
Abstract
Human norovirus is the leading cause of foodborne illness worldwide with the majority of outbreaks linked to fresh produce and leafy greens. It is essential that we thoroughly understand the type of relationship and interactions that take place between plants and human norovirus to better utilize control strategies to reduce transmission of norovirus in the field onto plants harvested for human consumption. In this study the expression of gene markers for the salicylic acid (SA) and jasmonic acid (JA) plant defense pathways was measured and compared in romaine lettuce (Lactuca sativa) and Arabidopsis thaliana Col-0 plants that were inoculated with Murine Norovirus-1, Tulane Virus, human norovirus GII.4, or Hank's Balanced Salt Solution (control). Genes involving both the SA and JA pathways were expressed in both romaine lettuce and A. thaliana for all three viruses, as well as controls. Studies, including gene expression of SA- and JA-deficient A. thaliana mutant lines, suggest that the JA pathway is more likely involved in the plant immune response to human norovirus. This research provides the first pieces of information regarding how foodborne viruses interact with plants in the preharvest environment.
Introduction
H
Human noroviruses account for 60% of all leafy green associated foodborne outbreaks (DeWaal and Bhuiya, 2007; Herman et al., 2008).While it is recognized that many outbreaks are caused by contaminated food handlers, it is also recognized that contamination can occur in the field. One major route of contamination of foods with human enteric viruses is through fruits and vegetables irrigated with fecally contaminated water or by direct contact with fecal material through the use of manure and biosolids in the field (Sair et al., 2002). Transmission of human pathogens to plants can also occur through use of manure in the field, as well as contaminated irrigation water and hands, which have been documented under both laboratory and field conditions (Sair et al., 2002; Endley et al., 2003; Stine et al., 2005).
Human norovirus has also been associated with contaminated groundwater, surface water, and recreational water and can survive in groundwater for more than 18 months (Cheong et al., 2009). It has been previously demonstrated that human norovirus, as well as norovirus surrogates, can be internalized into leafy greens through the roots and foliar surfaces of lettuce plants (Wei et al., 2010a, b; Hirneisen and Kniel, 2013; Wang and Kniel, 2015) although it is not known how often this occurs naturally in the field and how internalization physically affects the plant or the viruses themselves. It is essential that we thoroughly understand the type of relationship and interactions that take place between plants and human norovirus to better develop control strategies to reduce transmission of human norovirus in the field onto plants harvested for human consumption.
It is also important to determine the type of relationship that human pathogens have with plants, whether it be endophytic or antagonistic. For example, it was suggested by Schikora et al. (2008) that human pathogenic Salmonella Typhimurium can infect and intracellularly proliferate tissues of Arabidopsis thaliana through both the root and shoot of the plant. Salmonella infection also led to disease symptoms in the plants, including wilting, chlorosis, and death of infected plant organs (Schikora et al., 2008). Arabidopsis plants responded immunologically similarly to Salmonella as they would to the plant pathogen Pseudomonas syringae (DC3000) by induction of the mitogen activated protein kinase (MAPK) cascades, as well as enhanced expression of pathogenesis related (PR) genes (Schikora et al., 2008).
Plants can also be infected by bacteria normally pathogenic to humans and other mammals, including Salmonella enterica, Pseudomonas aeruginosa, Burkholderia cepacia, Erwinia spp., Staphylococcus aureus, Escherichia coli O157:H7, and Listeria monocytogenes (Plotnikova et al., 2000; Prithiviraj et al., 2005; Milillo et al., 2008; Haapalainen et al., 2009; Holden et al., 2009). Most of the studies described above have been performed on A. thaliana because the entire genome of the plant has been sequenced and much is known in regards to how this plant responds to plant pathogens physically and immunologically. However, questions remain regarding whether or not A. thaliana serves as a good model for the study of human pathogen–plant interactions.
Another study by Klerks et al. (2007) found that lettuce plants responded to the presence of Salmonella enterica serovar Dublin at physiological and molecular levels shown through stunted plant growth and the expression of pathogenicity-related genes. Other studies have found that E. coli O157:H7 induced significantly higher PR-1 gene expression in A. thaliana plants compared to Salmonella enterica serovar Typhimurium suggesting that plants may recognize and respond to some human pathogens differently than others (Roy et al., 2013).
While there are very few studies looking at the plant defense response of human bacterial pathogens, there are none regarding human pathogenic viruses. The mechanism by which the plant immune system operates is through the detection of conserved microbial components (Jones and Dangl, 2006) referred to as microbe associated molecular patterns (MAMPs) or pathogen associated molecular patterns (PAMPs). The plant physiological responses to plant infection can be due to activation of the induced systemic resistance (ISR) or systemic acquired resistance (SAR) pathway.
The SAR pathway is induced if the pathogen is able to elicit a hypersensitive reaction (HR) (Enyedi et al., 1992; Durrant and Dong, 2004), which causes apoptosis in the plant to rid itself of the infectious agent. These events may enable the plant to become more resistant to future attacks by the pathogen (Ross, 1961; Madamanchi and Kuc, 1991). When a plant is invaded by a pathogen, one of two defense signaling pathways is activated: a salicylic acid (SA) dependent pathway or a SA-independent pathway involving jasmonic acid (JA) or ethylene (ET) signaling molecules (Kunkel and Brooks, 2002). Genes in the SA pathway are also used as markers to study induction of plant resistance by viral pathogens, including studies regarding tobacco and the tobacco mosaic virus (Chivasa et al., 1997). JA-dependent and ET-dependent plant defenses are generally activated by necrotrophic pathogens and chewing insects, whereas SA-dependent defenses are often triggered by biotrophic pathogens (Pandey and Somssich, 2009).
The purpose of this study was to determine how the human enteric pathogen human norovirus interacts with plants in the preharvest environment. We also sought to determine if plants respond differently to human norovirus, as well as its widely used surrogates (murine norovirus [MNV] and Tulane virus [TV]). In addition, we wanted to determine if plant model A. thaliana and Romaine lettuce (Lactuca sativa) elicited an immune response toward human noroviruses and if so, which plant defense pathways were utilized by the plants. To accomplish this, the expression of gene markers for the SA and JA plant defense pathways was measured and compared in romaine lettuce (L. sativa) and A. thaliana Col-0 plants that were inoculated with MNV-1, TV, human norovirus GII.4, or Hank's Balanced Salt Solution (HBSS) (control).
Materials and Methods
Seed germination and cultivation
A. thaliana, in addition to romaine lettuce, was used as a model for plant defense studies toward noroviruses. A. thaliana seeds of the wild-type Colombiana (Col-0), as well as mutant lines nahg and jin1, were generously provided by Dr. Harsh Bais (University of Delaware, Newark, DE). Mutant nahg plants are deficient in the ability to induce production of SA. Mutant jin1 plants are insensitive to the production of JA.
A. thaliana and romaine lettuce (Paris Island cultivar) (Johnny's Selected Seeds, Waterville, ME) seeds were sterilized in a 50% (25,000 ppm) sodium hypochlorite solution and vortexed for 1 min then rinsed thrice with sterile deionized water.
Following sterilization, seeds were planted in individual containers containing sterile promix soil (square containers 4 × 3.5 × 4.5 cm in dimension; T.O. Plastics, Ontario, Canada). Holes were created in the bottom of each individual container, which was then placed into plastic bins inside a growth chamber.
The growth chamber temperature conditions were monitored daily, and temperature was maintained at 20°C ± 0.30°C, relative humidity was 60% ± 2.08%, and the water activity (Aw) of the promix was maintained at 1.046 ± 0.04. Romaine lettuce plants were utilized for all experiments once they reached 4 weeks of age, and A. thaliana plants were used at 2 weeks of age.
Virus inoculation and sample collection
Experiments were performed by inoculating romaine lettuce, A. thaliana Col-0, nah, as well as jin1 mutant plants, with either human norovirus surrogates, including MNV-1 and TV, or human norovirus GII.4 Sydney strain. A positive control using a plant pathogen was not able to be included due to restrictions on the use of such pathogens, as well as permit requirements.
Information regarding immune response and gene expression of A. thaliana in response to inoculation by plant pathogens has been widely published (Enyedi et al., 1992; Dong, 2004; Durrant and Dong, 2004; De Vos et al., 2005; Jones and Dangl, 2006; Haapalainen et al., 2009). A. thaliana nahg mutant plant lines are SA deficient and were used to evaluate the role of PR-1 as a genetic marker for the SA pathway in the immune response of A. thaliana Col-0 to human norovirus and its surrogates. A. thaliana jin1 mutant lines are JA deficient and were used to evaluate the role of PDF1.2 and VSP2a as genetic markers for the JA pathway in the immune response of A. thaliana Col-0 to human norovirus and its surrogates. Plants were also inoculated with HBSS as controls. HBSS was selected as a control as viral stocks inoculated onto plants were suspended in HBSS (Hyclone, Logan, UT).
Because plant viruses are known to enter plant cells through cellular damage (Mandadi and Scholthof, 2013), lesions (1 cm) were made on leaves of plants before inoculation using a sterile pipette tip. Plants with lesions only (no virus) were used as controls and for normalization of gene expression data.
Concentrated virus in aliquots of 50 μL (107 plaque-forming unit (PFU)/mL MNV; 104 PFU/mL TV; 109 genomic copies/mL GII.4) was applied onto plant leaves where lesions were made using a sterilized paint brush to avoid runoff of inoculum. For experiments, including romaine lettuce, three leaves per plant were pooled into one sample. Due to their small size, the leaves of three whole A. thaliana plants were also pooled into one sample.
Each experiment was performed in triplicate and replicated at least twice (n = 6). Samples were collected immediately following viral inoculation (0+) or at 6, 12, or 24 hours postinoculation (hpi). Plant tissues were immediately frozen in liquid nitrogen and ground up using a mortar and pestle. Samples were stored at −80°C until further analysis was performed.
Plant RNA extraction and real-time polymerase chain reaction
Plant RNA was extracted from frozen plant tissues using the ZR Plant RNA MiniPrep Kit (Zymogen Research, Irvine, CA). An in-column DNase digestion was performed using DNAse I (Zymogen Research, Irvine, CA) according to the manufacturer's instructions, and RNA purity and concentration assessment were performed using a NanoDrop (Thermo Scientific, Waltham, MA). Before cDNA synthesis, RNA was diluted to 500 ng in a 10 μL total volume. Synthesis of cDNA was performed using the Life Technologies High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific, Waltham, MA). Reverse transcription master mix was prepared according to the manufacturer's instructions. Thermal cycling parameters were as follows: 25°C for 10 min, 37°C for 120 min, and 85°C for 5 min followed by a hold at 4°C using the Eppendorf Mastercycler (Eppendorf, NY).
Primer design for plant defense genes in romaine lettuce
Primers were designed for romaine lettuce gene markers involved in the plant defense pathways. Sequence homologs for PR-1 and NPR-1 primers in romaine lettuce were designed from known A. thaliana nucleotide sequences using the NCBI Nucleotide BLAST database (tbastx) (NCBI, NIH) (Table 1). PR-1 and NPR-1 served gene markers involved in the SA pathway of plant defense. Primers for genes involved in defense pathways in A. thaliana included PR-1, PDF1.2, and VSP2a (Table 2). PDF1.2 and VSP2a served as gene markers for the JA/ET pathway of plant defense. A. thaliana primers were generously provided by Dr. Harsh Bais (University of Delaware, Newark, DE). Efficiency of primers was confirmed through polymerase chain reaction (PCR) and visualization of products using gel electrophoresis.
Accession numbers of sequences from GenBank through NCBI nucleotide BLAST.
SA, salicylic acid.
Accession numbers of sequences from GenBank through NCBI nucleotide BLAST (Altschul et al., 1990).
ET, ethylene; JA, jasmonic acid; SA, salicylic acid.
Quantitative PCR and calculation of relative gene expression
The quantitative PCR (qPCR) master mix was made using 10 μL of 2 × QuantiTect SYBR Green PCR Master Mix, 2 μL of forward and reverse primers (10 mM), 4 μL of RNase-free water, and 2 μL of cDNA template to each sample. Cycling parameters were as follows: PCR initial activation at 95°C for 15 min followed by 40 cycles of 94°C for 15 s, 60°C for 30 s, and 72°C for 30 s. Data were acquired from the SYBR Green channel at the extension step (72°C). The ΔΔ Cq calculation method was utilized to determine relative gene expression as described by Haimes and Kelley (2014) using actin (ACT) as an endogenous reference gene in lettuce and ubiquitin (UBQ) as an endogenous reference gene in A. thaliana. Expression levels of target genes from plants induced with lesions only (no virus) were used for normalization of the gene targets. Reproducibility of the assay was confirmed by conducting qPCR with the same samples on two different days obtaining the same results.
Statistical analysis and interpretation of results
For all experimental methods, experiments were performed in triplicate (n = 6) with two trials performed on different days. A one-way ANOVA was performed to compare means within the data set (Student's t-test) using JMP software (SAS Institute Inc., Cary, NC). All p-values ≤0.05 (α = 0.05; CI = 95%) were considered statistically significant.
Results
Romaine lettuce and A. thaliana elicit increased plant defenses in the presence of MNV, TV, and GII.4
The relative gene expression of plant defense genes in romaine lettuce and A. thaliana plants inoculated with MNV, TV, or GII.4 was compared to that of plants inoculated with HBSS. It is important to note that there were no physical symptoms or stressors observed in plants throughout the duration of the experiment, such as chlorosis or wilting.
In romaine lettuce, MNV-1 did not significantly induce expression of PR-1; however MNV did significantly induce the expression of NPR-1 at 6, 12, and 24 hpi compared to HBSS inoculated controls (Fig. 1). This indicates that romaine lettuce plants elicit an immune response toward MNV through induction of NPR-1 and the SA pathway of defense.
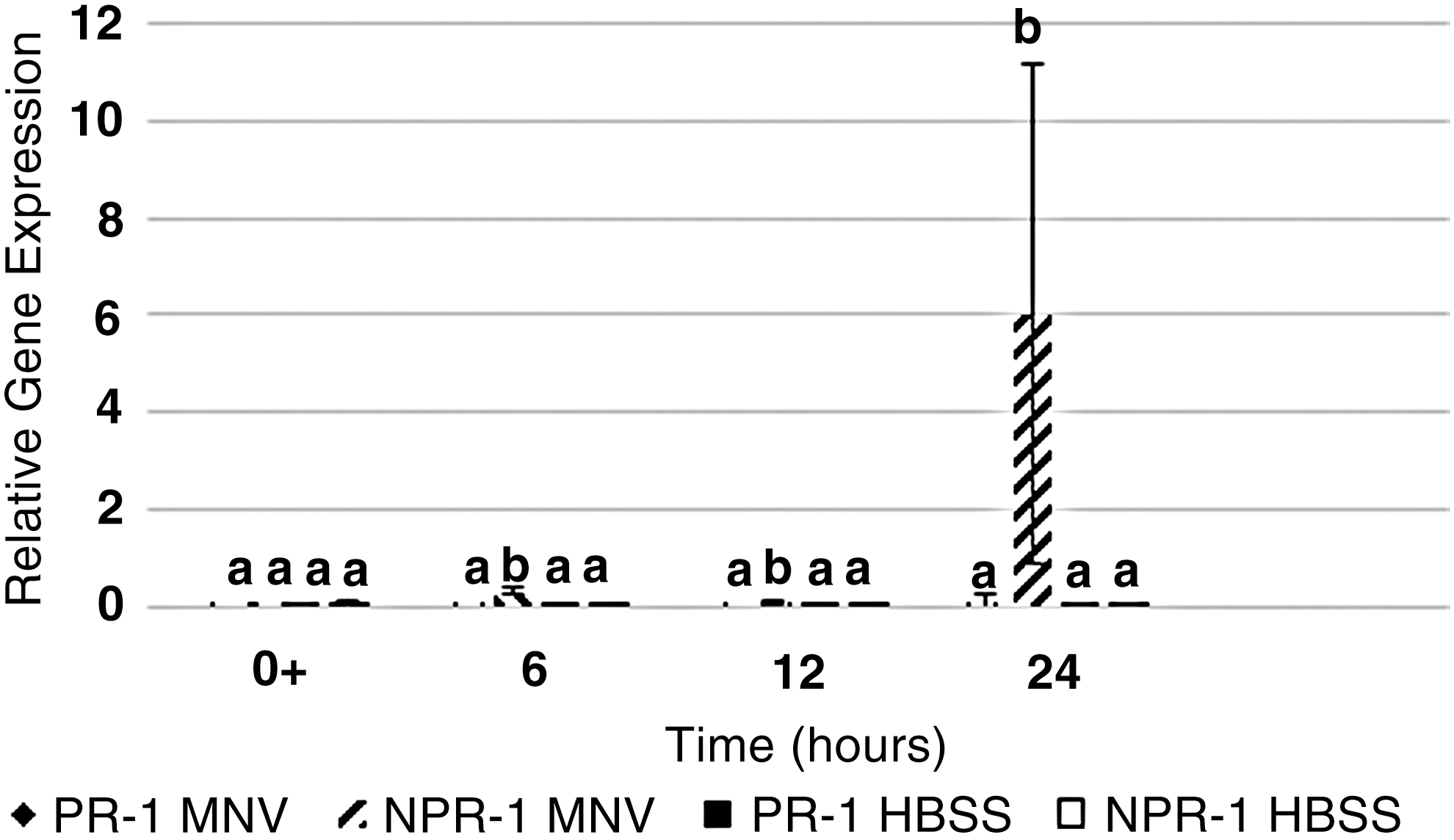
Relative gene expression of the pathogenesis-related 1 gene (PR-1) and the nonpathogenesis-related-1 (NPR-1) gene in romaine lettuce inoculated with MNV or HBSS (control) was plotted as a function of time. Samples were collected immediately after inoculation (0+) and at 6, 12, and 24 hpi. Error bars were calculated as the standard deviation between the mean of the relative gene expression between experimental replicates (n = 6). Bars not labelled with the same letter are significantly different (α = 0.05). HBSS, Hank's Balanced Salt Solution; hpi, hours postinoculation; MNV, murine norovirus.
Inoculation of TV onto romaine lettuce significantly induced PR-1 and NPR-1 at 6 hpi compared to controls (Fig. 2). Human norovirus GII.4 significantly induced PR-1 and NPR-1 in romaine lettuce at 6, 12, and 24 hpi (Fig. 3). Results indicate that human norovirus, as well as its surrogates, was able to induce an immune response in romaine lettuce through the induction of the SA pathway through the increased expression of PR-1 and NPR-1 compared to controls.
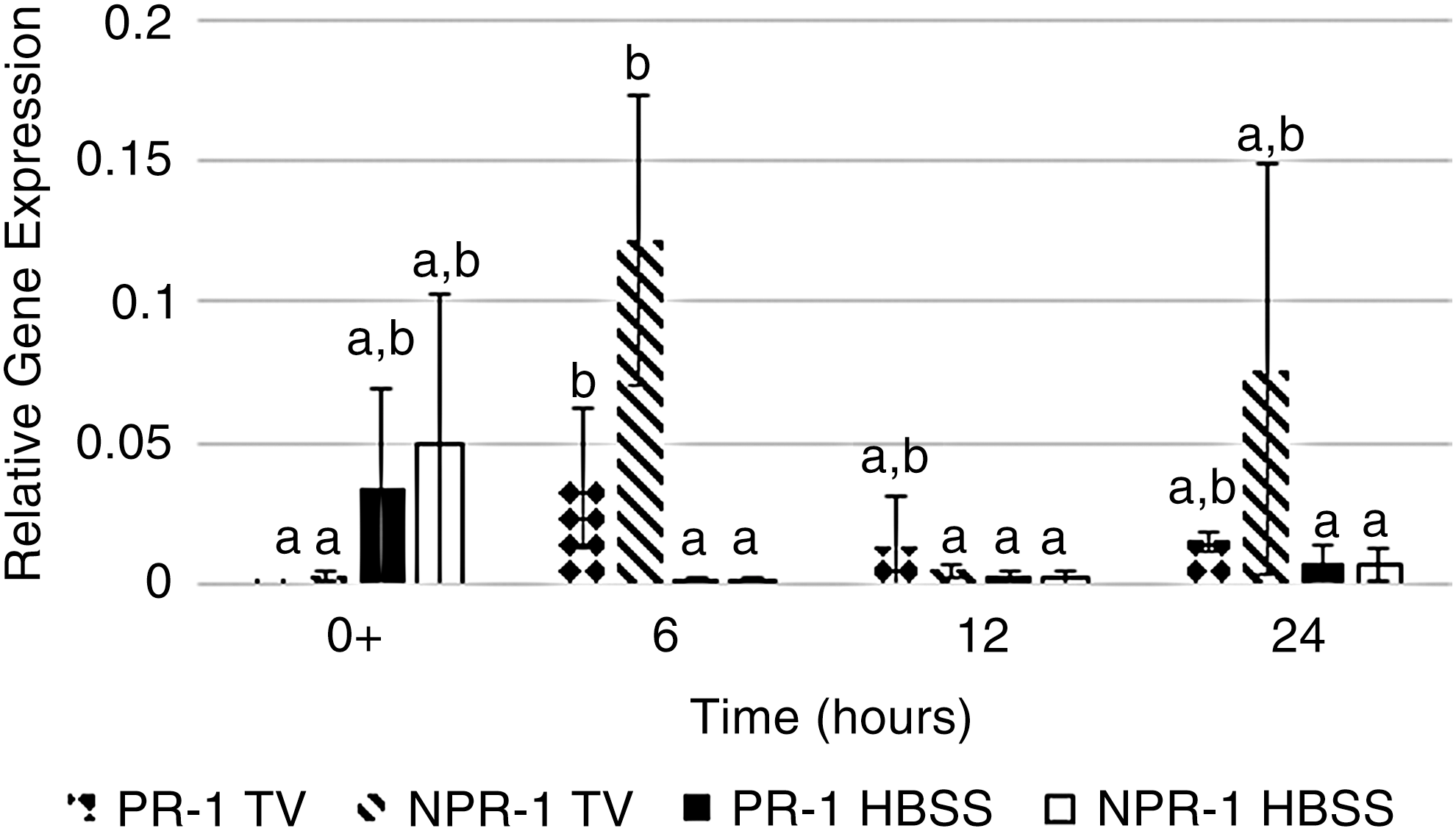
Relative gene expression of the pathogenesis-related 1 gene (PR-1) and the nonpathogenesis-related-1 (NPR-1) gene in romaine lettuce inoculated with TV or HBSS (control) was plotted as a function of time. Samples were collected immediately after inoculation (0+) and at 6, 12, and 24 hpi. Error bars were calculated as the standard deviation between the mean of the relative gene expression between experimental replicates (n = 6). Bars not labelled with the same letter are significantly different (α = 0.05). HBSS, Hank's Balanced Salt Solution; TV, Tulane virus.
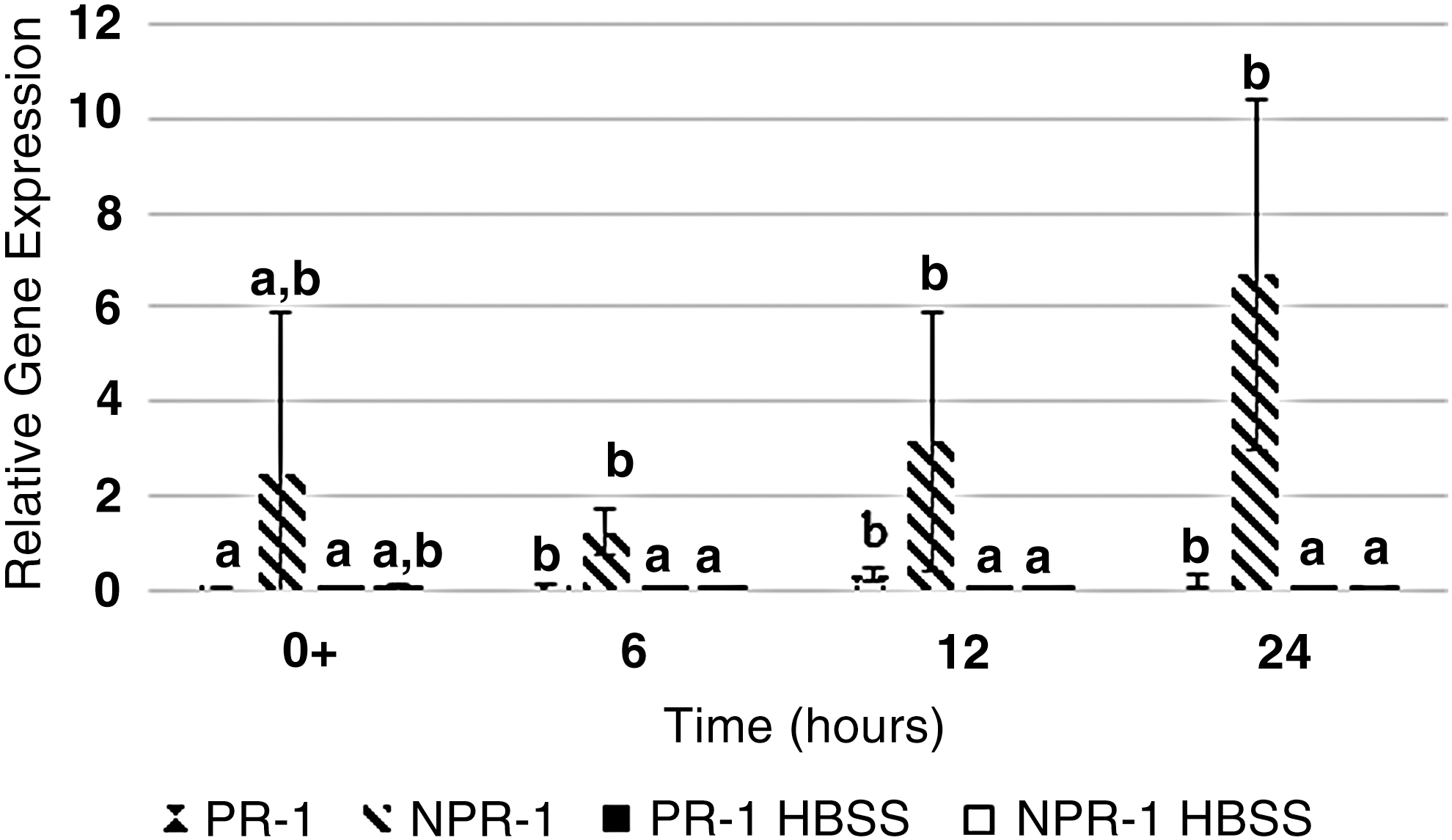
Relative gene expression of the pathogenesis-related 1 gene (PR-1) and the nonpathogenesis-related-1 (NPR-1) gene in romaine lettuce inoculated with human norovirus GII.4 or HBSS (control) was plotted as a function of time. Samples were collected immediately after inoculation (0+) and at 6, 12, and 24 hpi. Error bars were calculated as the standard deviation between the mean of the relative gene expression between experimental replicates (n = 6). Bars not labelled with the same letter are significantly different (α = 0.05). HBSS, Hank's Balanced Salt Solution.
For all viruses, NPR-1 was expressed significantly higher than PR-1 in romaine lettuce (p = 0.05). The highest expression levels were observed in lettuce plants inoculated with human norovirus GII.4, although this was not statistically significant. Average fold change in gene expression for PR-1 and NPR-1 on MNV, TV, and human norovirus GII.4 inoculated romaine lettuce at 0, 6, 12, and 24 h compared to HBSS (control) inoculated plants can be seen in Figure 4. Average fold change in gene expression was highest for NPR-1 in MNV and human norovirus GII.4 inoculated plants.
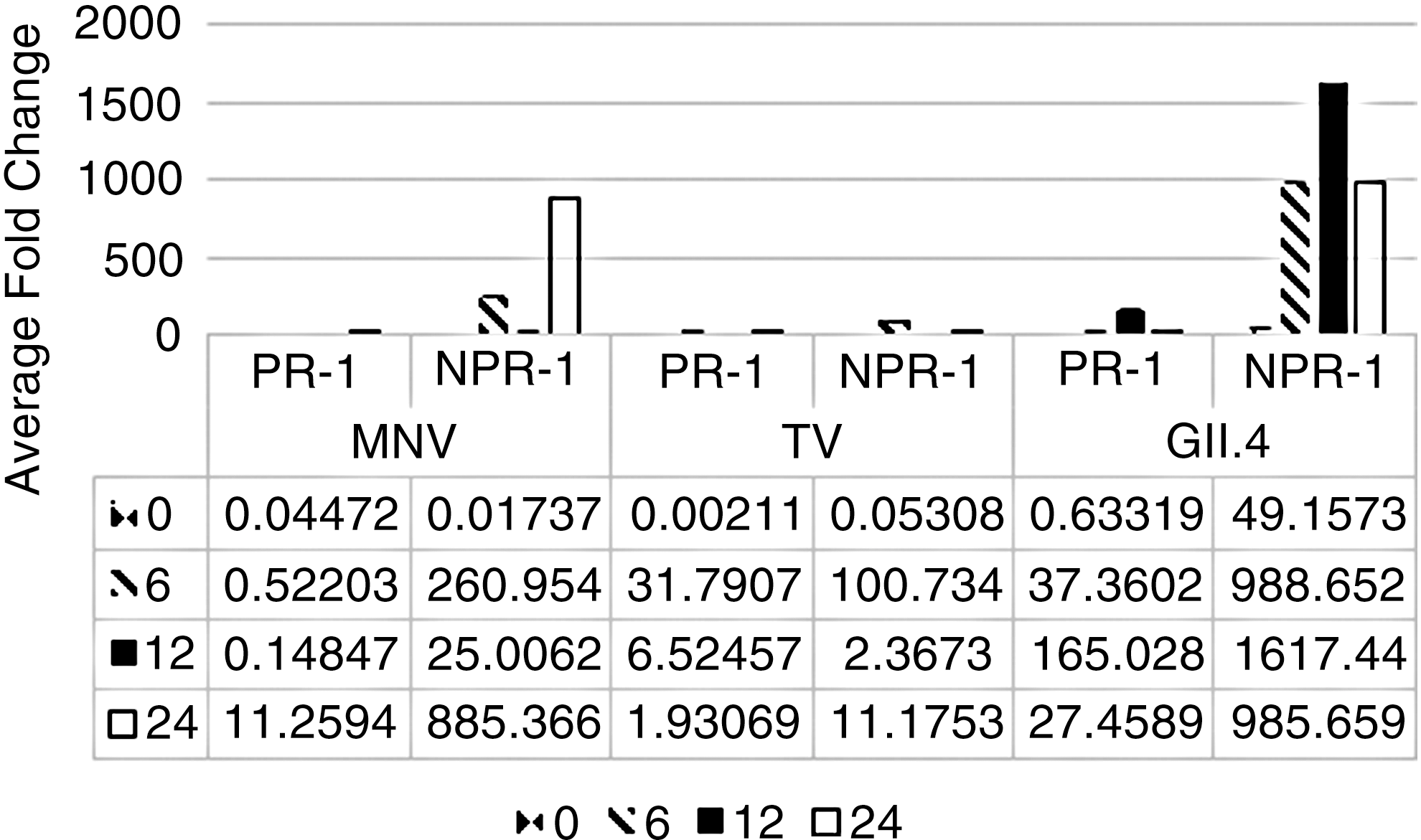
The average fold change in gene expression for virus inoculated romaine lettuce compared to HBSS treated controls plotted as a function of time. Samples were collected immediately after inoculation (0+) and at 6, 12, and 24 hpi. Error bars were calculated as the standard deviation between the mean of the relative gene expression between experimental replicates (n = 6). HBSS, Hank's Balanced Salt Solution.
In A. thaliana Col-0 plants, MNV significantly induced the expression of PR-1 and PDF1.2 at 6 and 24 hpi, respectively. VSP2a was significantly induced at 0+, 6, and 12 hpi (Table 3). Unlike in romaine lettuce, TV was not able to significantly induce expression for any of the genetic markers in A. thaliana. Human norovirus GII.4 was able to induce PR-1 expression in A. thaliana at 0+, 12, and 24 hpi, similar to that which was observed in romaine lettuce (Table 3).
Gene expression significantly higher in wild type as indicated by (+).
hpi, hours postinoculation; MNV, murine norovirus; TV, Tulane virus.
Human norovirus also significantly induced expression of PDF1.2 at 0+ and 12 hpi and VSP2a at 12 hpi. These results indicate that only MNV and human norovirus were able to significantly induce an immune response in A. thaliana. In both cases, gene markers in both the SA and JA pathways were significantly induced compared to controls. In A. thaliana, human norovirus GII.4 induced significantly higher levels of expression compared to MNV (p = 0.02) and TV (p = 0.01).
Gene expression in A. thaliana mutants confirms expression of plant defense genes in Col-0
The relative gene expression of PR-1 was determined in nahg mutant plants and compared to expression levels in wild-type (Col-0) plants to determine the role of PR-1 and the SA pathway in the plant immune response of A. thaliana to human norovirus and its surrogates. Wild-type plants (Col-0) inoculated with MNV and TV showed significant expression of PR-1 at 6 hpi compared to expression in mutant plants. There was no significant difference in the expression of PR-1 in wild-type and nahg mutant plants inoculated with human norovirus GII.4.
Wild-type plants inoculated with MNV and TV also showed similar expression patterns of PDF1.2 where significant expression was observed at 24 hpi for both viruses in wild-type plants compared to jin1 mutant plants. MNV and TV significantly induced VSP2a in wild-type plants at 6 and 12 hpi compared to jin1 mutant plants. Human norovirus GII.4 significantly induced expression of VSP2a in wild-type plants at 12 hpi compared to expression in jin1 mutant plants. Results are summarized in Table 3.
Discussion
In the current study, a significant increase in gene expression was observed for the NPR-1 gene only for MNV inoculated lettuce plants (Fig. 4). At 6 hpi in the current study a 0.52-, 31.79-, and 37.36-fold increase was observed in the PR-1 gene in addition to an even greater increase (260.95-, 100.73-, and 988.65-fold) in the NPR-1 gene for MNV, TV, and human norovirus GII.4 inoculated lettuce plants, respectively. Interestingly, gene expression studies in A. thaliana mutant studies demonstrated that JA pathway associated genes, including PDF1.2 and VSP2a, were also upregulated in MNV, TV, and human norovirus GII.4 inoculated A. thaliana plants.
In a previous study by De Vos et al. (2005) P. syringae DC3000 infection of A. thaliana induced a 60.4-fold increase in expression of the PR-1 gene (SA pathway) at 6 hpi and a 1.7-fold decrease in the PDF1.2 gene (JA pathway). In this case infection by the bacterial pathogen upregulated the SA pathway and downregulated the JA pathway. This was further confirmed by Spoel et al. (2003) who showed that JA responsive genes, including PDF1.2 and VSP, were suppressed upon Pst DC3000 infection of A. thaliana.
Studies investigating the immune response of susceptible A. thaliana plants to tobacco mosaic virus demonstrated that both PR-1 (39.29-fold increase) and PDF1.2 (1.44-fold increase) genes were upregulated at 6 hpi (Wang et al., 2009). Taken together, results of the current study are more similar to those of previous studies involving plant immune response toward plant viral pathogens where both genes involved in the SA- and JA-mediated pathways are upregulated.
In romaine lettuce, MNV did not significantly induce expression of PR-1; however MNV did significantly induce the expression of NPR-1 at 6, 12, and 24 hpi, which was somewhat surprising (Figs. 1 and 4). NPR-1 protein functions downstream of SA to mediate changes in expression of defense genes (Dong, 2004), so one would expect that PR-1 would simultaneously be induced because it is involved in stimulation of the SA defense pathway. However, depending on the pathogen type and stage of infection, NPR-1 also interacts with components of JA signaling (Mandadi and Sholthof, 2013).
It is possible that the increased expression of NPR-1 by MNV in romaine lettuce could be due to its interaction with the JA pathway. This can be further supported by the fact that wild-type A. thaliana plants demonstrated similar expression levels of gene markers related to the JA pathway (PDF1.2 and VSP2a) (Table 3). Based on these results, A. thaliana is able to recognize the presence of MNV; however, it does not recognize the virus as a biotrophic threat because PR-1 is not induced and HR is not observed in plants.
Inoculation with TV did not significantly induce any of the defense genes in A. thaliana (Figs. 2 and 4). According to A. thaliana mutant experiments, expression patterns are almost identical to that of MNV (Table 3). It is suspected that gene expression levels are lower from plants inoculated with TV compared to MNV and GII.4 because of the low titer level of TV stocks. It is very difficult to propagate a TV viral stock of greater than 105 PFU/mL in the laboratory. In this case, plants were inoculated with only 104 PFU/mL compared to 107 and 109 PFU/mL MNV and human norovirus, respectively. The high titer levels of human norovirus GII.4 inoculum may also explain the increased gene expression in romaine lettuce.
Results were similar for PR-1 expression in A. thaliana Col-0 where expression was significantly higher than controls at 0+, 12, and 24 hpi (Figs. 3 and 4). Expression of PDF1.2 was significant at 12 and 24 hpi, and VSP2a expression was significant at 12 hpi for A. thaliana Col-0 plants inoculated with human norovirus GII.4 (Table 3). Although PR-1 and NPR-1 expression was significant in both romaine lettuce and wild-type A. thaliana Col-0, according to A. thaliana mutant studies only VSP2a was significantly expressed in wild-type plants indicating the involvement of the JA pathway in the plant immune response to human norovirus GII.4. This is consistent with conclusions made from experiments with MNV.
It is very interesting that both romaine lettuce and A. thaliana plants responded differently to the three viruses. By definition, viruses do not possess conserved PAMP/MAMP-like features such as flagellin or chitin (Mandadi and Scholthof, 2013). However, structures, including, but not limited to, the viral capsid (virion), viral ribonucleoprotein complexes, and viral-encoded glycoproteins embedded on the host derived lipid membranes of plant rhabdoviruses (Goldberg et al., 1991; Jackson et al., 2005) are analogous to PAMPs/MAMPs (Mandadi and Scholthof, 2013). These viral structures are conserved among members of related virus taxa (Mandadi and Scholthof, 2013). Thus, it is hypothesized that such viral patterns are analogous to PAMPs/MAMPs and are accessible for recognition by membrane-bound receptor-like proteins to trigger immune responses in plants (Mandadi and Scholthof, 2013). It is possible that these structures vary among MNV, TV, and GII.4 explaining the differences in expression levels for the gene markers tested.
MNV and TV demonstrated different expression patterns in romaine lettuce and A. thaliana; however, gene expression patterns in A. thaliana mutant line studies were nearly identical. A similar pattern was observed with human norovirus GII.4 in romaine lettuce and A. thaliana. PR-1 was expressed in both romaine lettuce and A. thaliana Col-0; however, mutant line studies indicated that the only significant gene expression was related to VSP2a and the JA pathway.
While there is some argument in the field of food safety that A. thaliana does not serve as a good model to study the interactions between food plants and human pathogens, the results of this study indicate that A. thaliana should be used along with food (edible) plants to more clearly understand these interactions and how they can affect food safety and human health. Results from this study indicate that both romaine lettuce and A. thaliana demonstrate plant defense responses to human norovirus and its surrogates through both SA and JA mediated pathways.
Studies, including gene expression of SA- and JA-deficient A. thaliana mutant lines, suggest that the JA pathway is more likely involved in the plant immune response to human norovirus. It is likely that the plants are not recognizing noroviruses as plant pathogens and are instead recognizing conserved structural viral motifs that elicit a basal level of response through the JA pathway and induction of a systemic induced system response (ISR).
This study is the first of its kind to attempt to try and understand the type of interactions that occur between plants and viral human pathogens. Because the genome of romaine lettuce (L. sativa) has yet to be sequenced, we do not know the exact genes involved in their immune response pathways, as well as how romaine lettuce responds to plant pathogens or pests. Because of this limitation, we designed primers for plant defense genes that were homologous to those in A. thaliana for which the complete genome sequence is available.
Strict regulations regarding the use of plant pathogens in nonpermitted laboratories prevented the use of these pathogens in the present study. Instead our discussions were focused on comparing the observed plant–pathogen interactions between romaine lettuce and norovirus with the well-studied interactions between A. thaliana and P. syringae DC3000. Although there were several limitations as outlined above, we were able to design a study that has allowed us to begin to “scratch the surface” regarding plant–pathogen interactions, specifically those regarding viral human pathogens.
Conclusions
Romaine lettuce and A. thaliana demonstrated different expression levels of genetic markers involved in plant immune response to human norovirus and its surrogates (MNV and TV). While genes involving both the SA and JA pathways were expressed in both romaine lettuce and A. thaliana for all three viruses, studies, including gene expression of SA- and JA-deficient A. thaliana mutant lines, suggest that the JA pathway is more likely involved in the plant immune response to human norovirus.
This research provides the first piece of information regarding how foodborne viruses interact with plants in the preharvest environment. Ongoing research will address gene expression upon “natural” internalization of noroviruses through the roots of leafy greens, as well as effects of virus inoculation, on other immune response pathways. Further indicators of plant immune response should also be investigated, including callus deposition and hydrogen peroxide accumulation in the leaves of plants during a potential norovirus contamination event.
There are many knowledge gaps involving how plants respond to norovirus contamination, which is the number one cause of foodborne illness in the United States, with the majority of outbreaks involving leafy greens. Future studies should address the effects of the plant immune response on the persistence of human norovirus on plants. Such studies could aid in the development of preharvest interventions such as the development of plant breeding programs that will lead to varieties with enhanced plant defense mechanisms. Such interventions would provide sustainable and economically feasible methods for reducing contamination of plants by plant pathogens and human pathogens in the preharvest environment, including human enteric viruses.
Footnotes
Acknowledgments
The authors are grateful to Jan Vinje at the CDC for providing the human norovirus GII.4 Sydney strain. This work is supported, in part, by the United States Department of Agriculture, National Institute of Food and Agriculture project NoroCORE, award number 2011-68003-30395 and also by award number 2011-67018-30217.
Disclosure Statement
No competing financial interests exist.