
Abstract
The aim of the present study was to determine biocide tolerance and antibiotic resistance in Salmonella isolates from hen eggshells. A total of 39 isolates from hen eggshells, identified as either Salmonella spp. or Salmonella enterica according to 16S rDNA sequencing, were selected for biocide tolerance. Isolates with minimum inhibitory concentrations (MICs) above the wild-type MICs were considered to be biocide tolerant: benzalkonium chloride (BC, 7.7%), cetrimide (CT, 7.7%), hexadecylpyridinium chloride (HDP, 10.3%), triclosan (TC, 17.9%), hexachlorophene (CF, 30.8%), and P3-oxonia (OX, 25.6%). The resulting 21 biocide-tolerant isolates were further characterized. Most isolates (95.2%) were resistant to ampicillin, but only 9.5% were resistant to cefotaxime as well as to ceftazidime. Resistance to chloramphenicol (61.9%), tetracycline (47.6%), streptomycin (19.0%), nalidixic acid (28.6%), ciprofloxacin (9.5%), netilmicin (14.3%), and trimethoprim–sulfamethoxazole (38.1%) was also detected. Considering only antibiotics, 66.7% of isolates were multiresistant; furthermore, 90.5% were multiresistant considering antibiotics and biocides combined. Efflux pump and biocide tolerance genetic determinants detected included acrB (95.2%), oqxA (14.3%), mdfA (9.5%), qacA/B (4.8%), and qacE (9.5%). Antibiotic resistance genes detected included bla TEM (14.3%), bla CTXM-2 (4.8%), bla PSE (4.8%), floR (19.05%), tet(A) (9.5%), tet(C) (4.8%), dfrA12 (0.05%), and dfrA15 (0.05%). Significant positive correlations were detected between phenotypic tolerance/resistance to biocides, biocides and antibiotics, and also between antibiotics, suggesting that a generalized use of biocides could co-select antibiotic resistance.
Introduction
S
Biocides are frequently used in sanitation in food processing lines and surfaces in the food industry (SCENIHR, 2009). Exposure to subinhibitory concentrations of biocides, which might occur during poor disinfection and cleaning procedures, could lead to selection of strains with reduced susceptibility (Karatzas et al., 2007). The increased use of biocides has raised concerns about a possible role in selection for antibiotic-resistant bacteria (SCENIHR, 2009; Ortega Morente et al., 2013; Wales and Davies, 2015). A recent study suggested that Salmonella strains challenged by prolonged treatment with the disinfectants become resistant to antibiotics commonly used for the treatment of invasive Salmonella infections in humans such as cefotaxime and ciprofloxacin (Futoma-Kołoch et al., 2015). The purpose of the present study was to determine the frequency of biocide tolerance in Salmonella isolates from hen egg shells and to determine antibiotic resistance in the biocide-tolerant isolates.
Materials and Methods
Isolation of bacterial strains
A total of 30 egg packs (containing 12 eggs per pack) were purchased (1 egg pack per sale point per day) from four different grocery stores located in the province of Jaen and two supermarkets located in the province of Granada (both in Southern Spain), in the year 2013. Egg packs were purchased on 5 different days in a 1-month period. Egg packs were purchased at room temperature, and stored under refrigeration upon arrival to the laboratory (for not more than 24 h) until they were analyzed. All manipulations were carried out with hand gloves. A total of 120 hen eggs (including 4 eggs selected at random from each pack) were used for analysis. Each egg was placed inside a sterile Stomacher bag containing 35 mL Trypticase Soy Broth (TSB; Scharlab, Barcelona, Spain), and left at room temperature for 1 h. Then, eggs were rubbed by hand for one min. A fraction (10 mL) of the suspension recovered from egg surface was transferred to a sterile test tube and incubated at 37°C for 2 h. A loopful of the bacterial suspension recovered from egg shell surface was spread on violet red bile agar (VRBA) plates (Scharlab) and incubated at 37°C for 24 h (Musgrove et al., 2005). For each positive sample, two isolates were randomly selected from presumptive Enterobacteriaceae (dark red to purple colonies with red–purple haloes). Each selected isolate was streaked for purity onto VRBA plates and incubated at 37°C overnight. An isolate from the third streak plate was grown on TSB for 18 h, and then stored at −80°C in TSB supplemented with 20% glycerol.
Strain identification
Bacterial isolates were identified by conventional tests (Gram staining, catalase and oxidase tests) and 16S rDNA sequencing as described by Weisburg et al. (1991) and Abriouel et al. (2005).
Determination of biocide tolerance
Isolates were tested for sensitivity to the quaternary ammonium compounds (QACs) benzalkonium chloride (BC), cetrimide (CT), and hexadecylpyridinium chloride (HDP), the bisphenols triclosan (TC) and hexachlorophene (CF), the polyguanidine polyhexamethylene guanidine hydrochloride (PHMG), and the commercial sanitizer P3-oxonia (OX). BC (commercial solution containing 50% of the active compound by weight), CT, HDP, TC, and CF were from Sigma-Aldrich (Madrid, Spain). TC and CF were dissolved (10% w/v) in 96% ethanol. HDP (5%) and CT (10%) were dissolved aseptically in sterile distilled water. Biocide solutions were stored at 4°C for ≤7 days. PHMG solution (containing 7.8% of PHMG, by weight) was a kind gift of Oy Soft Protector Ltd. (Espoo, Finland). OX (25–35% hydrogen peroxide, 0.83–2.5 N acetic acid, and 0.26–0.66 N peracetic acid) was from ECOLAB (Barcelona, Spain). Minimum inhibitory concentrations (MICs) to biocides were determined by the broth microdilution method on 96-well flat bottom microtiter plates (Becton Dickinson Labware, Franklin Lakes, NJ). Briefly, serial dilutions of each biocide in TSB were inoculated (0.1%, v/v) with cultures of bacterial strains grown overnight in TSB. The inoculated broths were distributed aseptically in microtiter plates (150 μL per well) in triplicate. Growth and sterility controls were included for each isolate. Microtiter plates were incubated at 37°C for 24 h. Bacterial growth was determined spectrophotometrically (OD 595 nm) with an iMark Microplate Reader (Bio-Rad, Madrid, Spain). All assays were done in triplicate.
Determination of antibiotic resistance
Antibiotic resistance was determined by the disk diffusion method as described by the Clinical and Laboratory Standards Institute (CLSI, 2014) on cation-adjusted Mueller-Hinton agar (Fluka; Sigma-Aldrich). Ampicillin (AMP, 30 μg), ceftazidime (CFZ, 30 μg), cefotaxime (CTX, 30 μg), imipenem (IPM, 10 μg), streptomycin (SM, 10 μg), netilmicin (NET, 30 μg), tetracycline (TET, 30 μg), ciprofloxacin (CIP, 5 μg), nalidixic acid (NA, 30 μg), and trimethoprim–sulfamethoxazole (TM/STX, 1.25/23.75 μg) were from BioMérieux (Madrid, Spain). Chloramphenicol (CMP, 30 μg) was from BBL (Madrid, Spain). MICs of the antibiotic-resistant strains detected by disk diffusion tests were determined by broth microdilution according to CLSI (CLSI, 2014).
Investigation of biocide and antibiotic resistance genes
The presence of biocide and antibiotic resistance genes was investigated by polymerase chain reaction (PCR) amplification using the primers and PCR conditions described elsewhere. QAC resistance genes qacA/B was determined according to Noguchi et al. (2005), whereas qacC (smr), qacG, qacH, and qacJ were determined as described by Smith et al. (2008). The presence of qacE and qacEΔ1 genes and their association with Class I integrons was investigated as using the primer combinations and PCR conditions described by Chuanchuen et al. (2007): for amplification of qacE, qacEΔ1, and the 3′ coding sequence, forward primer qacEF was used in combination with reverse primer qacER and sulR. The integrase gene intI1 was investigated with intF and intR primers (Chuanchuen et al., 2007).
Genetic determinants for antibiotic resistance investigated by PCR included the beta-lactamase resistance genes bla TEM (Sáenz et al., 2004), bla PSE (Chiu et al., 2006), bla CTX-M, and bla CTX-M-2 (Bertrand et al., 2006); aminoglycoside resistance gene aac(6′)-Ib-cr (Park et al., 2006); and tetracycline resistance genes tet(A), tet(B), tet(C), tet(D), tet(E), and tet(G) (Ng et al., 2001). Phenicol resistance genes floR (Chiu et al., 2006) and cmlA (Guerra et al., 2001), Sulfonamide and trimethoprim resistance genes sul1, dfrA12, and dfrA15 were determined according to Guerra et al. (2001). The oqxA gene of the OqxAB multidrug efflux pump was investigated according to Hansen et al. (2005). Efflux pump genes acrB and mdfA were investigated according to Swick et al. (2011).
Statistical analysis
The relationships between biocide tolerance and antibiotic resistance were studied by Principal component analysis with Pearson correlation coefficients (r) by using IBM SPSS Statistics 22 (IBM Corporation, Armonk, NY) and MYSTAT statistics and graphics package (Systat Software, evaluation version 2015.1; Hounslow, London, UK). Positive correlations were defined as very weak (0.00–0.19), weak (0.20–0.39), moderate (0.4–0.59), strong (0.60–0.79), or very strong (0.80–0.99), with a p significances of <0.05 or 0.01.
Results
Identification of bacterial isolates and determination of biocide tolerance
A total of 39 bacterial isolates from egg shells were identified as either Salmonella spp. or S. enterica according to 16S rDNA sequencing, and selected for biocide tolerance determination. According to biocide tolerance tests, several isolates required biocide concentrations for growth inhibition that were above the wild-type MICs, and therefore were considered to be tolerant to the following biocides (Table 1): BC, 7.7%; CT, 7.7%; HDP, 10.3%; TC, 17.9%; CF, 30.8%; OX, 25.6%. None of the isolates was considered to be tolerant to PHMG. From this preliminary screening, 21 isolates were identified as being tolerant to at least one biocide.
Number of isolates for each MIC of biocide.
In milliliter of commercial solution per liter.
BC, benzalkonium chloride; CT, cetrimide; HDP, hexadecylpyridinium chloride; TC, triclosan; CF, hexachlorophene; PHMG, polyhexamethylene guanidine hydrochloride; OX, P3-oxonia; MIC, minimum inhibitory concentration.
Antimicrobial resistance of isolates
Most isolates (95.2%) were resistant to AMP, but only 9.5% were resistant to CTX as well as to CFZ (Table 2). All isolates were sensitive to IMP. The frequency of CMP resistance was high (61.9%), followed by TET (47.6%) and SM (19.0%). Resistances to NA (28.6%), NET (14.3%), TM/STX (38.1%), and CIP (9.5%) were also detected among biocide-tolerant isolates. MICs of antibiotic-resistant isolates are shown in Table 2.
Antibiotic MICs are indicated in parenthesis.
BC, benzalkonium chloride; CT, cetrimide; HDP, hexadecylpyridinium chloride; TC, triclosan; CF, hexachlorophene; OX, P3-oxonia; AMP, ampicillin; CTX, cefotaxime; CFZ, ceftazidime; CM, chloramphenicol; SM, streptomycin; TET, tetracycline; CIP, ciprofloxacin; NA, nalidixic acid; NET, netilmicin; TM/STX, trimethoprim–sulfamethoxazole; MICs, minimum inhibitory concentration.
A high percentage of isolates (66.7%) were resistant to at least three antibiotics of different classes, and 23.8% of isolates were tolerant to at least three biocides that in addition belonged to at least two different classes (Table 2). Considering biocides and antibiotics together, 90.5% of isolates were resistant/tolerant to at least three antimicrobials. Remarkably, isolates Salmonella spp. UJAS18 and UJAS19 were resistant to nine antibiotics and tolerant to five or four of the biocides tested, respectively.
Correlations between biocide tolerance and antimicrobial resistance
Results from principal component analysis of biocide tolerance and antibiotic resistance data are shown in Figure 1 and Table 3. Significant positive correlations between biocides were very strong (for CT and BC), strong (for TC and HDP), or moderate (for HDP and TC with BC and with CT). OX showed negative correlations with other biocides.
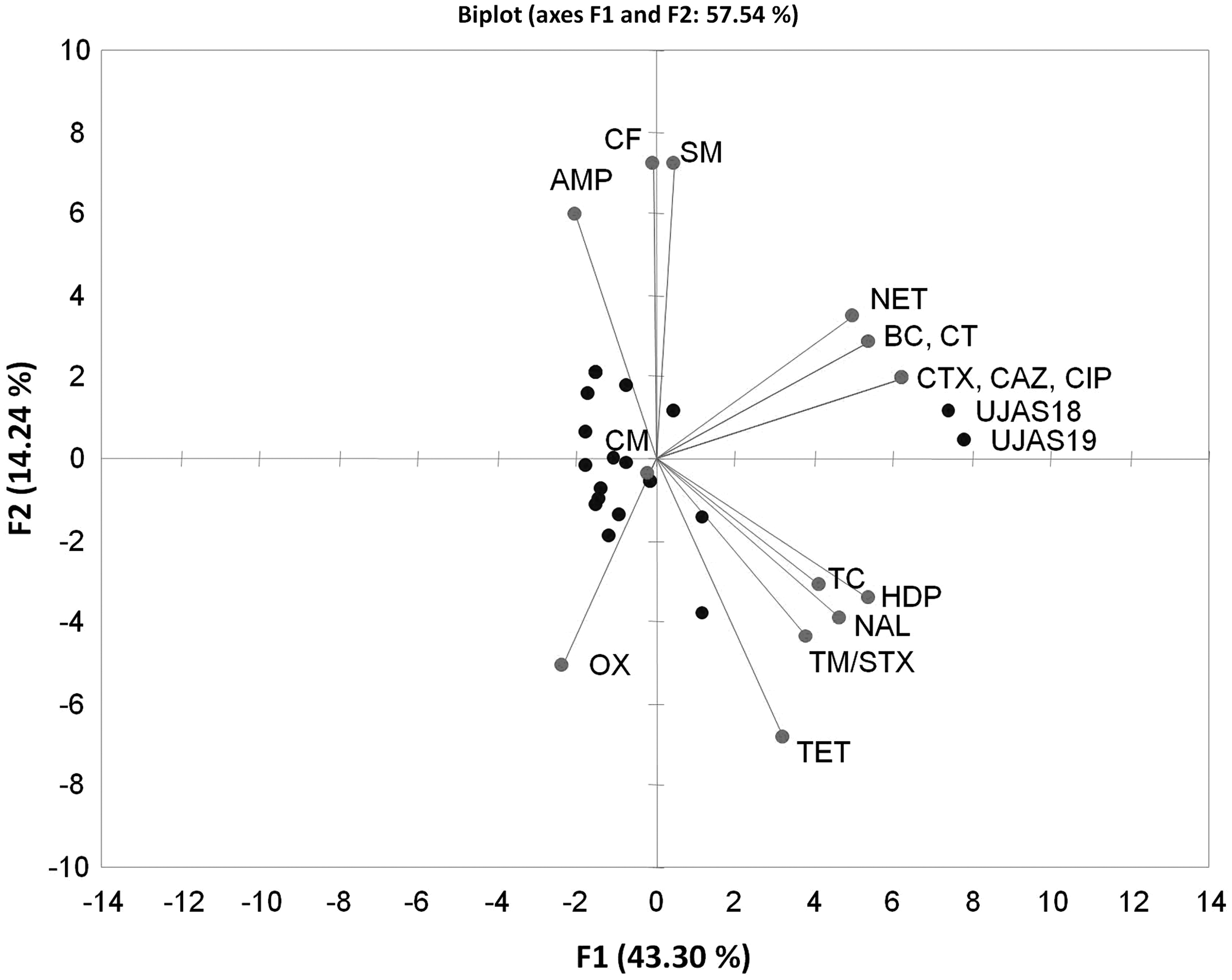
Biplot for biocide tolerance and antimicrobial resistance (red dots) in Salmonella isolates from egg shell surfaces (blue dots). The two isolates that showed greatest number of biocide and antibiotic resistance traits (UJAS18 and UJAS19) are indicated. BC, benzalkonium chloride; CT, cetrimide; HDP, hexadecylpyridinium chloride; TC, triclosan; CF, hexachlorophene; OX, P3-oxonia; AMP, ampicillin; CTX, cefotaxime; CFZ, ceftazidime; CM, chloramphenicol; SM, streptomycin; TET, tetracycline; CIP, ciprofloxacin; NAL, nalidixic acid; NET, netilmicin; TM/STX, trimethoprim–sulfamethoxazole.
Significant correlation at p < 0.01 level (two tailed).
Significant correlation at p < 0.05 level (two tailed).
BC, benzalkonium chloride; CT, cetrimide; HDP, hexadecylpyridinium chloride; TC, triclosan; CF, hexachlorophene; PHMG, polyhexamethylene guanidine hydrochloride; OX, P3-oxonia; AMP, ampicillin; CTX, cefotaxime; CFZ, ceftazidime; IMP, imipenem; SM, streptomycin; CIP, ciprofloxacin; CM, chloramphenicol; TET, tetracycline; NA, nalidixic acid; NET, netilmicin; TM/STX, trimethoprim–sulfamethoxazole.
Significant positive correlations were also detected between biocides and antibiotics. BC, CT, and HDP showed moderate or strong positive correlations with CTX, CFZ, CIP, and NET. HDP also showed moderate or strong positive correlations with TET, NA, and TM/STX. TC showed significant, moderate positive correlations with CTX, CFZ, CIP, and NA. CF was the only biocide having a significant positive correlation with AMP.
The study also revealed significant positive correlations between antibiotics that were strong or very strong for CIP with CTX and CFZ or for NET with CTX, CFZ and CIP. NA showed positive correlations with CTX, CFZ, CIP, TET, and TM/STX (Table 3). TM/STX also showed a significant positive correlation with TET.
Genetic determinants related to antimicrobial resistance
PCR amplification of efflux pump genes revealed that 95.2% of those isolated were positive for acrB of the AcrB/TolC efflux pump system (Table 2). By contrast, the oqxA component of OqxAB multidrug efflux pump and mdfA efflux pump were detected at much lower frequencies of 14.3% and 9.5%, respectively. The only biocide tolerance genes detected were qacA/B (4.8% of isolates) and qacE (9.5%). One isolate positive for qacE (UJAS39) also tested positive for the integrase gene intII described in Class I integrons; however, PCR amplification with a forward primer for qacE and a reverse primer for intII did not yield any amplification. Isolate UJAS39 also tested positive for the sulfonamide resistance gene sul1. However, sul1 was found in a very low percentage of isolates (9.5%). Similarly, dfra12 and dfra15 were detected in a very low percentage (0.05% for both). The investigated beta-lactamase genes were also found in low percentages: (bla TEM, 14.3%; bla CTXM-2, 4.8%; bla PSE, 4.8%). The only TC resistance genes detected were tet(A) (9.5%) and tet(C) (4.8%). The phenicol resistance gene floR was detected in 19.05% of isolates. The rest of antimicrobial resistance genes investigated were not detected.
Discussion
In the present study, a high percentage of isolates required biocide concentrations for growth inhibition that were above the wild-type MICs for at least one of the biocides tested, specially for the bisphenol CF and the oxidizing agent P3-oxonia. Presumably, Salmonella strains with higher tolerance to disinfectants, such as P3-oxonia or other biocides/disinfectants, would be expected to also have greater possibilities for survival to disinfection processes, especially when biocide solutions become diluted below their effective concentrations (like for example by mixing with residual liquids) or if biocide-tolerant strains are attached to surfaces in the form of biofilms. Furthermore, the positive correlations found with tolerance to different biocides could be taken as an argument for co-selection of biocide.
Another concern is antibiotic resistance of biocide-tolerant strains (Buffet-Bataillon et al., 2012; Ortega Morente et al., 2013). Ampicillin resistance was common in biocide-tolerant Salmonella isolates from the present study. Furthermore, two isolates (UJAS18 and UJAS19) were resistant to ceftazidime, cefotaxime, and ciprofloxacin, the last two being used commonly for the treatment of invasive Salmonella infections in humans. These two isolates were resistant to nine different antimicrobials and tolerant to four or five biocides. The strong positive correlations found between antibiotic resistance and the moderate or strong positive correlations between antibiotic resistance and biocide tolerance suggest that exposure to biocides could co-select for antibiotic resistance and facilitate persistence of antibiotic-resistant Salmonella in food premises. A previous study from our group on biocide resistance in Gram-negative bacteria from organically produced foods (other than eggs) showed that many bacterial isolates were simultaneously tolerant to biocides and antibiotic resistant (Fernández Fuentes et al., 2014). Also, Salmonella strains adapted by repeated exposure to TC and CF showed an increased tolerance to biocides and antibiotics (Gadea et al., 2016).
It has been suggested that antibiotic resistance in biocide-adapted strains may be mediated by efflux pumps through their upregulation and/or permeability alterations (Condell et al., 2012). Most antibiotics tested in the present study (eg., ampicillin, chloramphenicol, ciprofloxacin, nalidixic acid, and tetracycline) can be exported by efflux pumps of the resistance nodulation and cell division family (Poole, 2004). Particularly, the acrB gene of the AcrAB-TolC efflux pump system was detected in all isolates. However, other efflux pump systems investigated, such as mdfA of the major facilitator superfamily (MDF), the olaquindox efflux pump oqxA, and the biocide tolerance genes qacA/B and qacE, had low frequencies. Altogether, these results suggest that the observed biocide tolerance may not be due to acquisition of specific biocide tolerance genes, but rather to nonspecific adaptations that possibly may include changes in membrane susceptibility to biocides and/or activation of nonspecific efflux pumps.
Antibiotic resistance genes of special relevance detected in the present study includes blaTEM , blaCTXM-2 , and blaPSE . Antimicrobial resistance in Salmonella has been linked with integrons (Boyd et al., 2000; Doublet et al., 2005). Particularly, class 1 integron (Boyd et al., 2000; Levings et al., 2005) may confer pentaresistance to ampicillin (bla PSE-1), chloramphenicol/florfenicol (floR), streptomycin/spectinomycin (aadA2), sulfamethoxazole (sul1), and tetracycline [tet(G)] (Boyd et al., 2001). The integron may also contain a diverse array of other resistance genes (Beutlich et al., 2011), including QAC resistance genes qacEΔ1 and qacH (Mulvey et al., 2006; de Toro et al., 2011). Antimicrobial resistance genes associated with Salmonella class 1 integron were detected in isolates from the present study: floR, dfrA12, dfrA15, bla PSE, bla TEM, and qacE. Particularly, isolates UJAS38 and UJAS39 also carried sul1, and UJAS38 also tested positive for the integrase gene intI1. Both isolates carried tet(A) and bla TEM, a combination also found in the Salmonella genomic island variant SGI1-K1 (Beutlich et al., 2011). These isolates deserve to be further investigated to determine the possible integron association of the detected genetic determinants of resistance.
In conclusion, results from the present study indicate a positive correlation between biocide tolerance and antibiotic resistance in Salmonella isolates from eggshells. Biocide-tolerant isolates also carry antibiotic resistance genes. Rotation of the type of biocides used for disinfection processes or the use of alternative antimicrobial agents such as bacteriophages could be recommended to avoid possible selection of antibiotic resistance by biocides.
Footnotes
Acknowledgment
This work was financed by the University of Jaén (Plan de Apoyo a la investigación, proyectos en Biomedicina. Ref. UJA2013/10/06).
Disclosure statement
No competing financial interests exist