
Abstract
Salmonella enterica serotype Napoli (S. Napoli) is currently emerging in Europe and particularly in Italy, where in 2014 it caused a large outbreak associated with elevated rates of bacteremia. However, no study has yet investigated its invasive ability and phylogenetic classification. Here, we show that between 2010 and 2014, S. Napoli was the first cause of invasive salmonellosis affecting 40 cases out of 687 (invasive index: 5.8%), which is significantly higher than the invasive index of all the other nontyphoidal serotypes (2.0%, p < 0.05). Genomic and phylogenetic analyses of an invasive isolate revealed that S. Napoli belongs to Typhi subclade in clade A, Paratyphi A being the most related serotype and carrying almost identical pattern of typhoid-associated genes. This work presents evidence of invasive capacity of S. Napoli and argues for reconsideration of its nontyphoidal category.
Article
S
In this study, we have analyzed the invasive salmonellosis cases of S. Napoli during 2010–2014 in Lombardy (10,000,000 inhabitants) and investigated its phylogenetic classification, for the first time.
Data of human salmonellosis cases in Lombardy during 2010–2014 were obtained from IT-ENTER-NET (
To infer phylogeny of S. Napoli, 93 core-genome genes (den Bakker et al., 2011) from the draft genome of the outbreak isolate SN310 (Huedo et al., 2015) were selected and concatenated, generating a DNA sequence of 73,761 nucleotides. The same orthologous sequence of 15 different serotypes of Salmonella enterica subsp. enterica—a representative sample of clade A and clade B serotypes—was extracted from a previous work (den Bakker et al., 2011). The orthologous sequence of S. enterica subsp. arizonae was used as the out-group. All DNA sequences are available in the Supplementary Data and Genbank accession numbers are listed in Figure 1. Phylogenetic analysis was done using the Maximum Likelihood method based on the Tamura–Nei model using 200 bootstrap replications in MEGA 6.0. Presence of the typhoid-associated genes hlyE and taiA (SPI-18), cdtB, pltA, and pltB (CdtB-Islet), and the GICT18/1 islet (den Bakker et al., 2011) in the genome of S. Napoli was determined using BLAST.
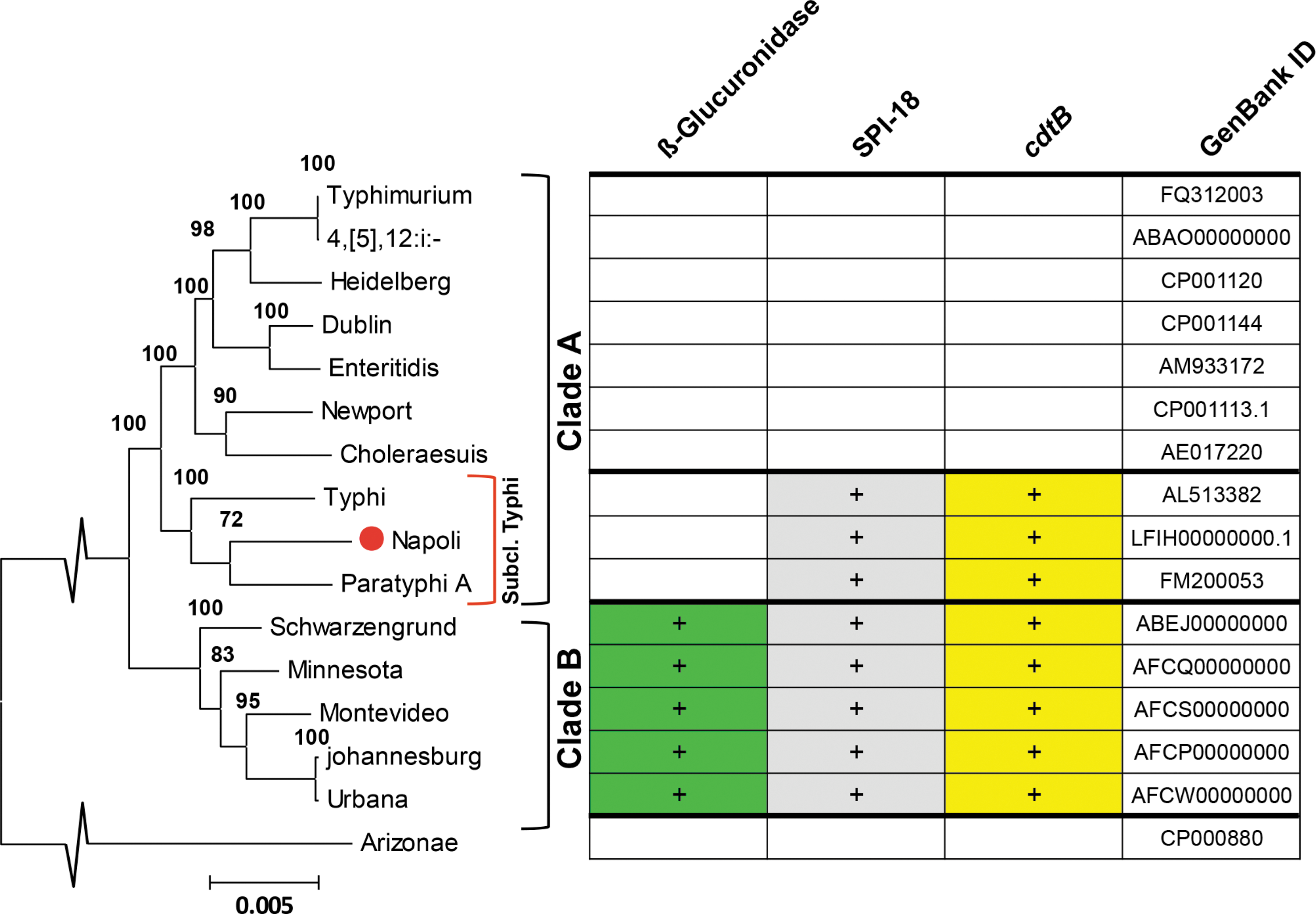
Phylogenetic relationships of 15 Salmonella enterica subsp. enterica serovars and one S. enterica subsp. arizonae isolate based on sequence data for 93 core-genome genes (den Bakker et al., 2011) using 200 bootstrap replications. The percentage of trees in which the associated taxa clustered together is shown above the branches. The blocks to the right of the phylogenetic tree indicate the presence of the β-glucuronidase operon (green), SPI-18 (gray), and the cdtB gene (yellow) in their genomes.
A total of 10,690 cases of NTS occurred in Lombardy during 2010–2014. Total cases, mean annual incidence, and mean annual increment of the top five serotypes causing NTS are reported in Table 1. The annual number of isolates significantly decreased from 2,441 in 2010 to 1,834 in 2014 (mean annual incidence decrement of −8.5%, p < 0.05). During this period, the only serotypes that showed an increment of NTS reports were 4,[5],12,:i:- (from 416 cases in 2010 to 713 in 2014) and Napoli (from 122 cases in 2010 to 151 in 2014). Remarkably, S. Napoli has reached the third position in 2014, overtaking S. Enteritidis (119 cases) (Figure 2). Nonetheless, a statistically significant increasing annual trend was only observed for S. 4,[5],12,:i:- (mean of 9.4%, p < 0.05).
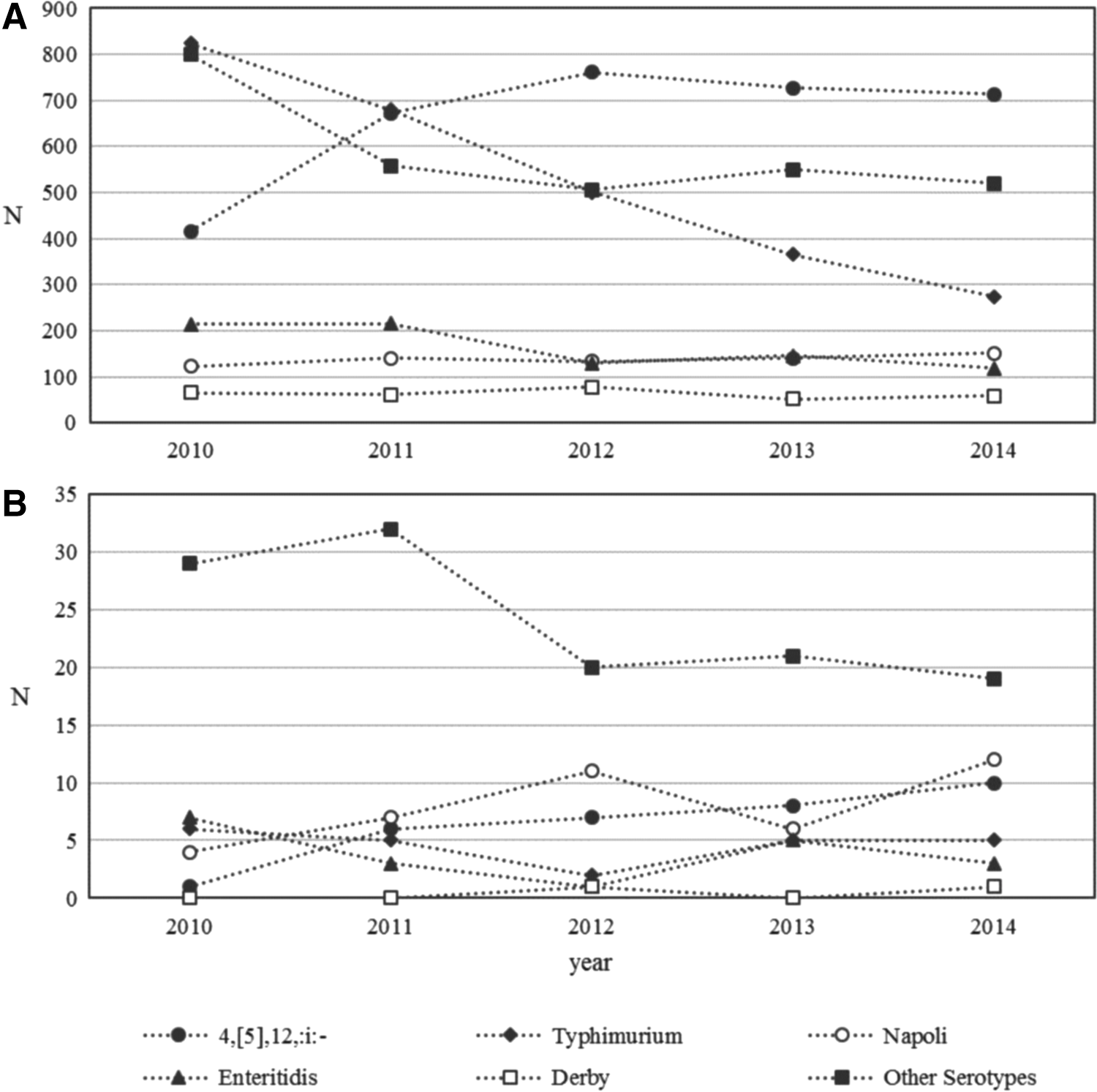
Total cases of
Mean of incidences (cases per 100,000 inhabitants) of all years (2010 to 2014).
Mean annual increment (%) estimated using a Poisson model.
Invasive index = (invasive infections/total cases) × 100; each invasive index is compared with the invasive index of all the other serotypes using a χ2 test.
p < 0.05.
NTS, nontyphoidal salmonellosis; iNTS, invasive nontyphoidal salmonellosis.
Of all NTS cases, 237 (2.2%) corresponded to iNTS (Table 1). From these, 116 (48.9%) were caused by the top five serotypes. The inter-annual trend of total iNTS remained stable. Among the top five serotypes, S. 4,[5],12,:i:- and S. Napoli showed an increment of iNTS notifications. iNTS caused by S. 4,[5],12,:i:- increased from 1 case in 2010 to 10 cases in 2014. S. Napoli iNTS reports increased from 4 cases in 2010 to 12 in 2014. As for NTS, only S. 4,[5],12,:i:- showed a statistically significant increase in the iNTS annual trend (mean of 37.3%, p < 0.05) (Figure 2).
Most importantly, during this period, S. Napoli was the first cause of iNTS affecting 40 cases (16.9% of total iNTS) out of 687, with an invasive index of 5.8% (95% CI, 4.2–7.8), which is more than twofold higher to that observed for the other nontyphoidal serotypes (2.0%, p < 0.001) (Table 1).
One limitation of this analysis is that all cases had been considered sporadic, because ENTER-NET database does not discriminate between outbreak-related and sporadic salmonellosis. Therefore, we could not determinate the exact number of cases derived from outbreaks in different years for all serotypes analyzed. The only outbreak that we could detect through our routine genotyping analysis was that caused by S. Napoli recently in Milan (Huedo et al., 2016) and no further PFGE clusters were identified among S. enterica isolates during the period under study. However, such possibility cannot be discarded, because not all isolates from the cases notified in the ENTER-NET database were available for genotyping.
Given the elevated incidence and invasive capacity observed for the serotype Napoli, we aimed to investigate its phylogenetic classification using a large multilocus sequence analysis comprising 93 core-genome genes and covering circa 73 Kb (den Bakker et al., 2011). Our phylogenetic analysis (Fig. 1) revealed that S. Napoli belongs to Typhi subclade in clade A with a strong bootstrap support (100%), Paratyphi A being the most related serotype (bootstrap value 72%). Likewise, the genome of S. Napoli carries the cdtB/SPI-18 combination and the putative virulent cluster composed of genes STY3343-3346. As for typhoid and clade B serotypes, the genome of S. Napoli carries the sapABCDF operon interrupted by the GICT18/1 islet (den Bakker et al., 2011). Contrary to clade B and in agreement with typhoid serotypes, S. Napoli lacks the uncommon β-glucuronidase operon (den Bakker et al., 2011).
From the recent S. Napoli outbreak investigation (Huedo et al., 2016), it has been noted that infections caused by S. Napoli and typhoid serotypes (Crump et al., 2015) present important similar characteristics. For instance, both cause prolonged incubation periods (7–14 days), common typhoid symptoms including high fever and bacteremia, and elevated rates of hospitalizations. In addition, both typhoid and S. Napoli serotypes require a very low infective dose to cause infection (Greenwood and Hooper, 1983; Crump et al., 2015).
Although our work is restricted to a limited geographical area and only one genome has been studied in detail, our results strongly suggest that S. Napoli and typhoid serotypes are highly related. More in-depth epidemiological and phylogenetic studies will be needed to further validate such relation. Meanwhile, the emergence of S. Napoli should be taken into serious consideration and its nontyphoidal classification could be discussed.
Footnotes
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.