
Abstract
Background:
Ceftriaxone resistance in Salmonella is a serious public health threat. Ceftriaxone is commonly used to treat severe Salmonella infections, especially in children. Identifying the sources and drivers of ceftriaxone resistance among nontyphoidal Salmonella is crucial.
Materials and Methods:
The National Antimicrobial Resistance Monitoring System (NARMS) tracks antimicrobial resistance in foodborne and other enteric bacteria from humans, retail meats, and food animals. We examined NARMS data reported during 1996–2013 to characterize ceftriaxone-resistant Salmonella infections in humans. We used Spearman rank correlation to examine the relationships between the annual percentage of ceftriaxone resistance among Salmonella isolates from humans with isolates from retail meats and food animals.
Results:
A total of 978 (2.9%) of 34,100 nontyphoidal Salmonella isolates from humans were resistant to ceftriaxone. Many (40%) ceftriaxone-resistant isolates were from children younger than 18 years. Most ceftriaxone-resistant isolates were one of three serotypes: Newport (40%), Typhimurium (26%), or Heidelberg (12%). All were resistant to other antimicrobials, and resistance varied by serotype. We found statistically significant correlations in ceftriaxone resistance between human and ground beef Newport isolates (r = 0.83), between human and cattle Typhimurium isolates (r = 0.57), between human and chicken Heidelberg isolates (r = 0.65), and between human and turkey Heidelberg isolates (r = 0.67).
Conclusions:
Ceftriaxone resistance among Salmonella Newport, Typhimurium, and Heidelberg isolates from humans strongly correlates with ceftriaxone resistance in isolates from ground beef, cattle, and poultry, respectively. These findings support other lines of evidence that food animals are important reservoirs of ceftriaxone-resistant Salmonella that cause human illness in the United States.
Introduction
A
Antimicrobials select for resistance wherever they are used—whether in human or veterinary medicine. Many antimicrobials used in food animals are identical or similar to those used in human medicine, and their inappropriate use can create a selective pressure for the development of resistant strains that may be transferred to people via food or through direct contact with animals. The National Antimicrobial Resistance Monitoring System (NARMS), a collaboration among the U.S. Food and Drug Administration (FDA), the Centers for Disease Control and Prevention (CDC), and the U.S. Department of Agriculture (USDA), was launched to monitor antimicrobial resistance in foodborne and other enteric bacteria.
Antimicrobial use in animal agriculture is a suspected contributor of ceftriaxone-resistant nontyphoidal Salmonella in humans; however, it is difficult to establish a definitive link between a specific use and a specific infection. We used NARMS data to further examine this association, by assessing correlations between the annual percentage of ceftriaxone resistance among nontyphoidal Salmonella isolates from humans, retail meats, and food animals. Correlations, while not proving causation, can help us understand the potential contribution of animal sources to human infections with ceftriaxone-resistant Salmonella.
Materials and Methods
Surveillance of human infections
We examined data on Salmonella infections in humans from two CDC surveillance systems: NARMS and the Foodborne Diseases Active Surveillance Network (FoodNet). We looked at NARMS data from 1996 to 2013. Clinical laboratories routinely send all Salmonella isolates to their state or local public health department for serotype determination. Public health laboratories forward isolates from sporadic cases and outbreaks of illness to the NARMS laboratory at CDC for antimicrobial susceptibility testing. In 1996, 13 states participated in NARMS, and by 2003 all 50 states participated. During 1996–2002, participating state public health laboratories forwarded every 10th isolate of nontyphoidal Salmonella to CDC; during 2003–2013, they forwarded every 20th isolate. Information including patient demographics, specimen source (e.g., blood, stool), and specimen collection date was submitted with isolates.
We also examined FoodNet data to collect more detailed information about patient demographics, clinical outcomes including hospitalization status and death, and history of recent international travel. FoodNet conducts active, population-based surveillance for laboratory-confirmed human infections with Salmonella and other pathogens commonly transmitted through food. FoodNet operates in 10 surveillance areas, which include ∼15% of the U.S. population (estimated 48 million). For the subset of NARMS isolates from FoodNet surveillance areas during 2004–2013, we linked FoodNet patient information with NARMS testing results by state laboratory identification number. From this point on, we refer to nontyphoidal Salmonella isolates included in the analysis as Salmonella.
Surveillance of retail meats
NARMS began retail meat sampling in 2002 in selected sites in Connecticut, Georgia, Maryland, Minnesota, Oregon, and Tennessee. By 2004, California, Colorado, New Mexico, and New York were added and Pennsylvania joined in 2008. Each month, personnel at each site purchased ∼40 packages of meat from grocery stores, 10 each of chicken (chicken breasts, wings, or thighs), ground turkey, ground beef, and pork chops. During 2002–2004, NARMS used convenience sampling of grocery stores but changed to a random selection of grocery stores beginning in 2005. Sites cultured the retail meats for Salmonella and forwarded isolates to the Center for Veterinary Medicine at FDA for serotype confirmation and antimicrobial susceptibility testing. Maryland did not collect retail meat samples in 2007.
Surveillance of food animals at slaughter
In 1997, NARMS began antimicrobial susceptibility testing of Salmonella isolates from chicken, turkeys, cattle, and swine. As part of the Pathogen Reduction/Hazard Analysis and Critical Control Point (HACCP) Salmonella verification testing program, the USDA Food Safety and Inspection Service (FSIS) cultured carcass rinsates (chicken), carcass swabs (turkeys, cattle, and swine), and raw ground products (chicken, turkey, and beef) collected from federally inspected slaughter and processing plants (U.S. Department of Agriculture et al., 2008).
From 1997 through June 2006, FSIS collected most of the samples at establishments selected at random, with additional samples collected from establishments that did not meet HACCP compliance standards. In mid-2006, the sampling of establishments changed to risk-based criteria, to focus resources on establishments with the most samples yielding Salmonella and the greatest number of samples with serotypes most frequently associated with human salmonellosis (U.S. Food and Drug Administration, 2014). FSIS forwarded isolates to the USDA Agricultural Research Service for antimicrobial susceptibility testing (U.S. Food and Drug Administration, 2014).
Laboratory methods
The NARMS laboratories at CDC, FDA, and USDA tested the Salmonella isolates for susceptibility to the following antimicrobials: amikacin, ampicillin, amoxicillin-clavulanic acid, cefoxitin (beginning in 2000), ceftiofur, ceftriaxone, chloramphenicol, ciprofloxacin, gentamicin, kanamycin, nalidixic acid, streptomycin, sulfonamides (sulfamethoxazole or sulfisoxazole), tetracycline, and trimethoprim-sulfamethoxazole. In 2011, azithromycin replaced amikacin on the test panel. Minimal inhibitory concentrations (MIC) were determined by broth microdilution (Sensititre®; Trek Diagnostic Systems, Oakwood Village, Ohio). The results were interpreted according to standards from the Clinical and Laboratory Standards Institute (CLSI), where established, including the current interpretive criterion for ceftriaxone resistance, defined as MIC ≥4 μg/mL (CLSI, 2014). CLSI does not have ceftiofur, streptomycin, or azithromycin interpretive criteria for Salmonella isolated from humans, so resistance breakpoints developed by NARMS were used (MIC ≥8, ≥64, and ≥32 μg/mL, respectively) (Centers for Disease Control and Prevention, 2013).
Statistical analysis
We categorized Salmonella isolates as ceftriaxone-resistant or nonresistant to ceftriaxone. The nonresistant group included ceftriaxone-susceptible isolates and those with an intermediate interpretation for ceftriaxone. We calculated odds ratios and 95% exact confidence intervals (CI) for categorical variables comparing infections caused by Salmonella resistant to ceftriaxone and nonresistant to ceftriaxone. We calculated CI around point estimates of annual percentage resistance by serotype and source, using methods described by Agresti and Coull (1998).
We used Spearman rank correlation (r) to examine the relationships between the annual percentage of ceftriaxone resistance among Salmonella isolates from humans and that among isolates from retail meats and food animals. We calculated exact p-values to determine statistical significance. Correlation analyses were performed for each of the three most common serotypes causing ceftriaxone-resistant Salmonella infections in humans and the specific food animals and animal-derived foods with which those serotypes have been linked through outbreak investigations or epidemiological studies–Newport in ground beef and cattle (Gupta et al., 2003; Varma et al., 2006), Typhimurium in retail chicken, ground beef, chicken, and cattle (Fey et al., 2000; White et al., 2001; Folster et al., 2014; Grass et al., 2014), and Heidelberg in retail chicken, ground turkey, chicken, and turkeys (Dutil et al., 2010; Folster et al., 2010, 2012; Centers for Disease Control and Prevention, 2013). Statistical analyses were conducted using SAS software, version 9.3 (SAS Institute, Cary, NC).
Results
Characteristics of human infections caused by ceftriaxone-resistant nontyphoidal Salmonella
During 1996–2013, the NARMS laboratory at CDC tested 34,100 Salmonella clinical isolates from NARMS-participating public health laboratories for antimicrobial susceptibility. Of these, 978 (2.9%) were resistant to ceftriaxone. The annual percentage of ceftriaxone resistance among Salmonella isolates from humans increased from 0.2% in 1996 to 2.5% in 2013, peaking at 4.4% in 2002–2003 (Fig. 1). Ceftriaxone resistance was detected in 44 Salmonella serotypes; most (78%) of ceftriaxone-resistant Salmonella isolates were one of 3 serotypes: Newport (393, 40%), Typhimurium (253, 26%), or Heidelberg (102, 12%). No other serotype comprised more than 4% of ceftriaxone-resistant isolates. The number and proportion of isolates resistant to ceftriaxone varied by serotype; for example, Newport was the serotype with the largest number of ceftriaxone-resistant isolates, 393 of 3388 (11.6%), while Concord was the serotype with the highest percentage of isolates resistant to ceftriaxone, 7 of 10 (70%) (Table 1).
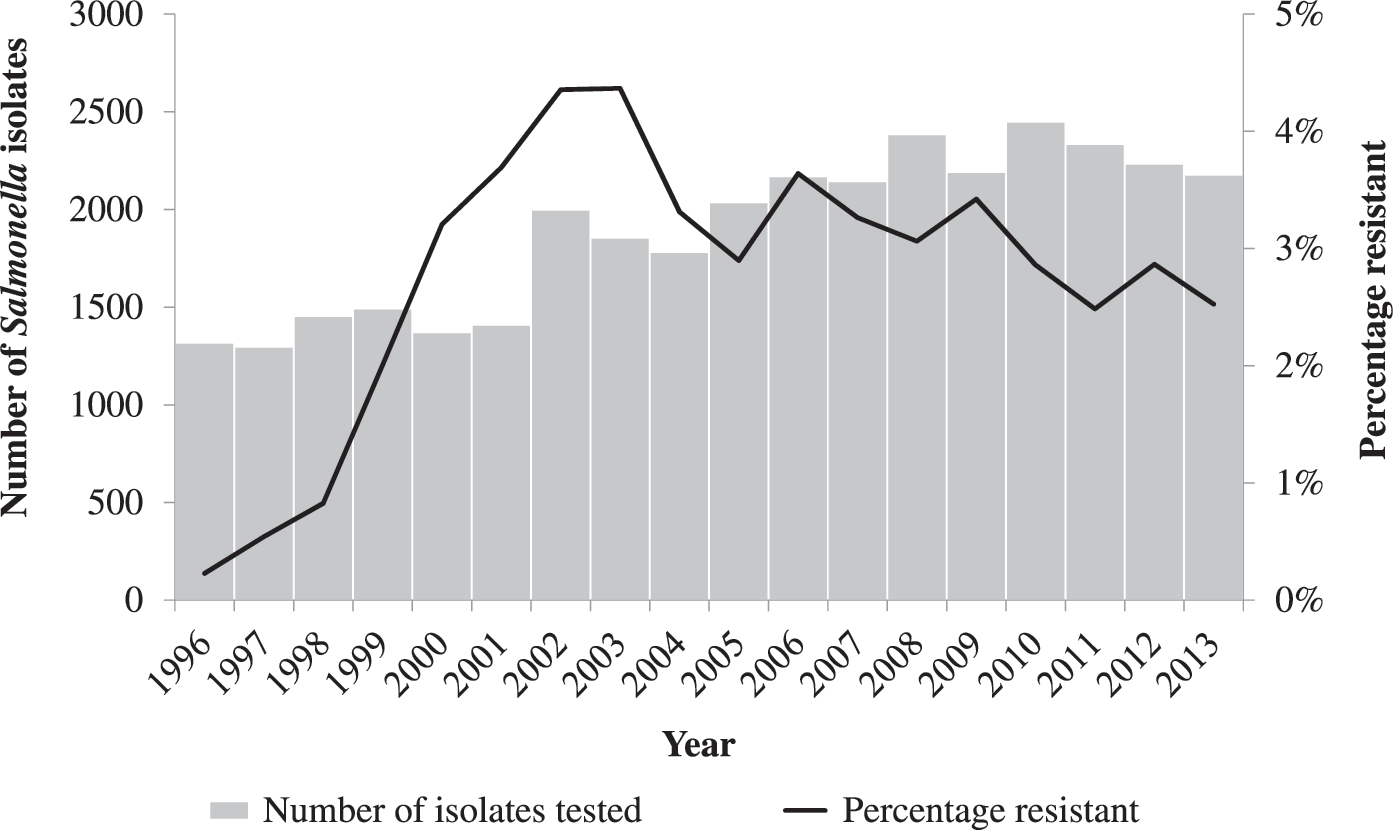
Number of nontyphoidal Salmonella isolates from humans tested and percentage resistant to ceftriaxone, by year, 1996–2013.
Other nontyphoidal Salmonella serotypes with isolates resistant to ceftriaxone were Mbandaka (3), Montevideo (3), Uganda (3), Albert (2), Bredeney (2), Cubana (2), Derby (2), Ealing (2), Ohio (2), Paratyphi B. var. L (+) tartrate + (2), Thompson (2), I 4,[5],12:-:1,2 (1), IV 44:z4,z23:- (1), Adelaide (1), Albany (1), Anatum (1), Blockley (1), Chester (1), Choleraesuis (1), Edinburg (1), Haifa (1), Hato (1), Lindenburg (1), Matopeni (1), Muenster (1), Poona (1), Stanley (1), Worthington (1).
NARMS, National Antimicrobial Resistance Monitoring System.
The 978 ceftriaxone-resistant isolates all harbored resistance to other antimicrobial agents. Resistance to other antimicrobial agents among the ceftriaxone-resistant isolates varied among serotypes (Table 2). Among Newport isolates, resistance to ampicillin, chloramphenicol streptomycin, sulfonamides, tetracycline, amoxicillin-clavulanic acid, and ceftriaxone (resistance-type ACSSuTAuCx) was high (377 of 393 ceftriaxone-resistant isolates, 95.9%). This resistance pattern was less common in ceftriaxone-resistant Typhimurium isolates (138 of 253, 54.5%), and much less common (6 of 120 isolates, 5.0%) in ceftriaxone-resistant Heidelberg isolates. Altogether, 57 (5.8%) ceftriaxone-resistant Salmonella isolates showed decreased susceptibility or resistance to ciprofloxacin (MIC ≥0.12 μg/mL).
Includes four isolates with partial or unknown serotype.
Sulfamethoxazole was replaced by sulfisoxazole in 2004.
β-lactam agents include amoxicillin-clavulanic acid, ampicillin, ceftiofur, and cefoxitin, in addition to ceftriaxone.
AAuCx: ampicillin, amoxicillin-clavulanic acid, ceftriaxone.
ACSSuTAuCx: ampicillin, chloramphenicol, streptomycin, sulfamethoxazole/sulfisoxazole, tetracycline, amoxicillin-clavulanic acid, ceftriaxone.
MIC, minimal inhibitory concentration.
Table 3 shows characteristics of patients with infections caused by ceftriaxone-resistant strains compared with those among patients having infections caused by strains not resistant to ceftriaxone. Among patients whose age was reported, 40% (367/922) of those with ceftriaxone-resistant Salmonella and 44% (13,407/30,386) of those with Salmonella not resistant to ceftriaxone were <18 years old. A lower proportion of infections in children 1–4 years, and conversely, a higher proportion of Salmonella infections in older adults were caused by resistant strains compared with adults 18–64 years, approaching statistical significance (Table 3). There was no statistical association between specimen source and ceftriaxone resistance (Table 3).
Odds ratio and Fisher's exact confidence intervals. Missing data were not used in computing odds ratios.
NARMS clinical isolates, 1996–2013, (n = 34,100).
Isolates in NARMS that were linked to FoodNet case reports, 2004–2013 (n = 3254).
CI, confidence interval.
During 2004–2013, 94 (2.9%) ceftriaxone-resistant Salmonella isolates were submitted among 3254 NARMS Salmonella isolates linked to FoodNet case reports. The proportion of patients hospitalized and the case fatality rate were higher in patients with ceftriaxone-resistant Salmonella than in patients with Salmonella not resistant to ceftriaxone, although these findings were not statistically significant (Table 3). Among patients with information on international travel before illness onset, 7 (9.2%) of 76 with ceftriaxone-resistant Salmonella reported international travel, compared with 224 (9.4%) of 2389 with Salmonella not resistant to ceftriaxone (Table 3). Patients with infection caused by ceftriaxone-resistant Salmonella were diagnosed in the United States after reported travel in China, Egypt, Ethiopia, Mexico, and Peru.
Correlations in resistance proportions among isolates from humans, retail meats, and food animals
During 2002–2013, the NARMS laboratory at FDA detected ceftriaxone resistance in 509 (26.2%) of 1940 Salmonella isolates from retail chicken, 167(9.0%) of 1862 isolates from ground turkey, and 21 (13.5%) of 155 isolates from ground beef. During 1997–2013, ceftriaxone resistance was detected in 1576 (9.5%) of 16,608 isolates from chicken at slaughter, 248 (5.6%) of 4457 isolates from turkeys, and 1192 (12.6%) of 9461 isolates from cattle. Estimates and CI of annual percentages of resistance to ceftriaxone in Salmonella serotypes Newport, Typhimurium, and Heidelberg isolated from humans, retail meats, and food animals are shown in Figure 2, while Table 4 shows the observed number of Salmonella isolates recovered by serotype and source and the number and percentage of isolates resistant to ceftriaxone. Supplementary Figure S1(Supplementary Data are available online at

Agresti-Coull estimates and 95% confidence intervals for percentage resistant to ceftriaxone among Salmonella ser. Newport
Isolates have been collected and tested since 1996 for humans; since 1997 for cattle, chicken, and turkeys; and since 2002 for ground beef, retail chicken, and ground turkey. Numbers in parentheses indicate total number of isolates obtained for antimicrobial resistance testing.
N/A indicates that no isolates were obtained for antimicrobial resistance testing.
Ceftriaxone resistance in Salmonella serotype Newport isolates from humans increased from 0% in 1996 to a peak of 25.4% in 2001 and then declined to 5.3% in 2013 (Fig. 2). Similarly, there was a sharp rise in ceftriaxone resistance among Newport isolates from cattle, with a peak of 81% during 2005. However, the annual percentage of ceftriaxone resistance in isolates from humans did not correlate with resistance in cattle (r = 0.34, CI = −0.14–0.81, p = 0.20). Newport was detected infrequently in retail ground beef samples, but resistance correlated strongly with that in isolates from humans (r = 0.83, CI = 0.47–1.0, p = 0.03) and from cattle (r = 0.90, CI = 0.69–1.0, p = 0.01).
Ceftriaxone resistance among Salmonella Typhimurium isolates from humans increased from 0% in 1996 to a high of 6.8% in 2011. The annual percentages of resistance were higher in cattle and chicken isolates than in human isolates. Ceftriaxone resistance in human isolates correlated strongly with that in cattle isolates (r = 0.57, CI = 0.11–1.0, p = 0.02). The correlation between resistance in human and chicken isolates was lower and was not statistically significant (r = 0.48, CI = −0.03–0.99, p = 0.05). For Salmonella Typhimurium, we found no correlations in resistance between retail meat (retail chicken or ground beef) and human isolates, or between retail meat and animal isolates.
Among Heidelberg isolates from humans, chicken, turkeys and retail poultry, ceftriaxone resistance has increased in recent years. We found significant correlations in resistance between human and chicken isolates (r = 0.65, CI = 0.60–1.0, p = 0.0004) and between human and turkey isolates (r = 0.62, CI = 0.27–1.0, p = 0.01). As with Typhimurium and Newport, relatively few Heidelberg isolates were available from retail meats, and correlations in resistance were lower and were not statistically significant between human and retail poultry isolates (retail chicken [r = 0.17, CI = −0.47–0.81, p = 0.59] and ground turkey [r = 0.24, CI = −0.37–0.85, p = 0.45]) and between retail poultry and poultry isolates (chicken sources [r = 0.40, CI = −0.11–0.91, p = 20 and turkey sources [r = 0.55, CI = 0.17–0.92, p = 0.07]).
Discussion
Ceftriaxone resistance has emerged and persisted among nontyphoidal Salmonella recovered from humans, retail meats, and food animals in the United States over the past several years, although resistance to ceftriaxone was relatively infrequent among Salmonella isolates from humans overall. NARMS data show ceftriaxone resistance among Salmonella in some retail meats and food animals is high and trends in ceftriaxone resistance among isolates from humans generally parallel those seen in food animals. We saw strong correlations between ceftriaxone resistance among Salmonella serotype Heidelberg isolates in humans and poultry at slaughter, which is consistent with other studies recognizing the connection between human Heidelberg infection and poultry at slaughter and at retail (Dutil et al., 2010; Folster et al., 2010, 2012; Hoffmann et al., 2012; Centers for Disease Control and Prevention, 2013). Similarly, the correlation we found between ceftriaxone resistance in Typhimurium isolates from humans and cattle supports previous reports of ceftriaxone-resistant Typhimurium infections associated with cattle and foods derived from them (Fey et al., 2000; Jackson et al., 2013; Folster et al., 2014; Grass et al., 2014). These correlations help support, though they do not prove, that food animals are an important source of ceftriaxone-resistant Salmonella, which may be transmitted to humans through the foods we eat.
The presence of ceftriaxone-resistant Salmonella in food animals may be a consequence of ceftiofur use in agriculture (Dunne et al., 2000; Fey et al., 2000; Zhao et al., 2001; Dutil et al., 2010). Ceftiofur is a third-generation cephalosporin-like ceftriaxone and cross-resistance between the two drugs is common (U.S. Food and Drug Administration, 2014). It is the only extended-spectrum cephalosporin approved for use in food animals in the United States (U.S. Food and Drug Administration, 2012, 2016). It was first approved for use in cattle in 1988, swine and chickens (day-old chicks) in 1992, sheep and turkeys (day-old poults) in 1996, and goats in 2001, with label indications for the treatment and control of certain diseases (U.S. Food and Drug Administration, 2014, 2016). Historically, the same molecular mechanism has been responsible for resistance to both ceftiofur and ceftriaxone in NARMS isolates (Dunne et al., 2000; Fey et al., 2000; Winokur et al., 2000; White et al., 2001; Zhao et al., 2001; Folster et al., 2011, 2012; Sjolund-Karlsson et al., 2013).
Although cattle and beef are known sources of ceftriaxone-resistant Newport infections in humans (Gupta et al., 2003; Varma et al., 2006), we detected a statistically significant correlation between Newport human and ground beef isolates only and not with cattle isolates. Additionally, we did not detect a correlation between Typhimurium human and chicken isolates, while ceftriaxone-resistant Typhimurium is commonly found in poultry (White et al., 2001; Folster et al., 2014; U.S. Food and Drug Administration, 2014).
It is difficult to draw conclusions from the lack of statistically significant correlations, because of several limitations to our data. NARMS surveillance data do not include the specific sources of the infections among humans. The sources of Salmonella Newport and Typhimurium infections are diverse and changing, and they include animal- and plant-derived foods and environmental sources (Varma et al., 2006; Greene et al., 2008; Jackson et al., 2013). We based our correlation analyses only on the prevalence of resistance captured by NARMS, that is, percentage of resistance among Salmonella isolates tested, not incidence of resistant infections in humans, frequency of isolation of resistant Salmonella from food animals and retail meats, or proportion of infections associated with food animal sources. For example, while there continue to be high levels of ceftriaxone resistance in Newport isolates from cattle, the number of Newport isolates from cattle has declined in recent years (U.S. Department of Agriculture and Food Safety Inspection Service 2011; U.S. Food and Drug Administration, 2014), concurrent with an increase in isolate submission rates of pansusceptible Newport in humans (Chai et al., 2012), suggesting that an increasing number and proportion of Newport infections in humans are attributable to non-bovine sources. Additionally, the sample sizes and periods of surveillance differed among sources; this may have resulted in comparisons that lacked power to detect statistical temporal associations, particularly with retail meats. NARMS retail meat sampling is limited to only a few surveillance areas and did not begin until 2002. Consequently, there were relatively few Salmonella isolates from retail meats, especially ground beef, available for antimicrobial susceptibility testing, resulting in wide CI around annual percentage estimates of resistance.
Our analysis was subject to limitations beyond those discussed above. NARMS sampling methods may have introduced biases in our study. NARMS monitoring of human infections was not national until 2003. Additionally, human isolates surveillance is passive, with possible biases in sampling because of differences in diagnostic strategies employed by clinicians and state-to-state variations in isolate submission to public health laboratories. NARMS isolates from food animals originated from USDA in-plant HACCP monitoring. When this sampling program was changed in 2006, more samples likely came from establishments that were out of compliance, rather than from a random selection of slaughter establishments. The potential biases that this sampling strategy introduced are unknown.
Our findings of correlations in ceftriaxone resistance between Salmonella isolates from humans and food animals contribute to the growing body of evidence that food animals are an important source of ceftriaxone-resistant Salmonella causing human infections, although they do not establish causality. Nevertheless, to reduce the transmission of resistant nontyphoidal Salmonella to humans, it is essential to decrease the prevalence of resistant Salmonella in food products and food animals. Our findings underscore the need to reduce unnecessary uses of extended-spectrum cephalosporins in animals and in humans and to find alternatives to their use, which will require multidisciplinary efforts by veterinarians, the agricultural industry, clinicians, and public health agencies. To help address the problem FDA issued an order prohibiting certain extralabel uses of cephalosporins (excluding cephapirin) in cattle, swine, chicken, and turkeys in 2012 (U.S. Food and Drug Administration, 2012). In Canada, a voluntary withdrawal of ceftiofur for in ovo use in hatcheries preceded a marked decline in ceftiofur-resistant Salmonella serotype Heidelberg in retail chicken and in humans, demonstrating that the management of ceftiofur use in hatcheries could be an effective control strategy (Dutil et al., 2010). Additionally, FDA issued guidance recommending that antibiotics important for human health be limited to uses in food animals that are necessary to assure animal health and involve veterinary oversight or consultation (U.S. Food and Drug Administration and Center for Veterinary Medicine, 2012, 2012). Continued national surveillance through NARMS will be important to evaluate the impact of regulatory actions and antimicrobial stewardship efforts and to direct additional efforts aimed at preserving the efficacy of extended-spectrum cephalosporins for the treatment of human infections.
Footnotes
Acknowledgments
We are indebted to the local and state health departments and laboratories that participate in the National Antimicrobial Resistance Monitoring System. We thank Jennifer Huang for her assistance in gathering the data, and Patricia Griffin and Patrick McDermott for their helpful review of this report.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.