
Abstract
Endophytic bacteria (EB) are a rich source of secondary metabolites with medicinal importance. In this study, EB were isolated from the bottle brush herb Equisetum arvense and identified based on 16S rRNA sequencing. Evaluation of its antibacterial potential was conducted using two common foodborne pathogenic bacteria, Staphylococcus aureus ATCC 12600 and Escherichia coli O157:H7 ATCC 43890. Out of 103 identified EB, three species, Streptomyces albolongus, Dermacoccus sp., and Mycobacterium sp., showed significant antibacterial activity against S. aureus with inhibition zones of 45.34 ± 0.15, 43.28 ± 0.19, and 22.98 ± 0.18 mm, respectively, whereas only two species, Streptomyces griseoaurantiacus (EAL196) and Paenibacillus sp. (EAS116), showed moderate antibacterial activity against E. coli O157:H7 with inhibition zones of 9.41 ± 0.29 and 10.44 ± 0.31 mm, respectively. Furthermore, ethyl acetate extract of S. albolongus, Mycobacterium sp., and Dermacoccus sp. showed antibacterial activity against S. aureus, with inhibition zones of 23.43 ± 0.21, 21.18 ± 0.22, and 19.72 ± 0.10 mm, respectively. The methanol extract of Dermacoccus sp. and Paenibacillus sp. showed antibacterial activity against S. aureus and E. coli O157:H7, with inhibition zones of 11.30 ± 0.17 and 10.01 ± 0.21 mm, respectively. Scanning electron microscopy indicated swollen and lysed cell membranes of pathogens treated with ethyl acetate extract. A possible reason might be, likely due to EB metabolites penetrating the bacterial cell membranes and affecting various metabolic functions resulting in lysis. To the best of our knowledge, this is the first study to report that EB from E. arvense can be used as a source of natural antibacterial compounds against foodborne pathogenic bacteria.
Introduction
E
Foodborne pathogenic bacteria are considered primary cause of foodborne diseases and there is demand from food industries for management of these (Menpara and Chanda, 2013; Mith et al., 2014; Afshordi et al., 2016). Escherichia coli and Staphylococcus aureus are responsible for a variety of health hazards (Rani, 2011; Addis and Sisay, 2015). Awareness and demand for fresh and negligibly treated food products without extra chemical additives by the customers have stimulated many studies of novel food preservation approaches. Naturally occurring compounds can be helpful for increasing the shelf life of various food products. To address the management of foodborne-related diseases, there is an immediate need for model antibacterial agents (Cetojevic-Simin et al., 2010; Sinha, 2012; Khaled et al., 2016).
E. arvense (horsetail plant) is a well-known perennial bottle brush herb. It generally grows in moist loamy or sandy soil (Geetha et al., 2011; Turker and Turkay, 2016). In many countries, this plant is used in medicines containing a highly complex pattern of phenolic compounds with a high concentration of silica, which makes a distinct contribution to its medicinal properties (Pereira et al., 2012; Altameme et al., 2015). However, no studies have isolated endophytes from E. arvense and evaluated their antibacterial capacity. Therefore, this study was undertaken to evaluate the antibacterial properties of EB isolated from E. arvense against foodborne pathogens.
Materials and Methods
Isolation and characterization of EB
E. arvense L. was obtained from Yeungnam University campus, Korea, during May 2014. Isolation of EB was conducted by a standard procedure (Costa et al., 2012). Two grams of each tissue was sterilized using 70% ethanol for 60 s, followed by 2% sodium hypochlorite for 90 s and 100% ethanol for 30 s. Finally, tissues were rinsed several times in sterilized distilled H2O and dried properly on sterilized filter papers. After completely drying, tissues were ground with 6 mL aqueous NaCl solution (0.9%) using a sterilized mortar and pestle and then held at room temperature for 3 h. The tissue extracts were diluted 10 or 100 times and plated on yeast extract, nutrient broth, and agar (YNA) media and incubated for 15 days at 28°C. Morphological characterization was conducted after the appearance of visible colonies, and after 15 days, all colonies were counted and expressed in log colony-forming unit (CFU)/g tissue.
The isolated EB were identified based on 16S rRNA sequencing using an ABI Prism 3730xl DNA sequencer (Geno Tech). The sequencing results, alignment, and the phylogenetic tree analysis were conducted using Molecular Evolutionary Genetics Analysis (MEGA) 6 software (version 6). Determination of phylogenetic trees was conducted by the neighbor-joining method (Kamei and Isnansetyo, 2003; Tamura et al., 2013; Susilowati et al., 2015).
Antibacterial assay against foodborne pathogenic bacteria
Staphylococcus aureus ATCC 12600 and Escherichia coli O157:H7 ATCC 43890 used in the study were obtained from American Type Culture Collection (ATCC) and maintained on YNA media.
Antibacterial screening was conducted following the standard procedure (Roh et al., 2009). EB were grown for 24 h at 28°C in yeast extract and nutrient broth (YNB; Becton, Dickinson and Company). Next, 10 μL of overnight grown culture (OD600 = 1.0) of EB was dropped gently onto YNA plates (1.5% agar), allowed to dry for 10 min, and then incubated at 28°C for 24 h. The patches of EB were then killed by adding 1 mL chloroform to the lids of the Petri plates and then the plates were inverted and kept for 10 min. The lids of the plates were opened, after which the open Petri plates were allowed to stand for 30 min to remove traces of chloroform. The Petri plates were then treated with UV light for 15 min to kill the bacteria completely. Next, 35 μL of freshly grown cultures of E. coli O157H7 and S aureus were suspended in 10 mL of YNA (0.75% agar) at 55°C, mixed gently, and then poured over the killed bacteria patches. After solidification, Petri plates were incubated at 28°C for 24 h, and the diameter of the zone of inhibition was measured. Rifampicin and streptomycin at 10 μg/disc were taken as reference standard.
Antibacterial activity of solvent extracts
The solvent extraction of secondary metabolites from EB with positive antibacterial activity was accomplished by standard procedure (Zubair et al., 2011). Briefly, EB were grown in 200 mL YNB at 28°C for 4 days. After incubation, the cultures were centrifuged at 1000 × g for 20 min at 4°C. The supernatant without bacteria (filter sterilized using membrane filter) was then added to same volume of ethyl acetate and kept overnight in a shaking incubator at 150 rpm and 28°C. Next, 50 mL of methanol was added to the bacteria pellet and sonicated for 10 min at room temperature, with overnight shaking. The bacteria pellet with methanol was then centrifuged at 1000 × g for 20 min at 4°C. The supernatant of the methanol fraction was subsequently collected and the ethyl acetate fraction was separated through a separating funnel. Both ethyl acetate and methanol fractions were dried in a rotary evaporator at 50°C. Based on the weight of the metabolite, 5% dimethyl sulfoxide (DMSO) was added to the ethyl acetate fraction and sterilized distilled H2O was added to the methanol fraction (crude extract). A small amount of methanol extract was kept to check for antibacterial activity, while the rest was mixed with an equal volume of hexane and then incubated overnight. The hexane fraction was then separated using a separating funnel and dried using a rotary evaporator. DMSO (5%) was added to it to give a final value of 0.1 g/mL.
Antibacterial assay of the solvent extracts was conducted by disc diffusion method (Diao et al., 2013). Briefly, 35 μL of freshly grown test pathogens (OD600 = 1.0) were mixed in 10 mL YNA soft agar (0.75%) at 55°C. Following solidification of the Petri plates, but before analysis, sterilized 8 mm paper discs (Advantec; Toyo Roshi Kaisha, Ltd.) were prepared by adding 50 μL of solvent extracts (500 μg/disc) onto the Petri plates, after which the samples were incubated at 37°C for 24 h and the zone of inhibitions were measured using an electronic digital caliper (M500-182M, Konex; Tool Parts Company) with precession upto 1/100 mm.
The minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of three solvent fractions (ethyl acetate, methanol, and hexane) were evaluated against S. aureus and E. coli O157:H7 by two-fold dilution method (Kubo et al., 2004). The lowest concentration of the solvent extracts that did not show any visible growth of the test pathogen was taken as the MIC value, and the lowest concentration of solvent extract that did not show any growth of pathogen on the YNA Petri plate was selected as the MBC.
Scanning electron microscopy
The effects of the ethyl acetate fractions on the surface morphology of S. aureus and E. coli O157:H7 were determined by scanning electron microscopy (SEM). For SEM analysis, pathogens were cultured overnight at 37°C for both control (treated with 5% DMSO) and treatment (treated with MIC of ethyl acetate fraction) groups (Bajpai et al., 2009). Before SEM analysis, the specimens were sputter coated with platinum for 2 min using an ion coater and then observed using an SEM (Hitachi; S-4100).
Statistical analyses
The results of all experiments were expressed as the mean ± standard deviation. Statistical analyses consisted of one-way analysis of variance and Duncan's multiple range tests, at p < 0.05 using Statistical Analysis Software (SAS) version 9.4 (SAS, Inc.). MEGA software version 6.06 was used for dendrogram cluster analysis.
Results
Isolation and identification of EB
Based on morphological characteristics, a total of 103 types of EB were isolated from the leaves and stems/roots of E. arvense. A total of 48 and 55 EB were isolated from leaves and from stems and roots, respectively. The density of EB was greater in stems and roots (4.8 ± 0.56 × 103 log CFU/g) together than in the leaves (1.3 ± 0.42 × 103 log CFU/g). The morphological characteristics of isolated bacteria are described in detail in Supplementary Table S1 (Supplementary Data are available online at
Out of all the isolated EB, 27 EB with different morphologies and positive antibacterial properties were identified by 16S rRNA sequencing analysis (Table 1). A phylogenetic tree was constructed using the 16S rRNA sequence for the five selected promising EB with their closest relative sequences. The constructed phylogenetic tree showed that EAL196 and EAS116 had the highest similarity with Streptomyces griseoaurantiacus and Paenibacillus sp., respectively, and EAL182, EAS167, and EAL169 showed maximum similarity with Streptomyces albolongus, Dermacoccus sp., and Mycobacterium sp., respectively (Fig. 1).
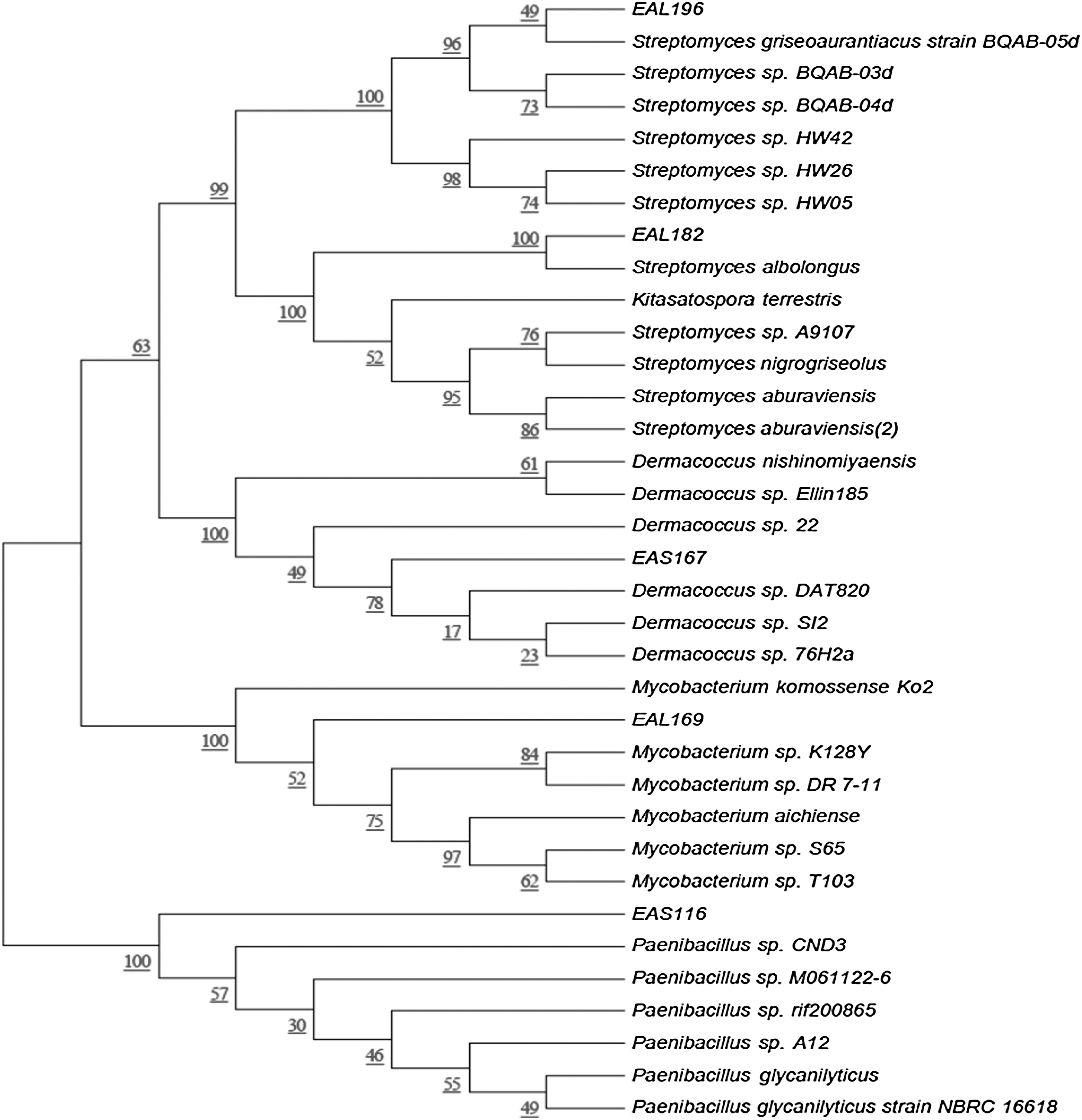
Phylogenetic tree exhibiting similarity among 16S rRNA gene sequences of five selected endophytic bacteria isolated from Equisetum arvense with the nearest identified species. Evolutionary distances were computed using the maximum composite likelihood method. Bootstrap values (1000 tree interactions) are indicated at the nodes.
Max. score, the highest alignment score (bit-score) between the query sequence and the database sequence segment; E value, the expect value (E) describes the number of hits expected when searching the database. The lower the E-value, or the closer it is to zero, the more “significant” the match is; Max Identity, highest percent of similarity between the query and subject sequences over the length of the coverage area; + activity, greater than 9 mm diameter zone of inhibition; − activity, no activity in antibacterial activity test.
Screening for the antibacterial activity
Out of the EB screened, five with significant antibacterial activity were selected for further analysis (Table 2 and Fig. 2a). Two EB, S. griseoaurantiacus (EAL196) and Paenibacillus sp. (EAS116), had positive antibacterial activity against E. coli O157:H7, while three EB, S. albolongus (EAL182), Dermacoccus sp. (EAS167), and Mycobacterium sp. (EAL169), showed positive activity against S. aureus ATCC 12600. Paenibacillus sp. and S. griseoaurantiacus only exhibited antibacterial activity against E. coli O157:H7 (Table 2 and Fig. 2a).

Diameters of zones of inhibition are expressed as the mean ± SD in mm. Different superscript letters represent significant differences at p < 0.05.
SD, standard deviation.
Antibacterial activity of solvent fractions of EB
The five selected EB showing positive antibacterial activity were further extracted with three different solvents. The cell-free ethyl acetate extract of S. albolongus (EAL182), Mycobacterium sp. (EAL169), and Dermacoccus sp. (EAS167) showed high antibacterial activity against S. aureus ATCC 12600, with zones of inhibition of 23.43, 21.18, and 19.72 mm, respectively. The EB S. griseoaurantiacus (EAL196) and Paenibacillus sp. (EAS116) had zones of inhibition of 12.44 and 11.13 mm, respectively, against E. coli O157:H7. Methanol extract of S. albolongus, Mycobacterium sp., and S. griseoaurantiacus did not show any activity against the two tested pathogens. Methanol extract of Dermacoccus sp. and Paenibacillus sp. showed inhibition zones of 11.30 and 10.01 mm against S. aureus and E. coli O157:H7, respectively. Hexane extract of S. albolongus showed positive activity against S. aureus, with an inhibition zone of 11.09 mm. The hexane extract of the other four EB did not show any inhibitory effects against the two tested pathogens (Table 3 and Fig. 2b).
Concentration of all extracts 500 μg/disc
Diameters of zones of inhibition are expressed as the mean ± SD in mm; Different superscript letters represent significant differences at p < 0.05.
NA, no activity.
The MIC and MBC values of the ethyl acetate extract ranged from 250 to 500 μg/mL and 500 to 1000 μg/mL, respectively (Table 4). MIC and MBC values of the methanol extract of Dermacoccus sp. were 500 and 1000 μg/mL, respectively, against S. aureus ATCC12600. In case of Paenibacillus sp., MIC and MBC values of the methanol extract were also 500 and 1000 μg/mL, respectively, against E. coli O157:H7. The MIC and MBC values of the hexane extract of S. albolongus were 500 and 1000 μg/mL against S. aureus, respectively.
MIC, minimum inhibitory concentration; MBC, minimum bactericidal concentration.
The results of SEM analysis of control bacteria treated with 5% DMSO and the treatment bacteria treated with MIC of ethyl acetate fractions are shown in Figure 3. The control S. aureus (Fig. 3A) bacterial cells were clear, uniform, round in shape, and present in greater concentrations than the treatment treated with ethyl acetate fraction of S. albolongus (Fig. 3B), which showed a disrupted cell membrane. In addition, treatment with Mycobacterium sp. (Fig. 3C) ethyl acetate fraction resulted in swelling, cell burst, and irregular shape of the control S. aureus bacterial cells. The Dermacoccus sp. (Fig. 3D) ethyl acetate fraction treatment resulted in cell disruption and reduced levels of control S. aureus. However, E. coli O157:H7 control (Fig. 3E) were found to be rod shaped and present in higher density, whereas the treatment bacteria with ethyl acetate fractions of S. griseoaurantiacus (Fig. 3F) and Paenibacillus sp. (Fig. 3G) had reduced bacterial cell density, elongated shape, and cell bursts (Fig. 3).
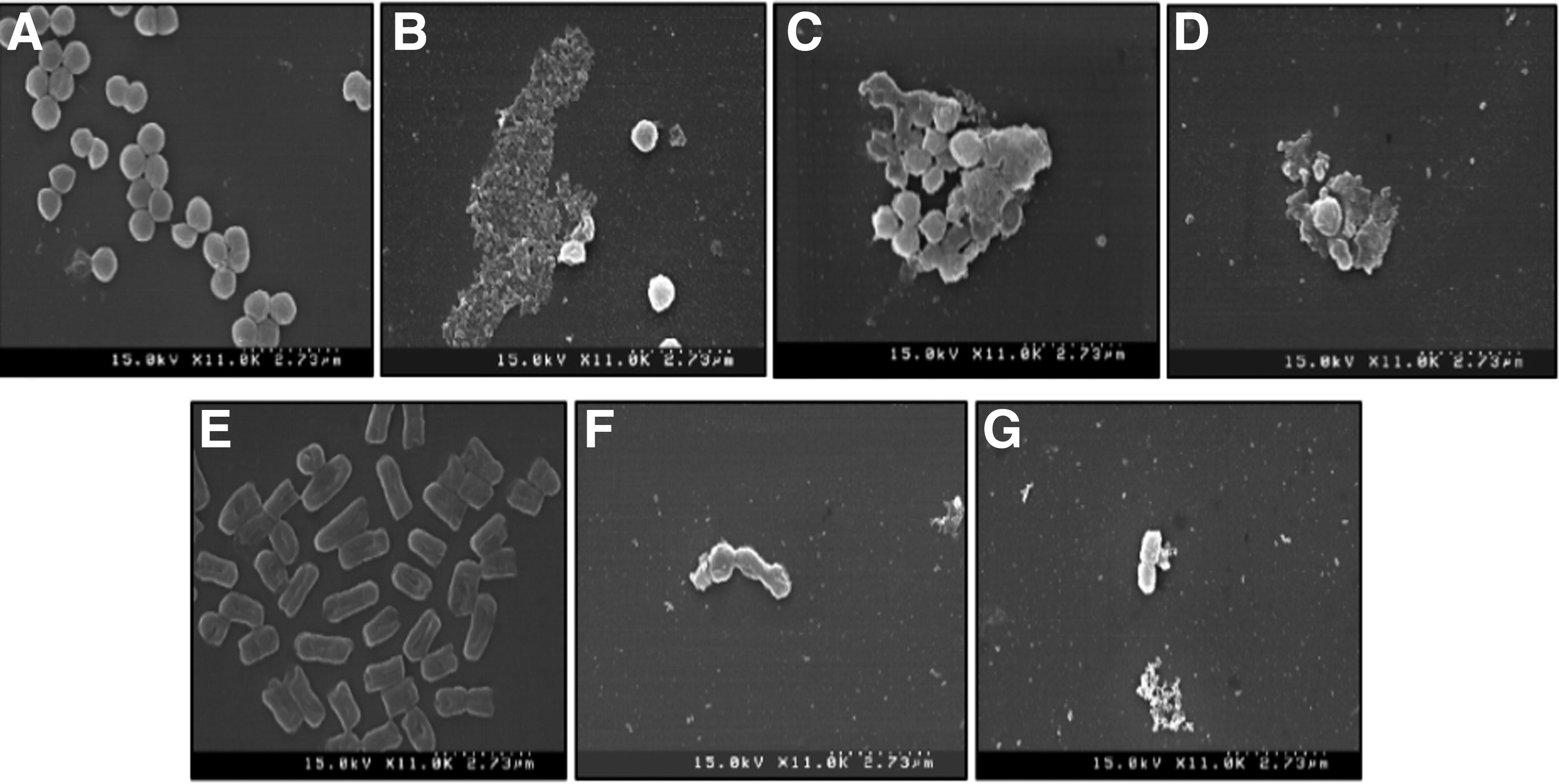
Scanning electron microscopy of solvent extracts of endophytic bacteria isolated from E. arvense.
Discussion
Food is a source for growth of various types of pathogenic microorganisms that give rise to various foodborne infections (Scott, 2003; Torgerson et al., 2015; Afshordi et al., 2016). S. aureus is a source of various diseases like food poisoning, bacteremia, sepsis, osteomyelitis, endocarditis, pneumonia, meningitis, and toxic shock syndrome (Afshordi et al., 2016). Pathogenic E. coli strains produce Shiga toxins, which can cause life-threatening sequelae hemolytic uremic syndrome, hemorrhagic colitis, and diarrhea in humans (Lim et al., 2010; Seo et al., 2016).
Microbial endophytes are promising source for the production of effective and unique metabolites with prospective therapeutic potential against pathogenic infections in the host plant (Bajpai et al., 2009; Aly et al., 2010; Kaaria et al., 2012; Christina et al., 2013; Bibi, 2015; Golinska et al., 2015; Pandey et al., 2015; Shukla et al., 2015). Because EB colonize same environmental areas that phytopathogens grow, they can produce molecules with antimicrobial activity that could act against these pathogens (Selim et al., 2014).
In various countries E. arvense is well known for its use as medicines (Dos Santos et al., 2005; Altameme et al., 2015). Therefore, in this study, EB were isolated from E. arvense, characterized morphologically and by molecular techniques, and evaluated for antibacterial activities (Tables 1 and 2 and Fig. 2). There is report of EB isolated from a medicinal plant with antibacterial activity against S. aureus and E. coli (Bibi, 2015; Shukla et al., 2015). Similar results on antibacterial properties of isolated EB from E. arvens were obtained in this study (Tables 1 and 2 and Fig. 2).
Actinomycetes are important bacteria and well exploited for producing secondary metabolites (Chaudhary et al., 2013). A number of species of Streptomyces are the most significant industrial microorganisms among the actinomycetes, because of their capacity to produce numerous bioactive molecules, mostly antibiotics (Procopio et al., 2012; Prashanthi et al., 2015; Shivlata and Satyanarayana, 2015). The genus Streptomyces produces many common antimicrobial agents with medicinal significance (Omura, 2001; Mohamed et al., 2013; Uddin et al., 2013; Shivlata and Satyanarayana, 2015). Moreover, a previous study showed that Streptomyces albolongus isolated from chilli field soil is a producer of bioactive metabolites with good antimicrobial activity against various species of human pathogens (Uddin et al., 2013). Positive antibacterial potential of the isolated Streptomyces sp. was also recorded in this investigation (Tables 2 and 3 and Fig. 2). The Dermacoccus sp. was shown to have antimicrobial and antitumor activity (Goodfellow and Fiedler, 2010; Graca et al., 2015). In our study, EAL182 and EAS167 identified as S. albolongus and Dermacoccus sp., respectively, showed significant antibacterial activity (Tables 2 and 3 and Fig. 2).
S. griseoaurantiacus isolated from the marine sediment is a producer of manumycin and chinikomycin antibiotics (Li et al., 2005, 2011). There is also report of S. griseoaurantiacus isolated from soil having anticancer property (Prashanthi et al., 2015). In this study, EAL196 identified as S. griseoaurantiacus also exerted antibacterial activity against E. coli. Mycobacterium sp. are also actinobacteria and are divided into different groups with bioactive potential (Dandie et al., 2004; Wang et al., 2006). The EAL169 identified as Mycobacterium sp. displayed a positive antibacterial activity against S. aureus (Tables 2 and 3 and Fig. 2).
Some Paenibacillus sp. are able to produce a number of secondary metabolites which acts as antibiotic agents (Lorentz et al., 2006), and the antibiotic activity of Paenibacillus has been confirmed against a wide spectrum of microorganisms (Beatty and Jensen, 2002; Berge et al., 2002; Weid von der et al., 2003; Lorentz et al., 2006; Ivy et al., 2012). There is also report of Paenibacillus sp. having a strong antimicrobial activity against E. coli and Candia albicans (Aw et al., 2016). In this study, Paenibacillus sp. (EAS116) also showed an antibacterial activity against E. coli., which confirmed its previous claim (Tables 2 and 3 and Fig. 2). The possible reason behind the specific antibacterial activity of EAS 196 and EAS 116 toward only the E. coli O157:H7 and not against the S. aureus is that, the bioactive compounds from EAS 196 and EAS 116 might be smaller in size and could have easily entered the bacterial cell through the general bacterial porins, after which it might have affected the cellular organelles like the DNA, protein, and bacterial enzymes, which might be responsible for various metabolic functions of the cell, or they might have made numerous pores in the outer cell wall of bacteria that resulted in the leakage of internal substance to the outside, resulting in the cell lysis and death (Kotzekidou et al., 2008). This is also evident from the SEM images of the control and treated bacteria that showed disruption of treatment bacteria in comparison to the smooth regular shape of the cell wall of control bacteria (Fig. 3).
Conclusions
EB have attracted a great deal of attention worldwide due to the presence of bioactive compounds with pharmaceutical significance. Because of the importance of E. arvense, many studies of its leaf extracts and isolation of endophytic fungi have been done. However, there is no report on the isolation of EB or its potential applications from this species, and this is the first report. Based on the results, EB isolated from E. arvense can be a good source of antibacterial compounds against S. aureus and E. coli O157:H7. The bacteria themselves or the extracts of the pure compounds may be useful as natural antimicrobials for food preservation. With the emergent attentiveness of people toward natural food and natural remedies, E. arvense has the potential for widespread use, especially in the developing countries in which herbs are produced and used as food additives.
Footnotes
Acknowledgment
This work was supported by a grant from the National Institute of Forest Science (Project No. FE0702-2016-02-2016 “Development of environment-friendly control agents against pine wilt disease based on BT”), National Forest Service, Republic of Korea.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.