
Abstract
Small ruminants have been implicated in outbreaks of Escherichia coli O157:H7 at livestock exhibitions throughout the United States. Additionally, goat meat or milk may serve as a reservoir for foodborne transmission of the organism. These associations highlight the public health importance of an effective strategy to reduce E. coli O157:H7 shedding in goats. We examined the efficacy of the SRP
Introduction
E
Numerous intervention strategies have been evaluated for reducing E. coli O157:H7 in cattle (Jacob et al., 2009; Cull et al., 2012; Matthews et al., 2013). These interventions are most often evaluated as a preharvest food safety strategy and few have been evaluated for use in goats. Zhang et al. (2014) evaluated a constructed recombinant vaccine using a caprine model; however, the availability of this vaccine for routine use is not known. In the United States, a commercially available vaccine targeting highly conserved siderophore receptor and porin proteins (SRP® vaccines) in E. coli O157:H7 has been extensively evaluated for use in cattle (Fox et al., 2009; Thomson et al., 2009; Thornton et al., 2009; Cull et al., 2012). The efficacy of this vaccine in goats has not been described and it is not marketed for use in this species. The use of vaccines to reduce the burden of E. coli O157:H7 in goats at livestock exhibition events may be a viable strategy to control public health risk.
The objective of our study was to determine the efficacy of the SRP vaccine against E. coli O157:H7 in goats housed in a setting similar to a petting zoo or fair. Secondarily, we aimed to determine if the persistence of E. coli O157:H7 strains previously associated with human outbreaks was different in orally challenged goats and if strain impacted vaccine efficacy.
Materials and Methods
Treatment groups and housing
This study was approved by the NC State University IACUC. Fourteen 6-month-old male goats (mean body weight = 22.7 kg) were randomly assigned to two treatment groups, vaccinated (n = 7) and unvaccinated (n = 7). The number of goats per treatment was determined a priori based on previous reports of vaccine efficacy (Fox et al., 2009; Thomson et al., 2009) and the availability of appropriate housing. Goats in the vaccinated group received a series of three injections with the E. coli Bacterial Extract Vaccine with SRP technology (Zoetis, Florham Park, NJ). Each dose (1 mL; subcutaneous) was administered 21 days apart (days −72, −51, and −29).
Before oral challenge with E. coli O157:H7 (day 0), both vaccinated and unvaccinated goats were cohoused on pasture and supplemented with free-choice Rumilab® (LabDiet, St. Louis, MO) for 66 days. During this time, eight goats were treated for respiratory disease using florfenicol (0.13 mL/kg; Merck Animal Health, Madison, NJ) or tulathromycin (0.024 mL/kg; Zoetis, Florham Park, NJ); however, the last dose was given 50 days before oral challenge. Approximately 1 month before challenge, goats were moved into pens (2.08 × 1.5 m) within a biosafety level-2 facility. Each pen was randomly assigned with one vaccinated and one unvaccinated animal. A plastic partition eliminated contact between pens. All animals had access to fresh water and were fed a forage-based diet, supplemented with Rumilab.
Vaccine response
Before the first vaccination (day −72) and at euthanasia (day 21), blood samples were collected from all animals. Additionally, blood samples were collected from animals in the vaccinated group just before the third vaccination (day −29). Serum (1.5 mL) was isolated through centrifugation (10,000 × g for 5 min) and sent on ice to Epitōpix (Willmar, MN) for enzyme-linked immunosorbent assay to assess serological response. The receiving laboratory was blinded to animal treatment group assignment and results were reported back as Sample:Positive ratios and categorized as adequate (S:P value 0.5–0.79), good (S:P value 0.8–1.0), or high (S:P value >1.0) as per their recommendations. The assay is used to determine if an animal had been exposed to E. coli O157 SRP found in the vaccine.
Screening for E. coli O157
Goats were screened twice (days −22 and −29) for wild-type E. coli O157 before oral challenge. Briefly, 1 g of feces was collected through rectal palpation and transported to the laboratory in a sterile Whirlpack bag (Nasco, Fort Atkinson, WI). The feces were enriched in 9 mL of Gram-negative broth containing cefixime (0.5 mg/L), cefsulodin (10 mg/L), and vancomycin (8 mg/L; GNccv broth). The broth was incubated for 6 h at 37°C, following which immunomagnetic bead separation (Dynal, Inc., New Hyde Park, NY) was performed as per manufacturer's instructions on one milliliter of enrichment. Fifty microliters was then plated onto sorbitol-MacConkey plates (Hardy Diagnostics, Santa Maria, CA) with cefixime (0.5 mg/L) and potassium tellurite (2.5 mg/L) and incubated overnight at 37°C. Gray colonies were restreaked onto Columbia agar with 5% sheep blood (BAP; Remel, Lenexa, KS) and incubated overnight at 37°C; all colonies were subjected to indole (Remel) and latex agglutination, using the O157 antigen (Oxoid; Remel).
Oral inoculation of goats
All goats were orally dosed with a cocktail of four outbreak strains of E. coli O157:H7 (obtained from the North Carolina State Public Health Laboratory) made resistant to 50 μg/mL nalidixic acid (NalR E. coli O157:H7). Isolates were positive for eae, fliC, rfbE, and stx2 and negative for stx1 using a previously described PCR (Bai et al., 2010); all four isolates exhibited distinct pulsed-field gel electrophoresis (PFGE) banding patterns. Briefly, isolates were streaked for isolation onto BAP and grown overnight at 37°C. A single colony from each isolate was selected and placed into 10 mL of tryptic soy broth (TSB; Difco; BD, Sparks, MD) for overnight incubation at 37°C. One milliliter of inoculated TSB was transferred into 100 mL of TSB and incubated at 37°C for 7 h. To determine the starting concentration after incubation, each strain was serially diluted 1:10 in phosphate-buffered saline (PBS) and spread plated in triplicate on sorbitol-MacConkey agar with cefixime (0.5 mg/mL), potassium tellurite (2.5 mg/L), and nalidixic acid (50 μg/mL; ctn-SMAC) and incubated at 37°C overnight. From the remaining starting inoculum, 25 mL of each strain was pooled together and 1 mL of the pooled sample was serially diluted 1:10 in PBS and assessed for concentration as described above. Five milliliters of the pooled NalR E. coli O157:H7 inoculum was then added to 100 mL of 1% sterile skim milk (Oxoid, Basingstoke Hampshire, United Kingdom) for orogastric inoculation of each animal. The final concentration of NalR E. coli O157:H7 dosed was 6.7 × 107 CFU/goat.
Goats were fasted for 16 h before inoculation. An orogastric tube was placed into the rumen and the inoculum was administered through a funnel and tubing. Both the tubing and funnel were rinsed with sterile water between each animal.
Sample collection and detection of NalR E. coli O157:H7
Post oral challenge, feces were collected each Monday, Wednesday, and Friday for 21 days. During sampling, personnel wore Tyvek suits (Tyvek; DuPont, Washington, DE), boot covers, bouffant cap, and nitrile gloves. Gloves were changed between animals and boot covers were changed between pens. Feces were collected through rectal palpation and placed into sterile Whirlpack bags. All samples were transported to the laboratory immediately after collection in a cooler for processing.
In the laboratory, feces were homogenized through mortar and pestle. Samples were weighed and placed in 9 mL of GNccv broth. After vortexing, 1 mL of the fecal enrichment was serially diluted 10-fold in PBS. One hundred microliters of each dilution was spread plated in triplicate onto ctn-SMAC plates. The remaining GNccv served as a secondary enrichment that was incubated overnight at 37°C. The following day, ctn-SMAC plates were enumerated to determine the concentration. Three isolated colonies from each fecal sample were randomly selected, restreaked onto BAP, confirmed by indole and agglutination to be E. coli O157, and stored in brain-heart infusion broth (Becton Dickinson, Sparks, MD) with 25% glycerol for further characterization. If fecal samples had no growth, then the secondary enrichment was plated onto ctn-SMAC. The secondary enrichment plates were incubated overnight at 37°C and confirmed as positive or negative for NalR E. coli O157:H7, as described above.
On day 21, all goats were humanely euthanized and necropsied. Contents from the rumen, cecum, colon, and rectum were collected into sterile Whirlpack bags and the concentration of NalR E. coli O157:H7 was determined following the methods above for feces. The rectum was incised to reveal the rectoanal junction (RAJ) mucosal. Visible fecal contents were gently washed away and the RAJ was thoroughly swabbed with a sterile foam-tipped applicator (VWR, Radnor, PA, Greenquist et al., 2005). The RAJ swabs were enriched in 9 mL of GN ccv and processed as described for feces.
Pulsed-field gel electrophoresis
Freezer stocks of individual NalR E. coli O157:H7 from each goat were plated back onto BAP and incubated overnight at 37°C in preparation for PFGE. Isolates were subjected to restriction enzyme digestion using XbaI for 2 h at 37°C and processed according to the PulseNet protocol (Ribot et al., 2006). Salmonella Braenderup H9812 was included as a control strain. Gel Doc XR (Bio-Rad, Hercules, CA) was used to image the gels and PFGE banding patterns were normalized and evaluated using BioNumerics software (Applied Maths, Inc., Austin, TX). Isolates displaying 100% Dice similarity were considered to be the same PFGE type.
Statistical analysis
NalR E. coli O157:H7 were enumerated from plates and the concentration calculated based on sample weight and dilution. All counts were log10 transformed. If direct plating yielded no growth and the secondary enrichment was positive for NalR E. coli O157:H7, the lowest detectable concentration was used. The environmental (oral cavity, pen feed pan, watering unit, and pen toy) and RAMS swab collected at necropsy were classified as either positive or negative for NalR E. coli O157:H7 and no concentration was determined. Our primary outcomes of interest were the number of days each goat remained positive for NalR E. coli O157:H7 and the log10 concentration of NalR E. coli O157:H7 in vaccinated and unvaccinated goats. A t-test was used to determine if there was a difference in the number of days goats remained positive between vaccinated and unvaccinated treatments. A repeated-measures analysis, allowing for a random goat effect, was used in SAS (v. 9.4, Cary, NC) to determine treatment effect on the concentration of E. coli O157 shed. Statistical significance was considered at p < 0.05.
Results
Immune response to SRP vaccination
Before vaccination, all goats (n = 14) were negative for SRP antibody production against E. coli O157:H7. After vaccination, all goats in the vaccinated treatment group had an S:P ratio in the adequate, good, or high response category (Table 1). At the termination of the study, all vaccinated animals had at least an adequate S:P ratio, with three animals categorized as high, three as good, and one as adequate. The seven unvaccinated animals had no apparent immune response to the SRP vaccine and were considered negative.
The S:P response was not calculated for nonvaccinated animals on day −29.
ND, not determined.
Detection of NalR E. coli O157:H7
All feces collected before oral challenge were negative for wild-type E. coli O157:H7. Following oral challenge, all goats shed NalR E. coli O157:H7 in their feces; the mean concentration was 5.7 log10 CFU/g on day 1. Beginning on day 6, not all animals had detectable NalR E. coli O157:H7 in the feces. The last day a positive fecal was detected was day 10 (two goats). The number of days that goats were fecal positive for NalR E. coli O157:H7 was not different (p = 0.42) between vaccinated and unvaccinated goats (Fig. 1). Additionally, the concentration of NalR E. coli O157:H7 shed on each study day was not different between vaccinated and unvaccinated goats (p = 0.57; Fig. 2), although numerically vaccinated goats always had a lower concentration. Within the group of vaccinated goats, there was no apparent relationship between goats' immunological classification (high, good, or adequate response) and their shedding pattern (Fig. 3); however, these associations were not evaluated statistically. No necropsy samples were positive for NalR E. coli O157:H7.
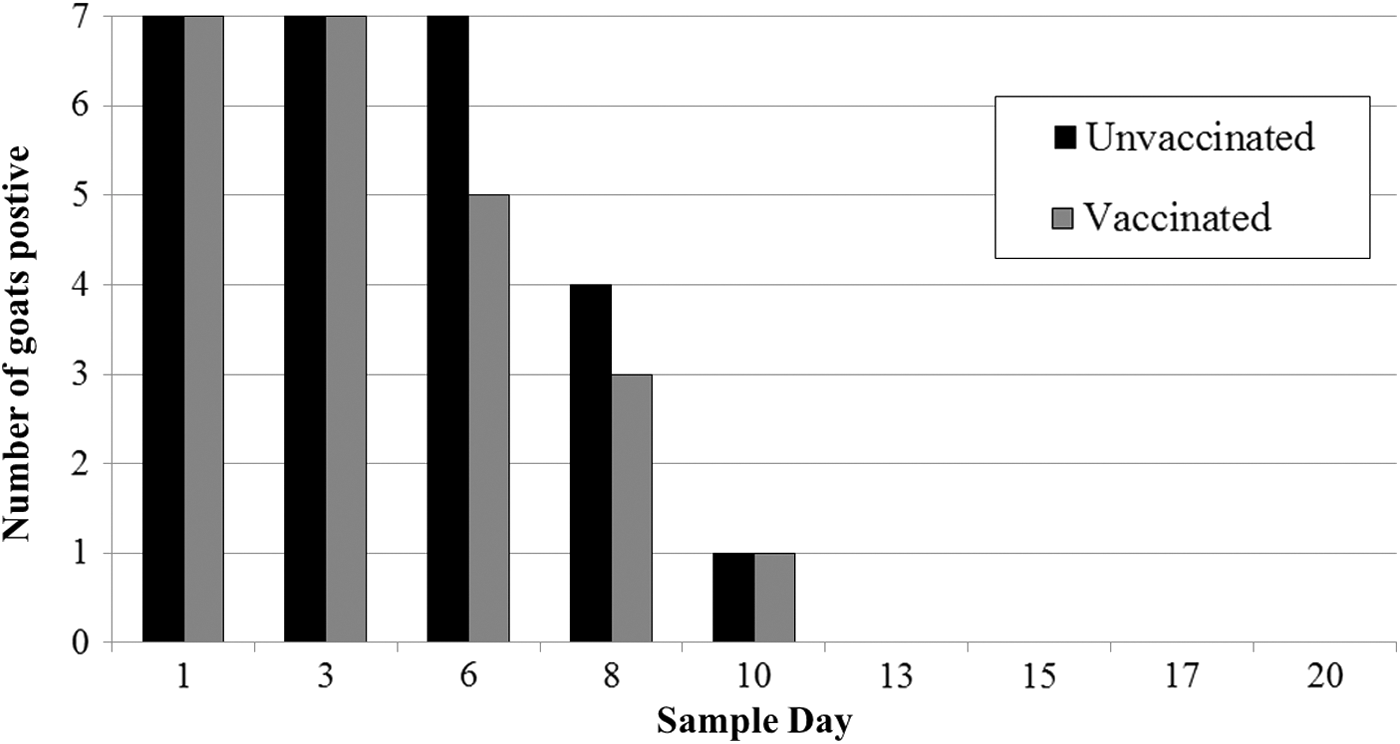
The number of vaccinated (n = 7) and unvaccinated (n = 7) goats that were fecal positive for NalR Escherichia coli O157:H7 on each sampling day postchallenge with a four-strain cocktail of E. coli O157:H7.
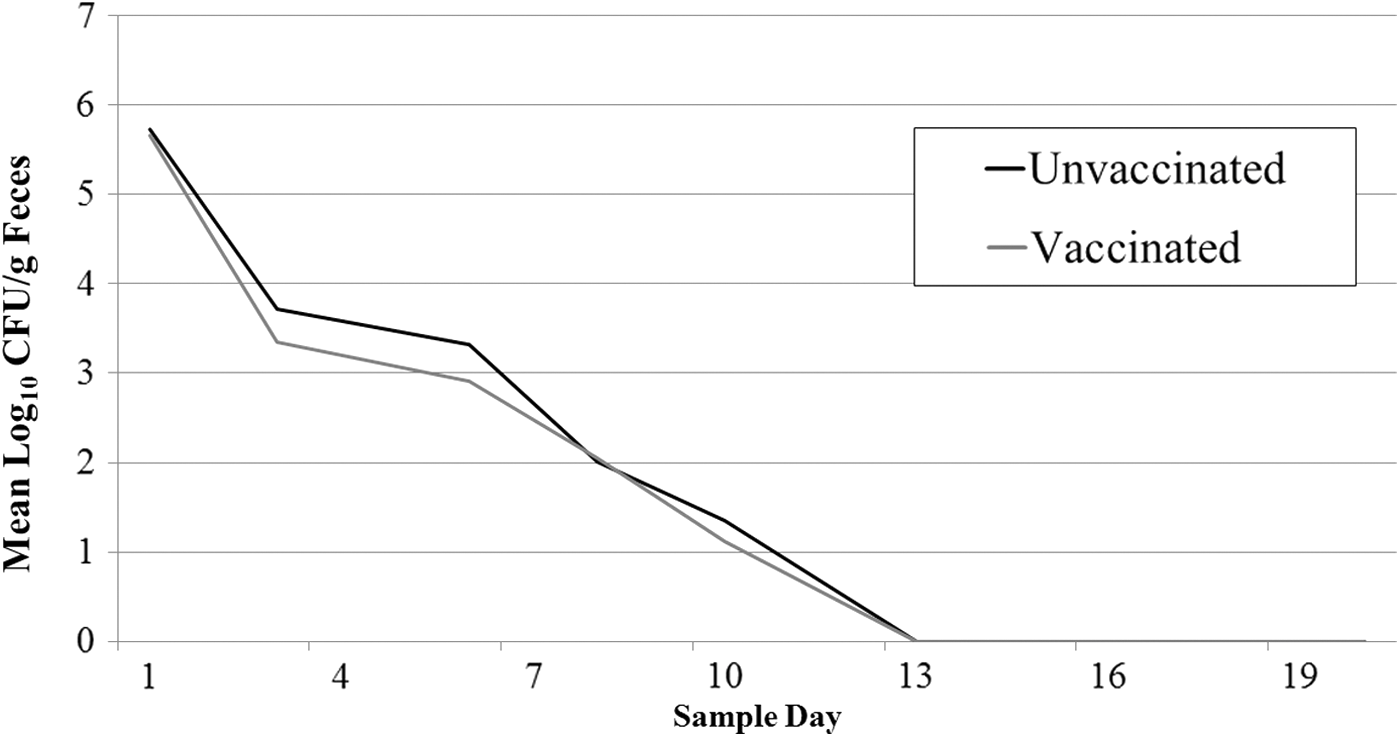
The mean concentration of NalR E. coli O157:H7 shed in feces of vaccinated (n = 7) and unvaccinated (n = 7) goats orally challenged with a four-strain cocktail of E. coli O157:H7.
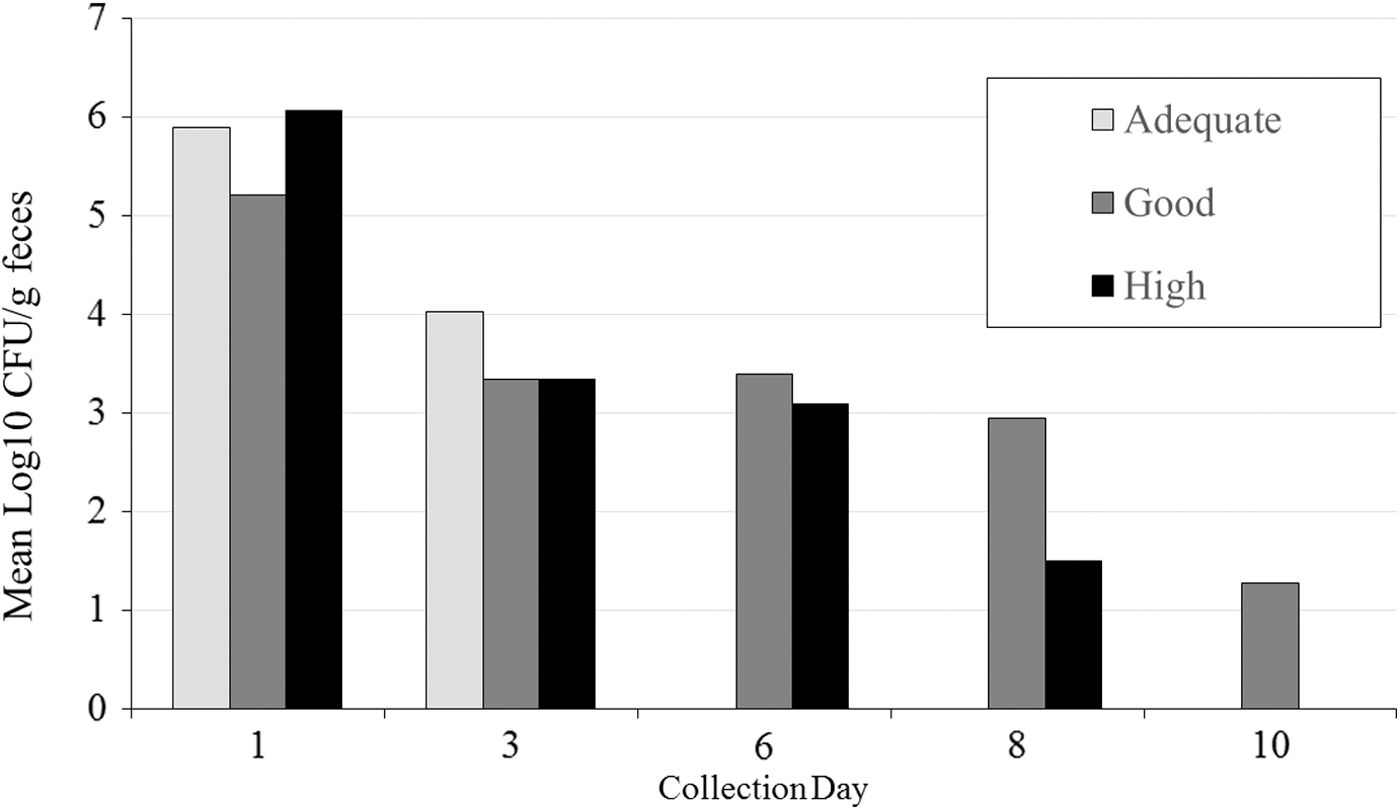
The mean concentration of NalR E. coli O157:H7 shed in vaccinated goats categorized as having adequate (n = 1), good (n = 3), or high (n = 3) immunological response to vaccination.
Pulsotype diversity of recovered E. coli O157:H7 isolates
All four challenge strains of E. coli O157 were detected in the feces from orally challenged goats. Strain 2 was recovered from 38% (52 of 137) of isolates, followed by strain 1 (28%; 39 or 137), strain 3 (18%, 24 of 137), and strain 4 (16%, 22 of 137). The distribution varied by day, with 89% (41 of 46) of strain 3 and strain 4 isolates recovered at the first collection, and only five isolates recovered in the rest of the study. Comparatively, only one isolate from strain 1 and no strain 2 isolates were recovered on day 1; however, these strains dominated the recovery after day 1. The number of isolates from each strain type and the total number of isolates collected from each sampling day are in Table 2. All goats shed multiple NalR E. coli O157:H7 strains during the study duration, and the average and median number of strains detected per goat per day was 1.6 and 1, respectively (range 1–3). At four collection points, feces from one goat yielded three different challenge strains. The proportion of each strain detected in the vaccinated and unvaccinated treatments is in Figure 4.
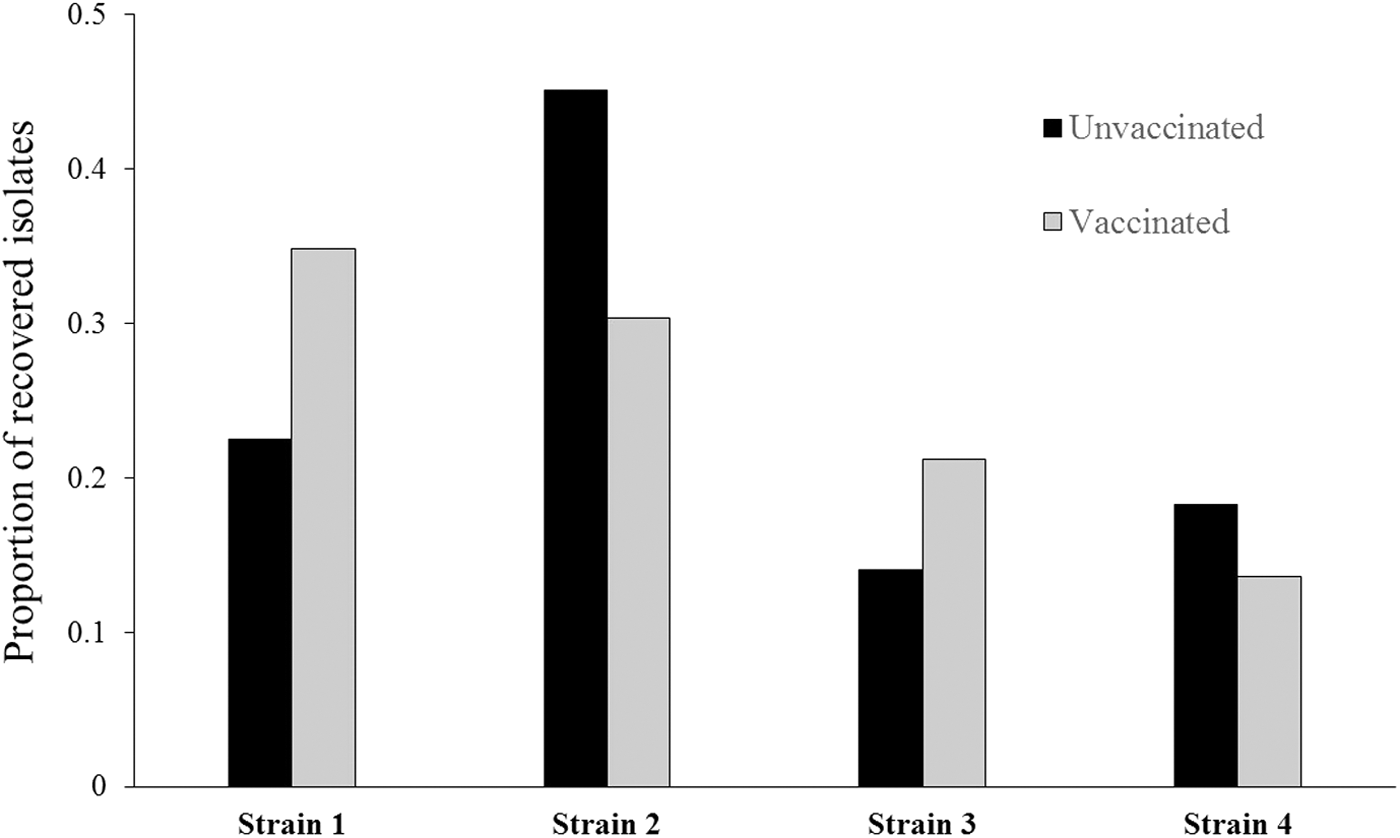
The proportion of strains recovered from vaccinated and unvaccinated goats orally challenged with a four-strain cocktail of E. coli O157:H7.
Discussion
Transmission of E. coli O157:H7 from healthy livestock to humans by direct contact at petting zoos or fairs has been reported (Durso et al., 2005; Goode et al., 2009). Intervention strategies aimed at reducing pathogen presence and concentration in livestock participating at these exhibitions or events may reduce the public health risk. Currently, control measures for pathogen spread at livestock exhibitions involve the physical separation of animals from the public (e.g., roped-off areas or limiting access) and implementation of hand washing stations and signage (Erdozain et al., 2015). Still, the proportion of visitors practicing hand hygiene remains low (Erdozain et al., 2013). This poor compliance results in the increased potential for transmission of zoonotic pathogens, including E. coli O157:H7.
Previously, vaccination has been shown to reduce the E. coli O157:H7 burden in cattle (Fox et al., 2009; Thomson et al., 2009; Cull et al., 2012). We hypothesized that this same commercially available vaccine would reduce shedding in goats. The evaluated product is currently labeled for use in cattle, and because there were no dosing instructions for use in goats, we consulted with the vaccine manufacturer and administered half of the labeled dose (1 mL) to 6-month-old goats. We did use the labeled vaccination schedule of three administrations, 21 days apart. It is unlikely that this decreased dose impacted the shedding of E. coli O157:H7 in goat feces in this study as all vaccinated goats had adequate immune responses.
Despite goats developing an immunological response to the vaccination, we found no significant differences in the concentration or the number of days in which NalR E. coli O157:H7 was detected from the feces of vaccinated and unvaccinated goats. Numerically, the mean concentration of E. coli O157:H7 shed in the feces was lower in vaccinated animals on all days but one; however, the difference was never more than 0.4−log10 E. coli O157:H7/g feces. Admittedly, the sample size for this study was small (seven animals per treatment); however, it was justified based on previous reports of vaccine efficacy (Fox et al., 2009; Thomson et al., 2009) and our desire to evaluate this intervention in an environment consistent with small animal livestock exhibitions or petting zoos. Certainly, an increased number of enrolled goats would make any treatment effect, if there was one, more apparent. However, in many goat-exhibiting or public events, the number of goats present is not large and a more consistent and dramatic vaccine effect would be expected to be needed.
Interestingly, goats in this study did not exhibit long-term shedding of E. coli O157:H7. It is well understood that experimental challenge studies do not necessarily reflect normal E. coli O157:H7-shedding patterns in cattle; however, this model has precedent to evaluate the potential of an intervention. It is possible that with longer shedding durations, more dramatic impacts of the vaccine could be observed. Certainly, by the termination of the study, goats were no longer colonized as E. coli O157:H7 was not detected from any sample type, including a swab of the rectoanal mucosal junction. The E. coli O157:H7-shedding dynamics in goats is not well described, certainly not as well as it is described in cattle. Further work in this area may improve the utility of experimental models using goats. To optimize our study, we inoculated goats with strains previously isolated from humans and submitted to the North Carolina Public Health Laboratory; these isolates were presumed to be associated with a goat-implicated outbreak. A previously described experimental challenge model in cattle showed that the frequency, concentration, pattern, and duration of E. coli O157 shedding were strain dependent (Gautam et al., 2012; Kulow, 2012). We also observed distinct shedding patterns in our study, where initially two strains of E. coli O157:H7 were most detectable, followed later by a shift to the other strains. This appeared to be largely independent of the individual animal. We observed no apparent differences in the shedding of strains between vaccinated and unvaccinated goats.
Conclusions
Although mitigation or intervention strategies to control pathogen shedding are needed for goats in livestock exhibition areas, we found little efficacy of the SRP E. coli vaccine for reducing E. coli O157 in orally challenged goats. Future studies to optimize the vaccine schedule for use in goats or identify other intervention opportunities are justified.
Funding
This work was supported by the USDA National Institute of Food and Agriculture, Animal Health Project accession number 1003363.
Footnotes
Disclosure Statement
No competing financial interests exist.