
Abstract
Introduction:
The Gram-positive bacterium Listeria monocytogenes is a ubiquitous intracellular pathogen, which has been implicated within the past decade as the causative organism in several outbreaks of foodborne disease. Bacterial ghosts (BGs) are nonliving and vacant cell envelopes of bacteria generated by releasing the bacterial cytoplasm through a channel in the cell envelope. This study attempted to produce Gram-positive pathogenic L. monocytogenes ghosts (LMGs) with simple chemicals.
Materials and Methods:
The generation of LMGs was based on minimum inhibition concentrations of 1.10 mg/mL NaOH, 0.0675 mg/mL SDS, and 0.035% (v/v) H2O2. The potential efficacy of LMGs as vaccines and their ability to induce protective immune responses against virulent L. monocytogenes challenge through multipoint subcutaneous injection were also evaluated.
Results:
The detected activity and viability of LMGs showed that nonliving LMGs could be induced. The detected LMG DNA and protein, as well as its morphological features, indicated that the produced LMGs were empty cells with the correct morphological structure. The subcutaneous vaccination of LMGs through multipoint injection conferred effective protection, with an antibody titer that reached 104 U.
Conclusions:
These findings strongly suggested that this chemical method can be used to produce LMGs and could be useful in future vaccine development against this foodborne pathogen.
Introduction
T
Bacterial ghosts (BGs) are nonliving empty cell envelopes of bacteria generated by releasing the bacterial cytoplasm through a channel in the cell envelope, which was previously created through the controlled production of the bacteriophage phi X174 lysis protein E (Witte et al., 1992; Mader et al., 1997; Eko et al., 1999). This method was recognized as the potential technology to develop efficient vaccines against foodborne pathogens (Eko et al., 1999; Lubitz et al., 1999; Ebensen et al., 2004; Wang and Lu, 2009).
The superior conservation of antigenic determinants on the surface of genetically inactivated BGs makes them attractive immunogenic inactivated vaccine candidates (Ebensen et al., 2004). Meanwhile, ghosts possess all the immunostimulatory surface properties of the original host strain, but none of the infectious threats associated with live vaccines (Eko et al., 1999; Lubitz et al., 1999). Unwanted leaky expression of gene E in the absence of appropriate induction temperature reduced biomass production of the host bacterium, consequently leading to a lower yield of BG (Lubitz et al., 1999). The application of BGs as vaccines is limited because of their low lysis efficiency and production, as well as the presence of pathogenic islands and/or antibiotic resistance genes within ghost preparations (Paukner et al., 2005; Zhang and Zhaoshan, 2006).
BGs received increasing interest in recent years for their promising medicinal and pharmaceutical applications. Most of the previously reported BGs were Gram-negative bacteria (Jechlinger et al., 1999; Panthel et al., 2003; Mayr et al., 2005; Peng et al., 2011; Chen et al., 2014). However, more Gram-positive foodborne pathogenic bacteria exist, but we cannot produce their BGs because no E lysis gene has been detected in Gram-positive bacteria.
BG preparation was recently established by methods other than the E lysis gene. The protocols were able to convert viable cells to BGs, mainly on the basis of critical concentrations of chemical compounds (Amara et al., 2013b; Amro et al., 2014; Vinod et al., 2014). BGs retain all morphological, structural, and antigenic features of the cell wall and can be used as a vaccine candidate. Besides, they can be exploited as a delivery system for proteins, which are either expressed or anchored to the envelopes before lysis or subsequently loaded (Haslberger et al., 2000). To prevent Listeria infection, finding a new method, which can be successful, used to generate LMGs is very important.
In the present study, the Gram-positive pathogenic L. monocytogenes ghosts (LMGs) were produced by treatment with simple chemicals. The potential efficacy of LMGs as vaccines and their ability to induce protective immune responses against virulent L. monocytogenes challenge in rats were also evaluated.
Materials and Methods
Bacterial strain and culture condition
L. monocytogenes CMCC54004 was provided by Jiangsu Provincial Center for Disease Control and Prevention in the People's Republic of China. L. monocytogenes was propagated in brain heart infusion (BHI) medium (Land Bridge Co. Ltd., Beijing, China) under aerobic conditions at 37°C on a shaking incubator at 200 rpm. The growth of L. monocytogenes was spectrophotometrically measured with optical density at 600 nm (UV-vis spectrophotometer, U-3900; Hitachi, Japan).
Detection of minimum inhibition concentration
Standard experiments to determine the minimum inhibition concentration (MIC) for NaOH, SDS, and H2O2 were conducted (Andrews, 2001). Briefly, agar dilution plates, which contain different concentrations of NaOH, SDS, or H2O2, were prepared. L. monocytogenes was coated on these plates under aerobic conditions at 37°C, then detection of MIC of these chemicals above. The final concentration of the CaCO3 solution used to prepare LMGs was 0.35 μg/mL as described by Amara et al. (2013b).
Production of LMGs
The production of LMGs was performed as previously described (Amara et al., 2013a, 2013b) with some modifications. The production of LMGs is described in Figure 1. In brief, the biomass of cultivated L. monocytogenes was collected by centrifugation (5000 × g for 15 min) and washed gently thrice with 0.5% sterile saline (NaCl solution). The collected L. monocytogenes cells were resuspended in 0.5% sterile saline to OD600 of 0.6∼0.8, which equals a cell concentration of about 106 CFU/mL. A 8 × stock solution was prepared for each NaOH, SDS, and H2O2, as determined from the MIC.
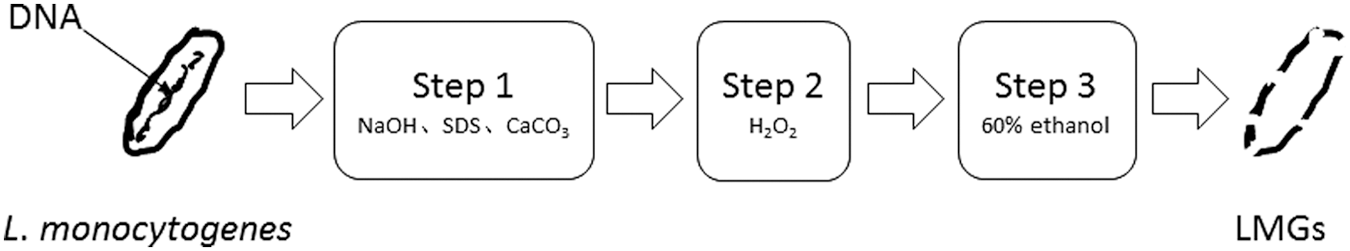
schematic illustration of LMGs generation. Live Listeria monocytogenes cells were changed into dead LMGs after treatment of three steps. LMG, L. monocytogenes ghost.
Step 1, 10 mL of 8 × stock NaOH, CaCO3, and SDS was taken and added into 50 mL of the bacterial suspension (106 CFU/mL). This mixture was incubated at 37°C for 1 h on a shaking incubator at 100 rpm. After incubation, the mixture was centrifuged at 5000 × g for 20 min.
Step 2, the cell pellets were suspended in 50 mL of 0.5% sterile saline before the preprepared H2O2 stock solution (8 × ) was added, for a final concentration as determined from the MIC. This mixture was incubated at 37°C for 30 min on a shaking incubator at 100 rpm. After incubation, the mixture was centrifuged at 5000 × g for 20 min.
Step 3, the cells were suspended in 20 mL of 60% ethanol at room temperature for 30 min with gentle vortexing of 30 s each for 5 min. The mixture was centrifuged at 5000 × g for 20 min. LMGs were harvested by centrifugation (5000 × g for 20 min) and washed thrice with 0.5% sterile saline. The final pellet was resuspended in sterile phosphate-buffered saline (PBS) (137 mmol/L NaCl; 2.7 mmol/L KCl; 10 mmol/L Na2HPO4; 2 mmol/L KH2PO4, pH 7.4) and stored at 4°C until further use.
Viability assay
To determine the bacterial viability at each step in producing LMGs, 50 μL of the suspension was spread onto BHI agar plates and incubated for 5 d at 37°C. At the same time, the bacterial viability kit (Molecular Probes BacLight; Thermo Fisher Scientific) was used to determine the live and dead cells during the production of LMGs according to the manufacturer's instructions and other reports (Gasol et al., 1999; Stocks, 2004) by mixing the suspension with the probes and visualizing under a microscope.
DNA and protein concentration and genomic DNA extraction
To analyze the release of DNA and proteins from inside the cells in each step, the DNA concentration was determined by measuring the absorption at 260 nm (UV-vis spectrophotometer, U-3900; Hitachi, Japan). The protein concentration was determined by measuring the absorption at 280 nm (UV-vis spectrophotometer, U-3900; Hitachi, Japan).
The genomic DNA of LMGs was extracted using an EZ-10 spin column bacterial genomic DNA isolation kit (Sangon Biotech, China) according to the manufacturer's protocol. The existence of genomic DNA of LMGs was examined by 1% agarose gel electrophoresis. The gel was detected by ChemiDoc XRS+ system (Bio-Rad Laboratories, Inc.) and photographed.
Morphological features
The morphological features of LMGs were examined by scanning electron microscopy (SEM). LMG cells were deposited on cover glass fixed on carrier by conductive double-faced adhesive tape and sputter coated with gold. The SEM observation was performed with a field emission scanning electron microscope (S-4800; Hitachi, Japan) at 10 kV.
The internal structural features of LMGs were examined by transmission electron microscopy (TEM). After fixation, dehydration, and sectioning, the LMG cells were observed under a transmission electron microscope at 80 kV, according to the method of Cheng et al. (2014).
Experimental animals
The 6-week-old female Balb/c rats, weighing ∼18 g, were maintained at 25°C in a 12-h light/12-h dark cycle with access to standard pellet diet and water. All animal experimental procedures were approved by the Institutional Animal Care and Use Committee at Beijing Kwinbon Biological Technology Co. Ltd. (Beijing, China).
Immunization and challenge experiment
To assess the immunogenicity and protective efficacy of LMGs, a group of 10 rats were equally divided into Groups A and B (n = 5 per group). The rats in group A were nonimmunized controls that received sterilized PBS. The rats in group B were immunized with 2 mL of LMGs (1 × 108 cells/mL) in sterile PBS by multipoint subcutaneous injection. The rats in groups A and B were immunized three times at 2-week intervals.
To determine the immune response, blood samples were obtained from the tail veins of individual rats at 2-week intervals during immunization and determined by an ELISA kit (Beijing Kwinbon Biological Technology Co. Ltd., Beijing, China) according to the protocol. Two weeks after the last immunization, all the rats were intravenously challenged with 2 mL of virulent L. monocytogenes (1 × 108 CFU/mL) in sterile PBS. The viability of the rats in groups A and B was recorded after the challenge experiment.
Statistical analyses
Statistical analyses were performed with one-way ANOVA using SPSS software (version 20.0). Data are expressed as mean ± standard error of the mean. Differences were considered statistically significant if p < 0.05.
Results
MIC of chemicals
The determination of results showed that the MIC of NaOH, SDS, and H2O2 for L. monocytogenes was 1.10, 0.0675 mg/mL, and 0.035% (v/v), respectively.
Viability
In generation of LMGs, the live L. monocytogenes cells will be changed into dead LMGs. The viability of L. monocytogenes cells/LMGs changed from 1 × 106 CFU/mL in the first processing step to 0 CFU/mL in the last processing step (p < 0.05) in Figure 2 (numerical variation of the logarithm). Nonliving LMGs were detected in the third-step process. In the present study, the aforementioned method showed 100% lysis efficiency.

Viability of LMG cells at different steps. The cells were spread onto BHI agar plates and incubated for 5 d at 37°C. The X-axis indicates the steps of LMG production. Data are presented as the mean ± SE of three independent experiments. Different letters in the same column indicate a statistical difference between the mean values (p < 0.05). BHI, brain heart infusion; LMG, L. monocytogenes ghost; SE, standard error.
Figure 2 shows the determination of living or dead L. monocytogenes cells during LMG production. After step-by-step treatment, the number of live L. monocytogenes cells was gradually reduced; ultimately, no live L. monocytogenes cells remained. These results were also confirmed by using the bacterial viability kit, as there were all red cells left after the third-step treatment (data not shown). These data suggested that the MIC of NaOH, SDS, and H2O2 would be sufficient to produce inactivated bacteria from L. monocytogenes. Nonliving LMGs suggested its safe use in the immune experiment.
DNA and protein concentration and genomic DNA extraction
As shown in Figure 3a, the total DNA content was well comparable at the beginning with the untreated control and chemical-treated L. monocytogenes cells. During the LMG production, the DNA content increased and then declined (p < 0.05); the protein content followed a similar trend during the processing (Fig. 3b). The change in the DNA and protein concentration indicated the occurrence of cell lysis, which was monitored during the preparation of BGs.

Release of LMG DNA
The presence of genomic DNA from LMGs was examined by agarose gel electrophoresis, detected by the ChemiDoc XRS+ system, and photographed. Figure 4 shows the existence of genomic DNA of L. monocytogenes and LMGs. The genomic DNA of L. monocytogenes was much more than LMGs. The genomic DNA of L. monocytogenes was probably degraded during LMG production.
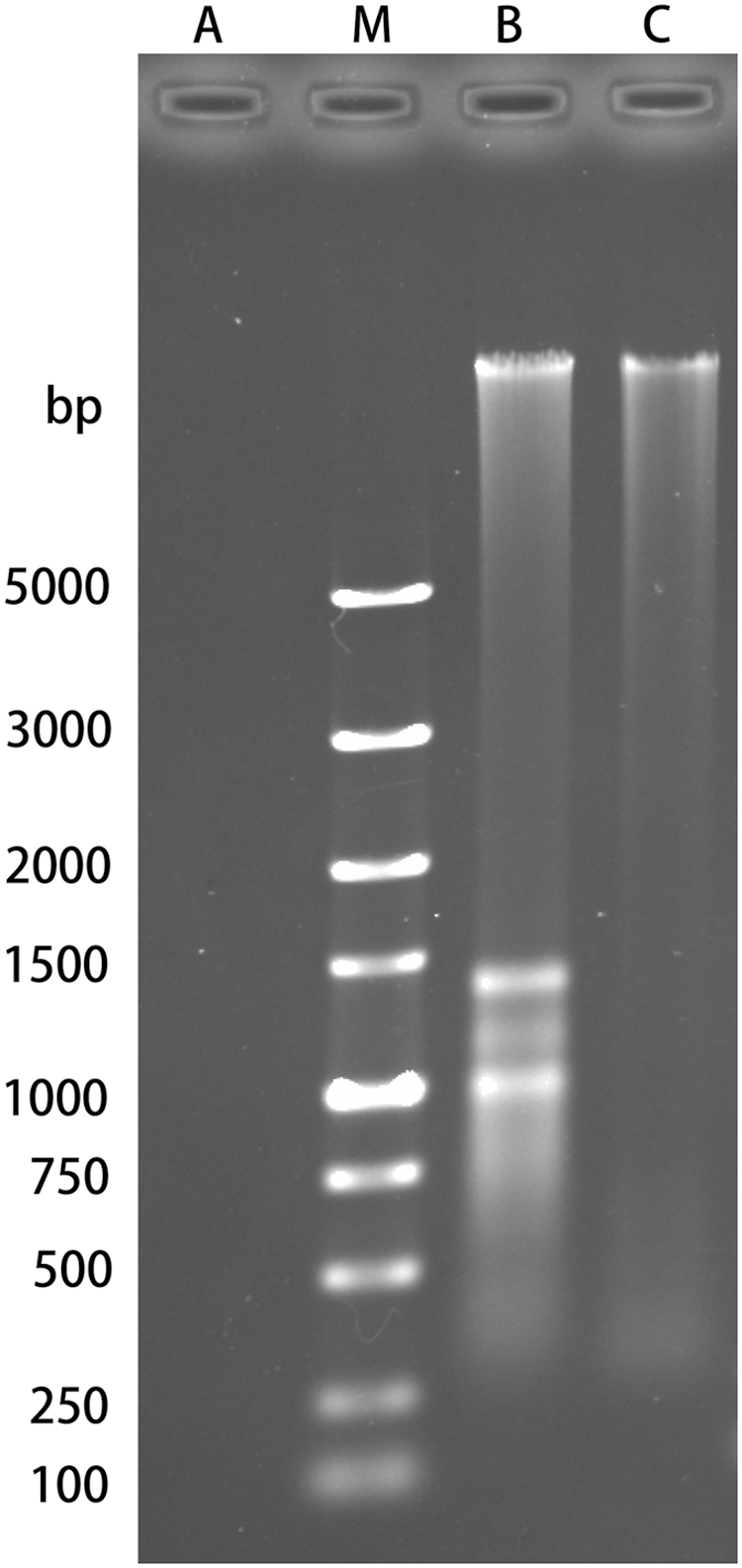
Presence of genomic DNA in LMGs by agarose gel electrophoresis. Lane A, negative control (without DNA); Lane M, DNA standard; Lane B, genomic DNA of LMGs; Lane C, genomic DNA of L. monocytogenes. LMG, L. monocytogenes ghost
Morphological features
As shown in Figure 5, the LMGs had a morphological structure similar to that of L. monocytogenes. Therefore, LMGs may have all the immunostimulatory surface properties of the original host strain, L. monocytogenes. The internal structural features of LMs were examined by TEM. As shown in Figure 6, the LMGs are empty cell envelopes produced by releasing the bacterial cytoplasm through a channel in the cell envelope. The morphological features of LMGs indicate the successful production of LMGs.
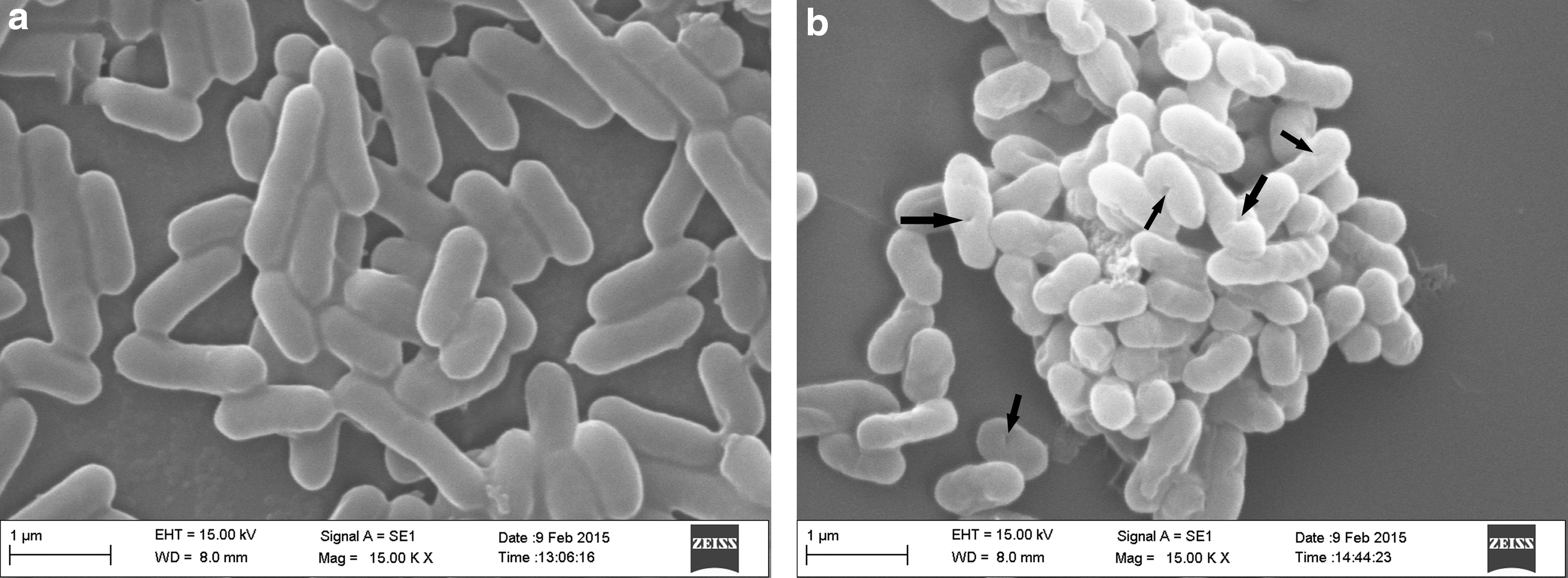
Morphological features of
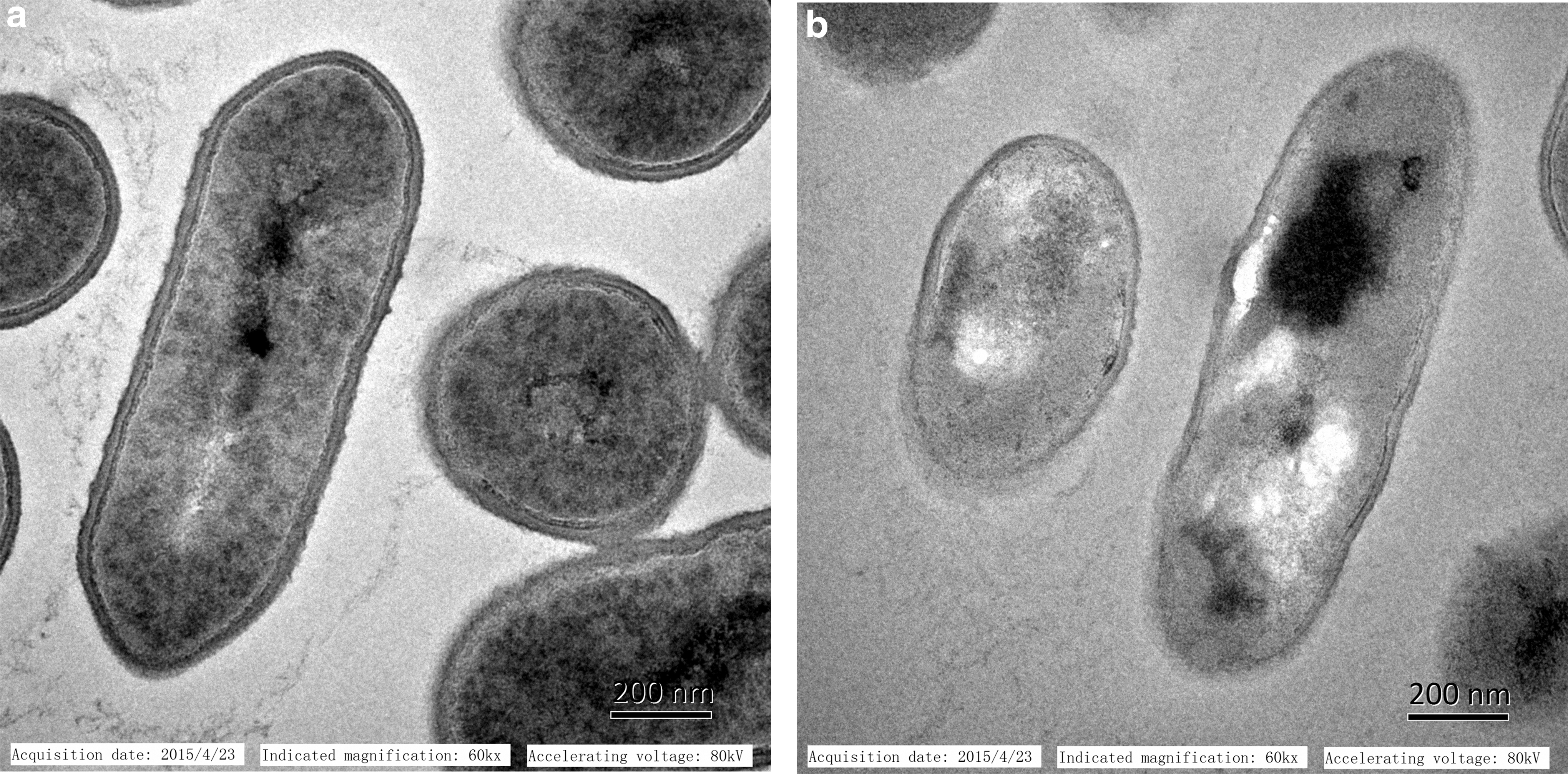
Morphological features of
Immunization and challenge experiment
In this study, the rats in group B produced a significant immune response (p < 0.05), with an antibody titer of serum up to 104 U (Fig. 7). The viability of rats from groups A and B is shown in Figure 8. All the rats in group A failed to pass the challenge experiment (0% survival), whereas 100% survival was achieved in group B. LMGs could be used to induce protective immune responses against virulent L. monocytogenes challenge in rats.
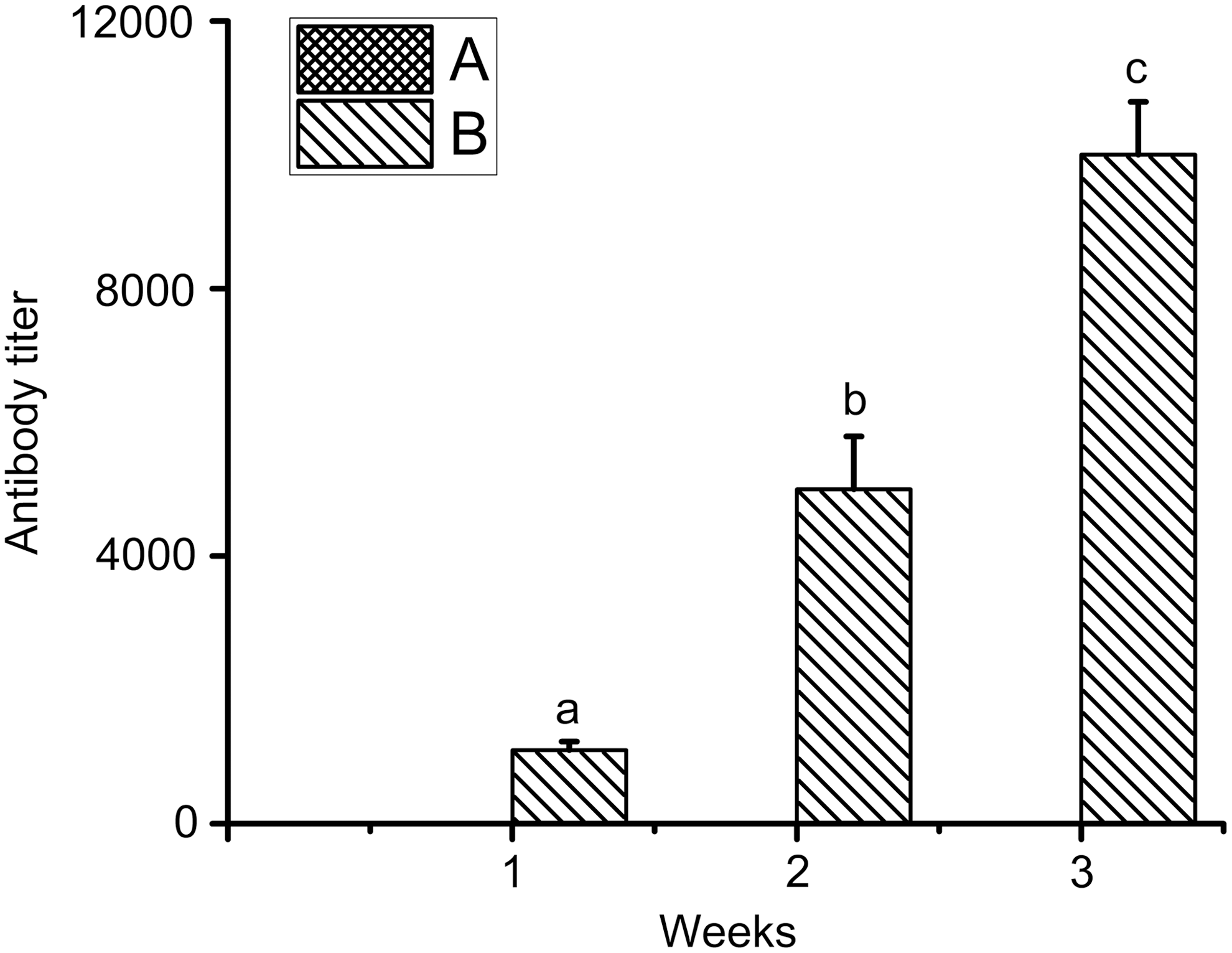
Antibody titer of rats immunized by LMGs. Antibody titer was not detected in group A (black) rats immunized by PBS. Data are presented as mean ± SE of three independent experiments. Different letters in the same column indicate statistical difference between the mean values (p < 0.05). LMG, L. monocytogenes ghost; PBS, phosphate-buffered saline.
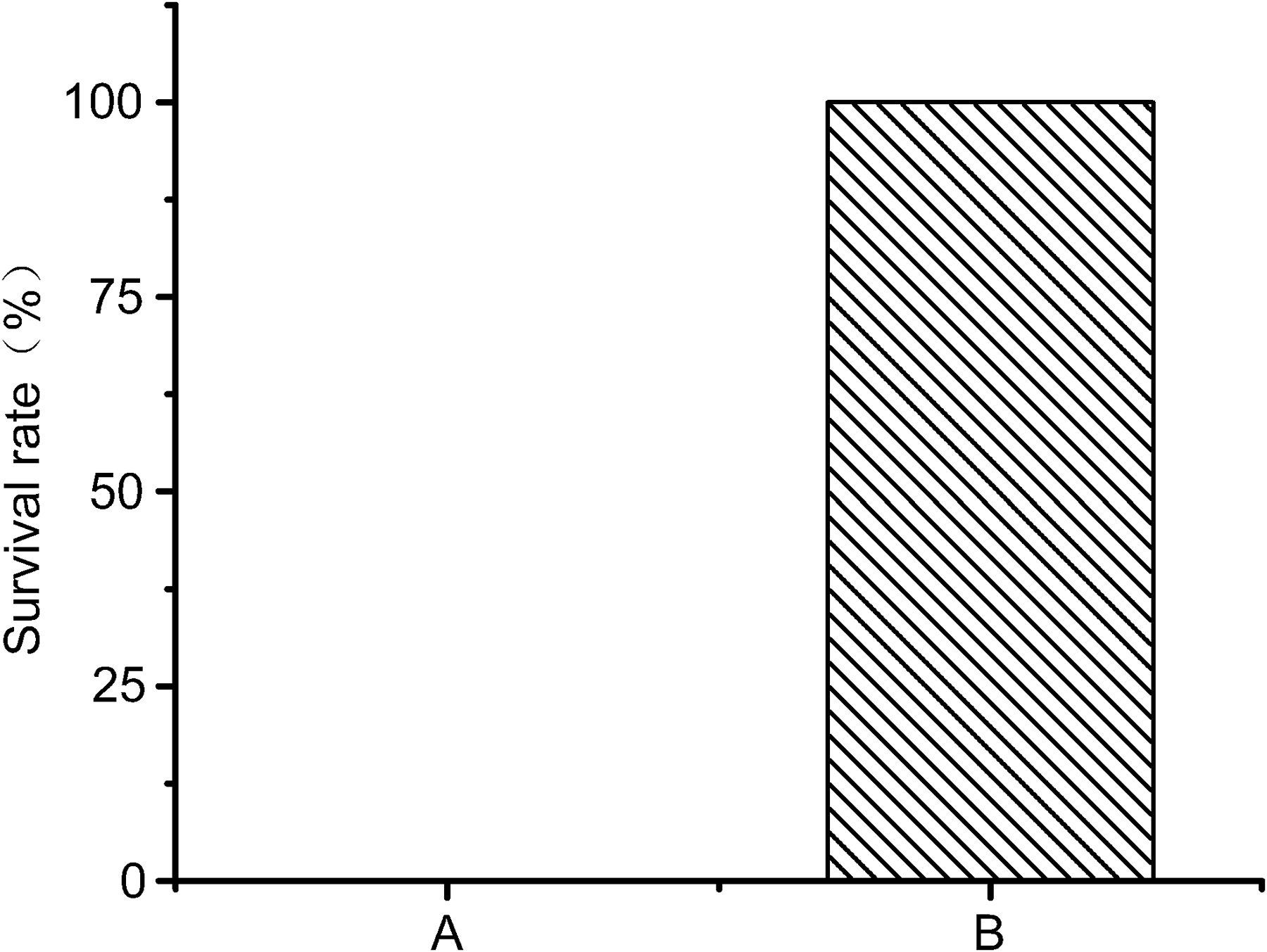
Survival of rats in groups A and B of the challenge experiment. The survival rate of rats in group A is 0%.
Discussion
In the past, BG was previously created through the controlled production of the bacteriophage phi X174 lysis protein E (Witte et al., 1992; Mader et al., 1997; Eko et al., 1999), which exits in Gram-negative bacteria. In the present study, the Gram-positive pathogenic LMGs were produced by a novel method without phi X174 lysis protein E. It may bring new opportunities to produce Gram-positive BGs.
The application of BGs as vaccines was limited because of their low lysis efficiency and production within ghost preparations (Paukner et al., 2005; Zhang and Zhaoshan, 2006). The new method in this study has much higher lysis efficiency and production. The results in this study strongly suggest that this chemical method could be used to generate Gram-positive BGs. Recently, Staphylococcus aureus Ghost, a Gram-positive BG, was successful generated by NaOH (Vinod et al., 2015). It was similar with the method used in our study. The method in this study could be a common method for generation of Gram-negative and Gram-positive BGs. A major problem for generation of BGs seems to be the choice of appropriate concentrations of these chemicals above. The easy manufacturing process and low production costs provide additional advantages for mass implementation of this method.
BGs possess all the immunostimulatory surface properties of the original host strain, but none of the infectious threats associated with live vaccines (Eko et al., 1999; Lubitz et al., 1999). In this study, LMGs were produced with the correct morphological features and intact antigens on the cell membrane, which are necessary of protective immune responses against virulent strains. The potential efficacy of LMGs as vaccines and their ability to induce protective immune responses against virulent L. monocytogenes challenge in rats were evaluated. LMGs could be used to induce a protective immune response against virulent L. monocytogenes challenge in rats.
It was reported that the existence of BG genomic DNA is a potential risk in the immune system (Ebensen et al., 2004; Paukner et al., 2005). In this study, we find there was a little genomic DNA in LMGs, and this LMGs generated can be used in rats immunization against virulent L. monocytogenes. In the present study, the Gram-positive pathogenic LMGs were produced by treatment with simple chemicals containing NaOH, CaCO3, SDS, and H2O2. To precisely control the lysis of BGs, the time of each step process and no genomic DNA in BGs need further study in the follow-up research.
The BGs of Escherichia coli (Witte et al., 1992), Vibrio cholera (Eko et al., 2000), Pasteurella multocida (Marchart et al., 2003a), Pasteurella haemolytica (Marchart et al., 2003b), and Helicobacter pylori (Mayr et al., 2005) have been successfully produced and can be used to induce protective immune responses against corresponding virulent bacteria. In the present study, the Gram-positive foodborne pathogen L. monocytogenes was added to the BG family.
Conclusion
The LMGs were produced with the correct morphological features and retained intact antigens on the cell membrane with simple chemical treatment. The LMGs produced by this method can be used to induce a protective immune response against virulent L. monocytogenes challenge in rats.
Footnotes
Acknowledgments
This project was financially supported by the National Natural Science Foundation of China (no. 31271930) and the Innovation of Graduate Culturing of Jiangsu Province (KYLX_1167).
Disclosure Statement
No competing financial interests exist.