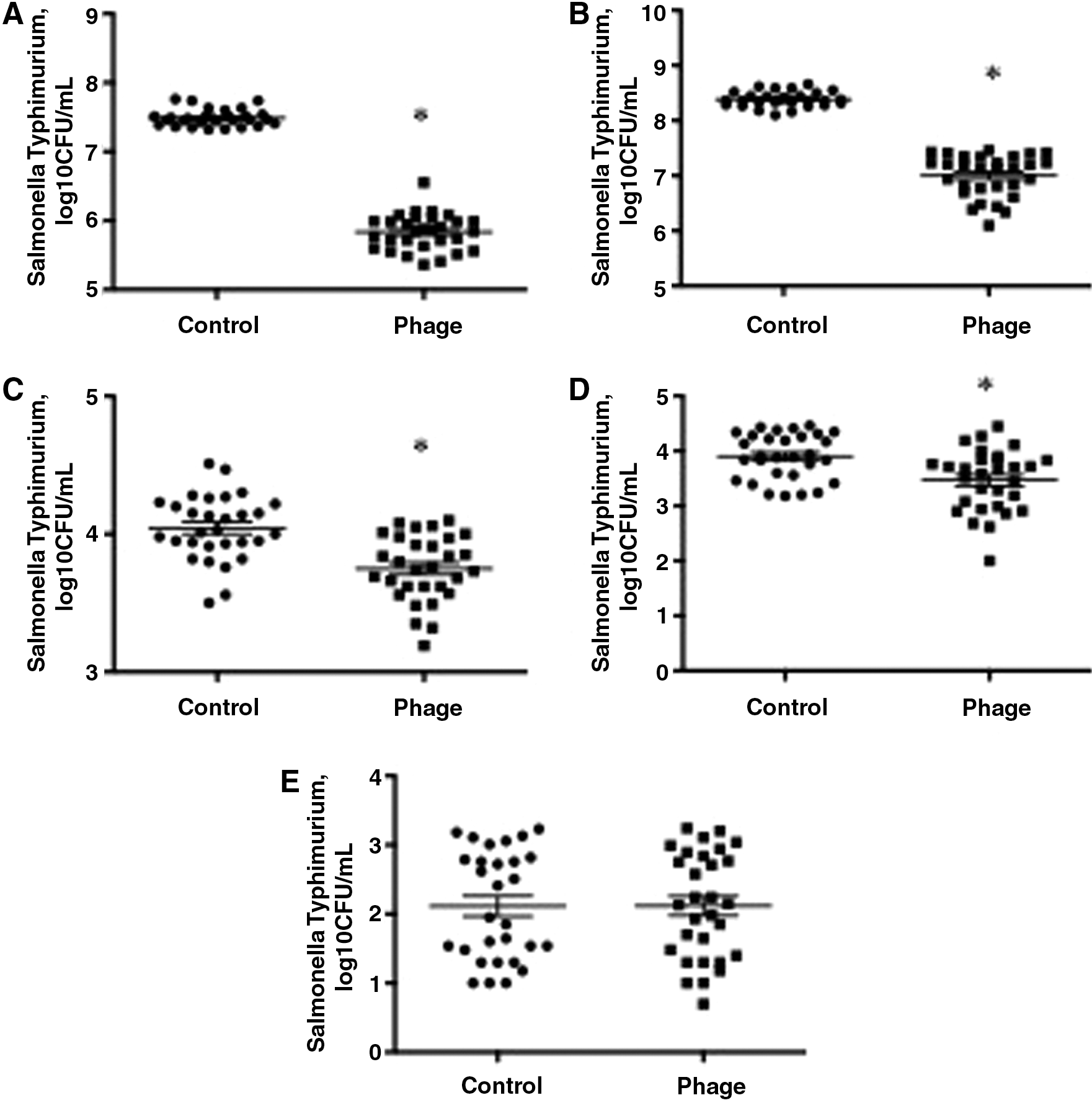
Reduction of Salmonella Typhimurium contamination in phage-treated ground pork at RT after 24 h
Similarly, in Salmonella Enteritidis-contaminated ground pork, samples that were treated with SJ2 had significantly lower concentrations of Salmonella Enteritidis than untreated samples by 1.04 Log10 CFU/mL (Fig. 2A, p = 0.009) and 1.30 Log10 CFU/mL (Fig. 2B, p = 0.002) after incubation at room temperature for 24 or 48 h, respectively. Concentrations of Salmonella Enteritidis were lower in samples treated with SJ2 than in untreated samples by 0.25 Log10 CFU/mL (Fig. 2C, p = 0.02) and 0.64 Log10 CFU/mL (Fig. 2D, p = 0.02) after 24 and 48 h, respectively, when incubated at 4°C. There were no differences in the concentrations of Salmonella Enteritidis in ground pork between phage-treated and untreated samples after incubation at 4°C for 2 weeks (Fig. 2E, p = 0.52).

Reduction of Salmonella Enteritidis contamination in phage-treated ground pork at RT after 24 h
Phage treatment of Salmonella-contaminated liquid egg
In Salmonella Typhimurium-contaminated liquid eggs, the concentrations of Salmonella Typhimurium in samples treated with SJ2 were significantly lower than those in control samples by 3.14 Log10 CFU/mL (Fig. 3A, p = 0.001) and 0.49 Log10 CFU/mL (Fig. 3B, p = 0.02) after incubation at room temperature for 24 and 48 h, respectively. Under refrigeration, the concentrations of Salmonella Typhimurium in SJ2-treated samples were lower than control samples by 0.61 Log10 CFU/mL (Fig. 3C, p = 0.008) and 0.51 Log10 CFU/mL (Fig. 3D, p = 0.02) after incubation for 24 h and 1 week, respectively. No differences were observed between phage-treated and untreated samples after incubation at 4°C for 2 weeks (Fig. 3E, p = 0.82).
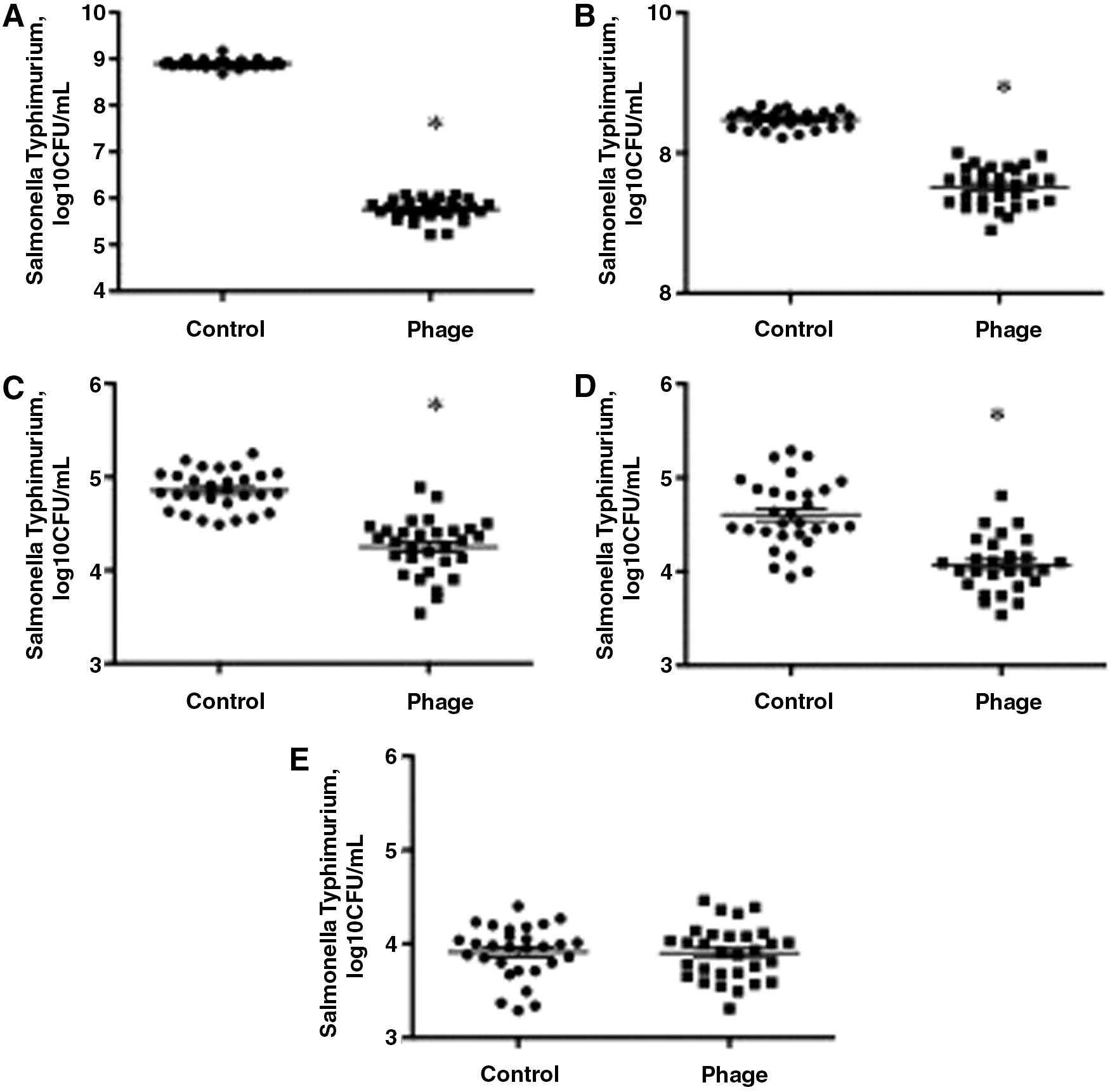
Reduction of Salmonella Typhimurium contamination in phage-treated liquid egg at RT after 24 h
In Salmonella Enteritidis-contaminated liquid egg, the concentrations of Salmonella Enteritidis in SJ2-treated samples were significantly lower than those in control samples by 0.93 Log10 CFU/mL (Fig. 4A, p = 0.003) and 0.62 Log10 CFU/mL (Fig. 4B, p = 0.046) after incubation at room temperature for 24 and 48 h, respectively. There were no differences in the concentrations of Salmonella Enteritidis between phage-treated and untreated samples after incubation at 4°C for 24 h (Fig. 4C, p = 0.052), 1 week (Fig. 4D, p = 0.054), or 2 weeks (Fig. 4E, p = 0.24).
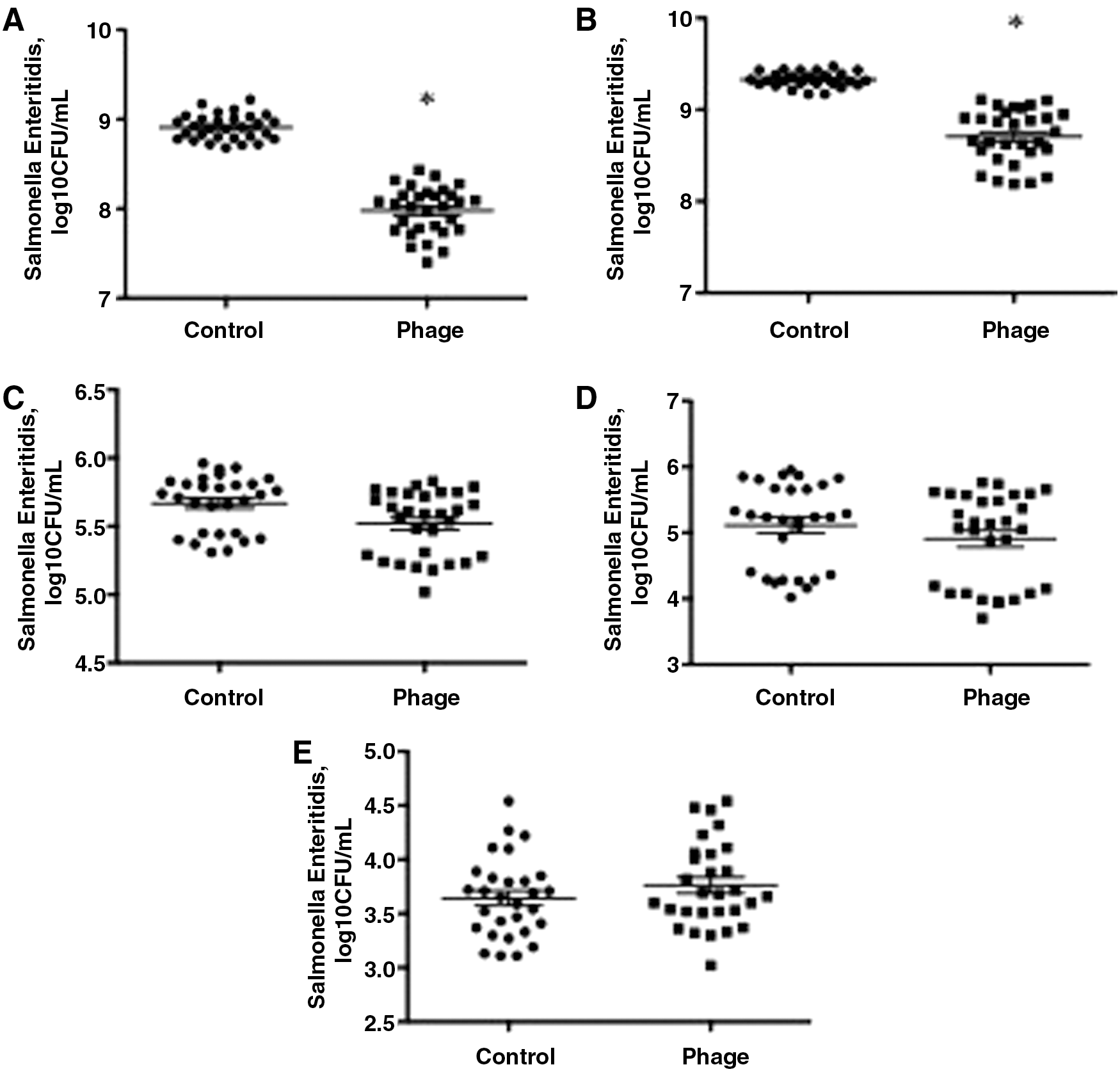
Reduction of Salmonella Enteritidis contamination in phage-treated liquid egg at RT after 24 h
Isolation of phage-resistant Salmonella
By using nonparametric analysis, the percentage of phage-resistant Salmonella Typhimurium isolates recovered from liquid eggs treated with SJ2 at room temperature was marginally higher than in untreated eggs after incubation for 24 h (untransformed means of 14.44% vs. 0.56%, p = 0.08, Fig. 5A) and significantly higher after 48 h (untransformed means of 92.50% vs. 0.56%, p = 0.007, Fig. 5B). There were no differences in the percentage of phage-resistant Salmonella Enteritidis recovered from liquid eggs treated with SJ2 compared to untreated eggs after incubation at room temperature for 24 h (untransformed means of 3.75% vs. 1.11%, p = 0.20, Fig. 5C). At 48 h, however, the percentage of phage-resistant Salmonella Enteritidis recovered from eggs treated with SJ2 was significantly higher than in untreated eggs (untransformed means of 50.83% vs. 0.56%, p = 0.03, Fig. 5D).
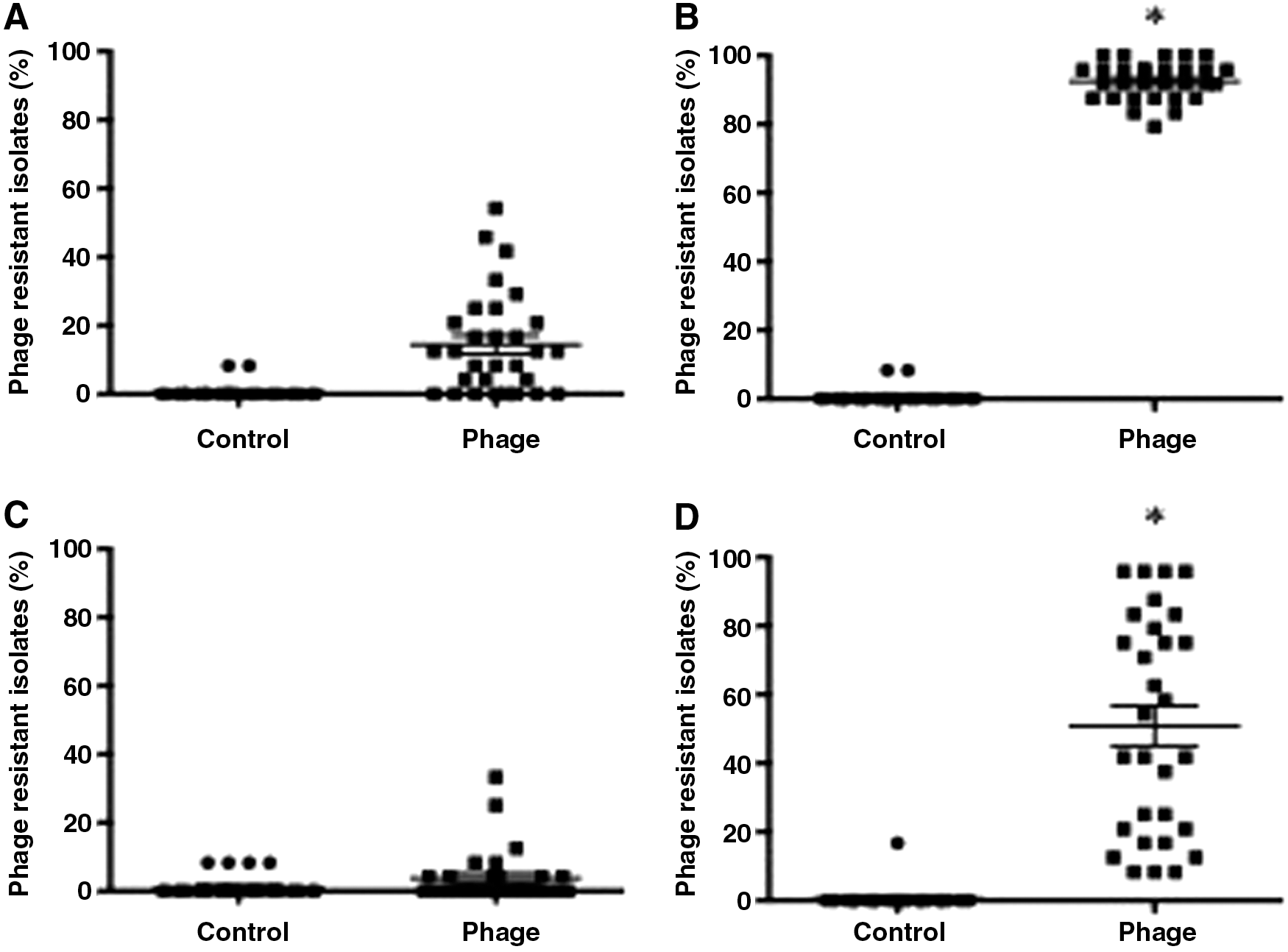
Percentage of phage-resistant Salmonella Typhimurium isolates recovered from phage-treated liquid eggs after incubation at room temperature for 24 h
In contrast, no differences were observed in the percentages of phage-resistant Salmonella Typhimurium or Salmonella Enteritidis isolated from pork samples treated with phage and untreated samples at any time point or temperature (Fig. 6A–D).
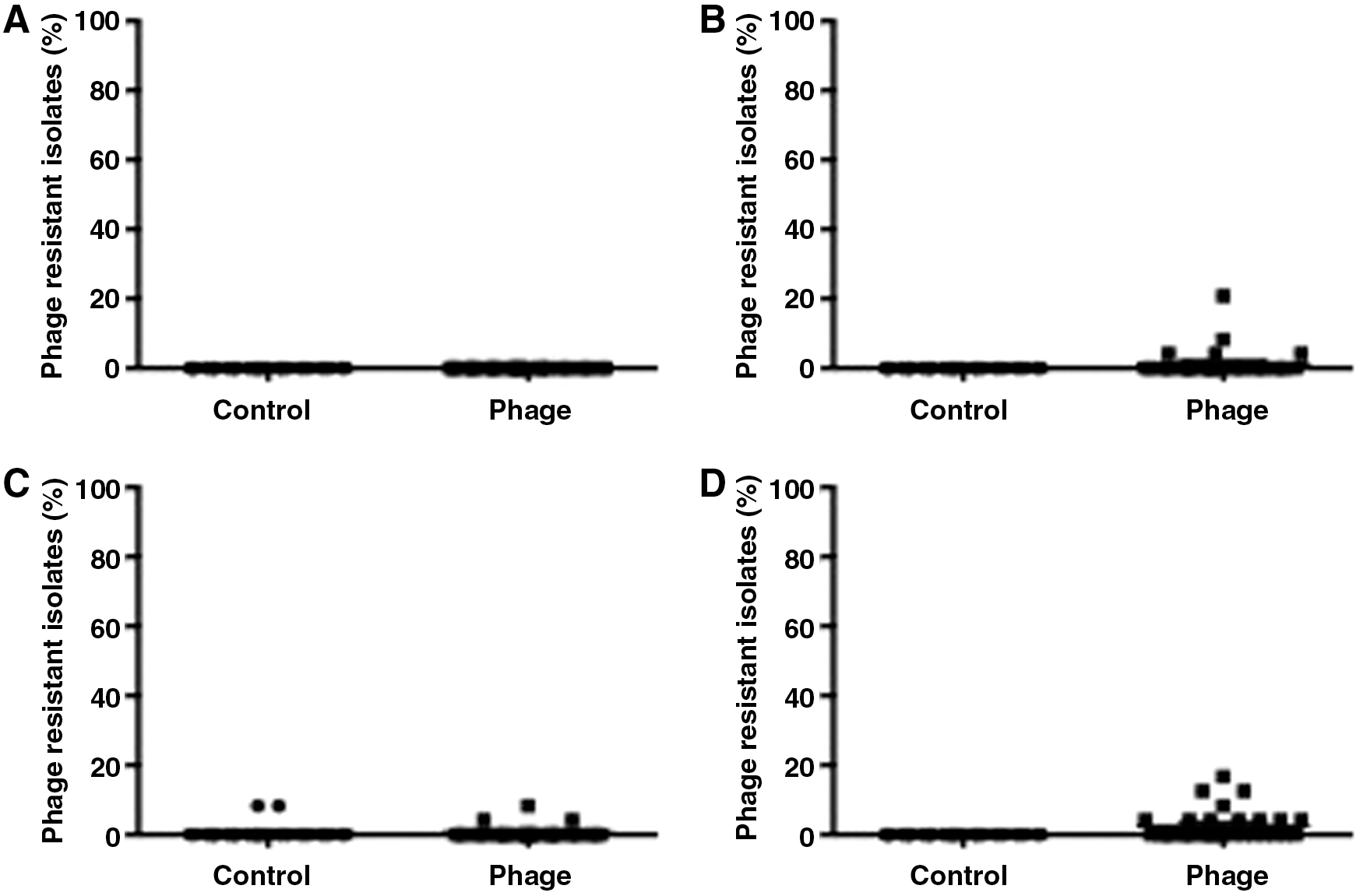
Percentage of phage-resistant Salmonella Typhimurium isolates recovered from phage-treated ground pork after incubation at room temperature for 24 h
Discussion
Phage-based antibacterial methods have received much attention in recent years due to some of their inherent qualities (e.g., ease of isolation, self-limiting nature) as well as their ability to kill antibiotic-resistant bacteria (Thiel 2004). However, in practice, one of the distinct advantages of phages, the high target specificity, may limit efficacy of phages for certain treatments (Loc-Carrillo et al., 2011), such as the applications in foods, where the exact species or strain of bacteria is often not known. Several groups have used phage cocktails (Leverentz et al., 2001, 2003; Hungaro et al., 2013; Spricigo et al., 2013; Hong et al., 2014) and broad-spectrum phages (Carlton et al., 2005; Guenther et al., 2009, 2012) in an attempt to increase application flexibility. While using phage cocktails can easily expand the target spectrum, it is not without its own set of challenges (Chan et al., 2013). As seen previously, phages in cocktails can function synergistically or antagonistically (Hall et al., 2012); therefore, to optimize the effect of a phage cocktail, phages must be carefully selected and highly characterized to avoid deleterious interactions within the cocktail itself. Comprehensive characterization of individual phages, however, is complicated by the fact that most published phage genomes contain very large sections of genes with unknown functions. For example, 73.6% of the predicted genes in phage SJ2 genome do not match any records with known functions in the NCBI database (Zhang et al., 2015). Similar published phage genomes contain comparative percentages of unknown and unannotated sequences (Kropinski et al., 2013; Hong et al., 2014; Zhang et al., 2014). This is concerning as unknown sections can include undesirable elements such as genes involved in lysogeny or virulence. Thus, it may be more time-efficient to use a single very well-characterized phage if challenges associated with resistance development are easily overcome. Regardless, more effort is likely needed in completely characterizing this and all phage genomes to more thoroughly assess their ability to transfer undesirable traits to other bacteria.
While the balance of achieving a relatively broad-spectrum phage and maintaining simplicity in the phage product may be found in the use of a single broad-spectrum phage, the efficacy of such application in foods is currently understudied. While nearly 2600 Salmonella serovars are known, Salmonella Typhimurium and Salmonella Enteritidis are the predominant serovars involved in human cases of salmonellosis both in the United States and Europe (Guenther et al., 2012) and were therefore selected in this study. Foods of animal origin, such as meat, poultry, and eggs, are major vehicles of Salmonella.
Usually, much effort is focused on strictly controlling temperatures during processing (FDA, 2005). While the efficacy of using a single broad-spectrum anti-Salmonella phage was tested under refrigeration to resemble conditions during food storage and transportation, many foodborne illness outbreaks arise from temperature and time violations at the retail or consumer level. As such, we were also interested, from a food safety perspective, in the impact of temperature violations and, from a biological standpoint, the impact of more extreme conditions and changes over time. As expected, reductions of Salmonella were less effective under refrigeration. The reduction in efficacy was seen in both Salmonella serovars and in both food matrices. Similar results were observed in our previous research focused on reducing E. coli O157:H7 in ground beef, where treatment with phages reduced concentrations of E. coli O157:H7 by 0.48 log CFU/mL when incubated at 4°C after 24 h, while concentrations were reduced by 1.97 log CFU/mL when incubated at room temperature (Hong et al., 2014). Phages are obligate parasites; therefore, temperatures closer to the optimal temperature of the host likely favor higher phage activity. Under lower than optimal temperature, the penetration of phage genetic materials into host cells is inhibited, leading to the lower lytic activity of the phages (Jończyk et al., 2011).
While a previous study indicated that phages were more effective under refrigeration (Sharma et al., 2009), it should be noted that a higher multiplicities of infection (MOI) (MOI = 100) was used in those experiments, allowing phages to lyse or at least adhere to bacteria in greater concentrations during the first few minutes of treatment. Lower MOI (virus:host ratio in viral infections), however, do not favor lysis from without, and lysis would likely be more dependent on actual phage replication, which would be stunted under refrigeration. Therefore, samples infected with lower concentrations of phages, similar to our experiment, may require a more optimum temperature to result in appreciable cell lysis.
It is interesting to note that in most cases, treatment differences in Salmonella contamination in refrigerated samples disappeared at 2 weeks. It may be that this long incubation time allowed regrowth of the targeted bacteria. Bacteria:phage concentrations are interdependent as phages are obligate parasites. As bacterial concentrations are reduced, phage concentrations may be reduced. Once threshold concentrations are reached, bacterial populations may rebound similar to the Von Magnus effect seen in defective interfering virus particles (Ebner et al., 2008).
A high concentration of Salmonella inoculum (∼106 CFU/g in ground pork and 106 CFU/mL in liquid egg) was used to prevent bacterial concentrations from falling under detection levels (103 CFU/g in meat and 10 CFU/mL in liquid egg). “Not detected,” in this case, would describe a range (e.g., between none and <102 CFU/g) in final bacterial concentrations and reduce the accuracy of most quantitative analyses. In foods in the real world, pathogens are usually found sporadically and are present in lower concentrations (e.g., <104 CFU/mL; Bigwood et al., 2009) leading to a higher MOI and greater pathogen reductions as shown in a previous study (Abuladze et al., 2008). It is worth noting that, however, the reduction becomes independent of MOI when bacterial concentrations are extremely low and a sufficiently high concentration of phage results in similar reduction regardless of the resulting MOIs (Bigwood et al., 2009).
Our study, however, was also focused on quantifying the development of phage resistance in the target bacteria as the one purpose of using phage cocktails is to mitigate the development of phage resistance (Hong et al., 2014). In this study, we aimed to determine how often and how soon targeted bacteria become resistant to a single phage within a food matrix.
Interestingly, our results showed that the emergence of phage-resistant bacteria varied between the two different food matrixes. After phage treatment at room temperature for 2 days, significantly higher levels of phage-resistant Salmonella isolates were detected in liquid egg samples, but not in ground pork samples. Similar results were observed for both Salmonella Typhimurium and Salmonella Enteritidis, indicating that the food matrix may influence the development of phage-resistant Salmonella isolates. The presence of other bacteria may influence the development of phage resistance. The reduction of Salmonella by phage provides surrounding bacteria an opportunity to grow, which in turn results in growth competition with phage-resistant Salmonella strains. Due to the nature of the food and processing method, ground pork likely harbors a more diverse bacterial community than liquid eggs. While all samples, both pork and eggs, were screened for the presence of Salmonella before the study, ground pork can have a total bacterial load of 5.6 log CFU/g (Duffy et al., 2013), whereas internal egg components are often functionally sterile, unless the hen has infected reproductive organs or bacteria translocate from outside the shell (Gantois et al., 2009). Therefore, phage-resistant Salmonella strains that emerge in ground pork may be subject to stronger growth competition. High levels (10 out of 10 tested colonies per food sample) of phage-resistant isolates were also recovered from egg yolk and chocolate milk after treatment with a single Salmonella phage (treated samples were incubated at 15°C for 6 days). The same study also showed that the emergence of phage-resistant isolates might not compromise the initial phage efficacy when initial high MOIs result in significant reduction, at least as observed in chocolate milk (Guenther et al., 2012).
However, in the present study we tested up to 720 isolates per testing condition (time × incubation temperature) per challenge strain per food matrix. Thus, our study looked at more than 10,000 isolates. While not statistically significant, marginally higher percentages of phage-resistant Salmonella Typhimurium isolates were also detected in phage-treated liquid egg samples after 1 day at room temperature, suggesting that development of phage resistance may be rapid in certain foods. In addition, the percentages of phage-resistant isolates were not different between phage-treated and untreated samples under refrigerated conditions at any time point. This is understandable as low temperature likely inhibited the physical activity of both the host bacteria and the phage as previously mentioned.
Collectively, this study demonstrated the potential of using a single broad-spectrum phage, SJ2, in reducing Salmonella Typhimurium and Salmonella Enteritidis contamination in ground pork and liquid egg. In future studies, the efficacy of SJ2 should also be measured at lower bacterial conditions, which more closely resemble the characteristics of the foods in their storage environment. It would also be of interest to measure efficacy when temperatures are more dynamic. For instance, applying the phage during processing, when temperatures are more strictly controlled, to prepare for potential temperature violations at consumer levels.
In addition, phage resistance can develop rapidly in treated food, but the level of resistance depends, at least in part, on the characteristics of the food. While the significantly higher levels of phage-resistant Salmonella isolates in phage-treated liquid egg samples at room temperature indicate that liquid egg is not an ideal food matrix for phage treatment, it would still be of interest to see whether similar high percentages of phage resistance occur at high MOI settings and to quantify the potential impact of resistance on subsequent phage applications.
Footnotes
Disclosure Statement
No competing financial interests exist.