
Abstract
Broiler chickens may serve as reservoirs for human colonization by vancomycin-resistant Enterococcus (VRE). We examined the effects of vancomycin and two commonly used antimicrobial feed additives on VRE colonization in broiler chickens during grow-out. Chicks received unsupplemented feed or feed containing vancomycin, chlortetracycline, or tylosin from day of hatch to grow-out at 6 weeks. At 3 days of age, chicks received by crop gavage 107 colony-forming units (CFUs) of a human or poultry VRE isolate. Cecal contents were monitored weekly for VRE, short-chain fatty acids (SCFAs), and bacterial denaturing gradient gel electrophoresis (DGGE) profile methods. Vancomycin promoted persistent and high-level colonization with human- and poultry-derived VRE to grow-out in comparison with controls, while treatment with chlortetracycline and tylosin did not. Colonization by the poultry isolate in control, chlortetracycline, and tylosin groups persisted throughout the grow-out period with low concentrations present at 6 weeks, whereas the human isolate decreased to an undetectable level by week 6. Vancomycin resulted in significant reductions in cecal acetic acid and butyric acid in comparison with controls, but chlortetracycline and tylosin did not. DGGE profiles contained two main clusters with all vancomycin profiles in a smaller cluster and all other profiles in a larger cluster. These results demonstrate that vancomycin, but not chlortetracycline or tylosin, disrupted the indigenous microbiota and SCFA patterns of broiler chickens and promoted colonization by VRE.
Introduction
E
Several European studies demonstrated that extensive VRE colonization of broiler chickens persisted for years in the absence of selective pressure exerted by avoparcin (Borgen et al., 2001; Heuer et al., 2002b; Bortolaia et al., 2015). In these studies, the fecal concentration of VRE in broilers was low with detection only by selective enrichment (Bortolaia et al., 2015). The microbiological basis for persistence of VRE in the absence of selective pressure is not clear. Because VRE strains are often resistant to multiple classes of antibiotics, one possible explanation is that selective pressure from other agents may contribute to persistence of VRE. Many classes of antibiotics that disturb the indigenous intestinal microbiota promote VRE colonization in mice and humans (Donskey, 2004), but little information is available regarding the effects of commonly used therapeutic and nontherapeutic antibiotics on the intestinal microbiota of food animals and it is not known if such treatments promote VRE.
The major objective of the current study was to examine the effects of vancomycin and two commonly used feed additive antibiotics, chlortetracycline and tylosin, on VRE colonization during grow-out of broiler chickens as experimental animals. We hypothesized that the antibiotic exposure would result in persistent VRE colonization for up to 6 weeks, a typical grow-out period before harvest.
Materials and Methods
Bacterial strains
Two vancomycin-resistant E. faecium strains were used. E. faecium C68 was a VanB VRE strain isolated from a patient in Cleveland, Ohio (Donskey et al., 1999). A poultry-derived E. faecium GRE13, a Van B isolate, was donated by Dr. Toni Poole, USDA, ARS, SPARC, FFSRU. C68 in combination with subcutaneous administration of antibiotics that inhibit intestinal anaerobes resulted in the establishment of persistent high-density stool colonization in mice (Donskey et al., 1999). Minimum inhibitory concentration (MIC) of vancomycin for C68 is 512 μg/mL (Donskey et al., 1999) and 256 μg/mL for GRE13 (Poole et al., 2004). MICs of tetracycline and erythromycin for C68 are >10 μg/mL (Donskey et al., 1999). GRE13 MIC for tylosin is 256 μg/mL and tetracycline and erythromycin >10 μg/mL (Poole et al., 2004).
Experimental model
Animal experiments were conducted in accordance with the relevant federal guidelines and institutional policies of the USDA-ARS Southern Plains Agricultural Research Center Animal Care and Use Committee Policy Statement. Separate experiments were performed with the human and poultry VRE strains. Analysis of droppings on crate liners from the hatchery demonstrated that chicks were VRE negative. Four hundred day-of-hatch broiler chicks (Sanderson Farms, Bryan, TX) were divided into four treatment groups: Group 1 received no treatment; Group 2 was given access to feed containing 50 g/ton of chlortetracycline (Fluka, Milwaukee, WI); Group 3 was given access to feed containing 50 g/ton of vancomycin (Sigma Chemical Company, St. Louis, MO); and Group 4 was given access to feed containing 50 g/ton of tylosin (Tylan 10; Elli Lilly, Indianapolis, IN). All chicks at 3 days old were inoculated by crop gavage with 107 CFUs of VRE in 0.5 mL of sterile phosphate-buffered saline. Chicks were provided free access to water and an unmedicated corn–soy bean meal-based ration that contained or exceeded the levels of critical nutrients recommended by the National Research Council (National Research Council, 1984). Vancomycin was studied rather than avoparcin because avoparcin is no longer available. Chicks were maintained on pine litter in separate experimental rooms with independent air and water systems. Rooms were cleared of used litter, cleaned with disinfectant and bleach, and remained empty for at least 3 weeks between subsequent experiments. Results from two experiments with the human isolate were consistent and were combined for analysis and presentation; a single experiment was conducted with the poultry strain.
Sample collection
Cecal contents (0.35 g) were collected each week over a 6-week grow-out period from 10 chicks in each group and mixed into 3.65 mL of sterile Butterfield's buffer (0.62 mM monobasic potassium phosphate, pH 7.2). Samples were serially diluted in sterile Butterfield's buffer for spread plating and enumeration. VRE CFUs/g were determined following plating onto Enterococcosel agar (Becton Dickinson, Sparks, MD) containing 20 μg/mL of vancomycin and incubation for 48 h at 45°C. The lower limit of detection for VRE was 10 CFUs/g of cecal contents. As the VRE cecal concentrations approached the limits of detection, 1 mL of sample volumes was transferred to 9 mL of tryptic soy broth, incubated at 42°C for 48 h, then streaked onto Enterococcosel agar containing 20 μg/mL of vancomycin, and incubated for 48 h at 45°C for confirmation. Sample short-chain fatty acid (SCFA; acetic, propionic, and butyric) concentrations were determined by gas liquid chromatography as described previously (Hume et al., 1993).
Denaturing gradient gel electrophoresis
Cecal bacterial diversity was determined by performing denaturing gradient gel electrophoresis (DGGE) of 16S rDNA gene polymerase chain reaction (PCR) amplicons as previously described by Muyzer et al. (1993) with modifications by Hume et al. (2003). Band pattern percentage similarity coefficient (%SC) analyses and dendrograms were constructed using Pearson product-moment correlation coefficient and unweighted-pair group method with arithmetic mean (UPGMA) options in GelCompare II 6.6 (Applied Maths, Inc., Austin, TX).
Statistical analyses
Statistical analyses were conducted with PC-SAS® (Version 8.2; commercial statistical software; SAS Institute, Inc., Cary, NC). Analysis of variance and Duncan's multiple range test were utilized where applicable to determine statistically significant differences at p < 0.05.
Results
Vancomycin-resistant Enterococcus
Vancomycin-treated chicks given the human isolate maintained high densities of cecal VRE throughout grow-out, whereas significant decreases (p < 0.05) in VRE density were seen in the control, chlortetracycline, and tylosin groups by week 1. None of the chicks in these groups other than the vancomycin group had detectable VRE by week 6 (Fig. 1). For the chicken isolate, vancomycin-treated chicks also maintained high densities (p < 0.05) of VRE throughout grow-out in comparison with the other groups. VRE decreased over time in the chlortetracycline, tylosin, and control groups; however, the chicken-derived VRE was not eliminated by grow-out in any of these groups.
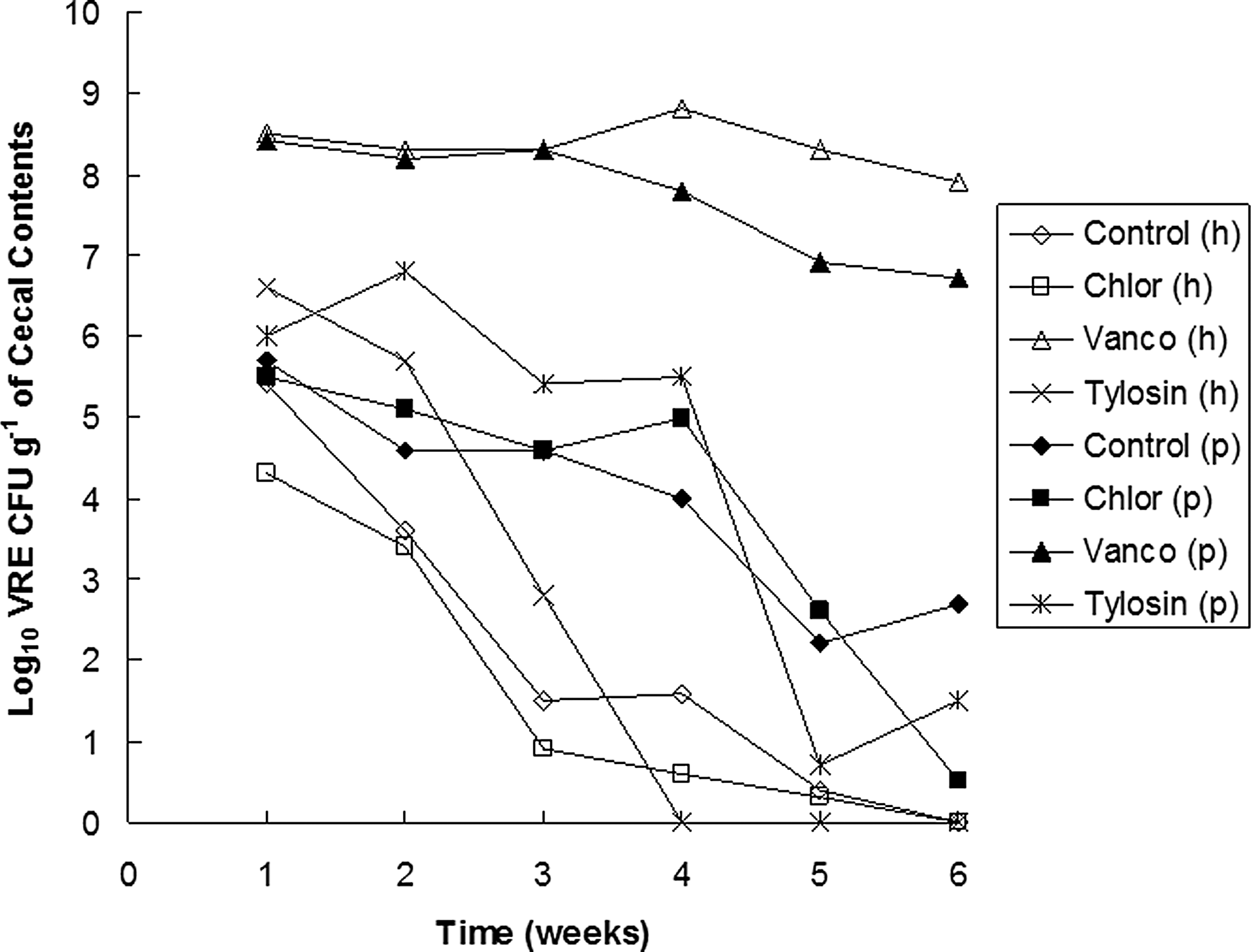
Cecal VRE during grow-out in broilers given VRE and antimicrobials. h = human isolate, n = 20; P = poultry isolate, n = 10. Chlortet, chlortetracycline; Vanco, vancomycin. VRE, vancomycin-resistant Enterococcus.
Short-chain fatty acids
Acetic, propionic, and butyric acids, as end products of intestinal bacterial metabolism, may indicate shifts in population constituents and proportions. For both the human and poultry VRE experiments, acetic and butyric acid concentrations were significantly lower (p < 0.05) in chicks given vancomycin in comparison with the other groups, whereas propionic acid levels were not significantly reduced (p > 0.05) by vancomycin treatment (Fig. 2). For all groups, the general pattern was that of increases in individual and total cecal SCFAs as chicks aged. Treatment with tylosin and chlortetracycline did not result in significant reductions in the levels of individual or total SCFAs in comparison with the controls (p > 0.05).
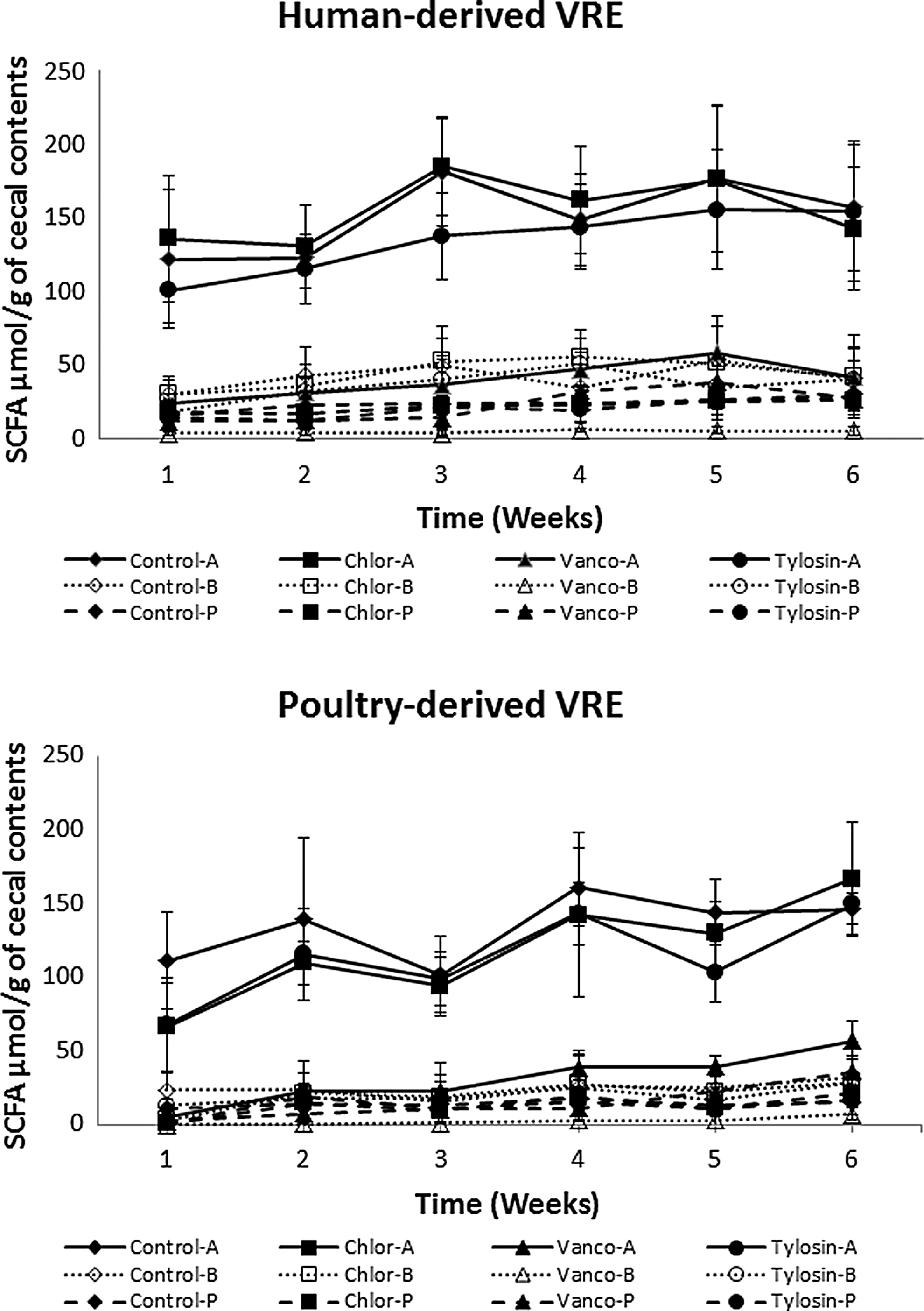
Cecal acetic, butyric, and propionic acids during grow-out in broilers given vancomycin-resistant Enterococcus faecium and antimicrobials. Means ± standard deviations, n = 20 with the human isolate, n = 10 with the poultry isolate. A, acetic acid; B, butyric acid; Chlor, chlortetracycline; P, propionic acid; SCFAs, short-chain fatty acids; Vanco, vancomycin; VRE, vancomycin-resistant Enterococcus.
Denaturing gradient gel electrophoresis
Bacterial 16S rRNA genes were amplified by PCR and DGGE profiles were developed to determine differences in cecal bacterial populations resulting from treatment with the antimicrobial feed additives. A representative dendrogram of an electrophoretic profile of cecal DGGE amplicons is presented in Figure 3. Cecal bacterial profiles from chicks treated with vancomycin formed one cluster with about a 75.6%SC for bacterial constituents over the 6-week grow-out. The vancomycin clade shared a 52.2%SC with the remaining treatment groups, indicating no similarity between the two clades. Despite the formation of vancomycin profiles in a single cluster, there was no apparent ordering of the profiles according to time during grow-out. Bacterial profiles from all the remaining treatment groups had about a 58.7%SC; week 2 tylosin was not included as a result of low DNA harvest. Generally, there were two larger clusters within the group that did not include vancomycin that were organized according to time during grow-out. Those two clusters had a 58.7%SC and were generally organized with weeks 1 to 3 having a 63.8%SC and weeks 4 to 6 a 70.4%SC. There was no overt clustering of individual treatment groups. However, there was a general clustering of control profiles with profiles from chicks given chlortetracycline. Furthermore, there was an apparent clustering of these two treatment groups in relation to time during grow-out. That is, week 1 control and chlortetracycline profiles formed a cluster, while similar clustering was seen with weeks 2 and 3 profiles and with weeks 5 and 6 profiles. An analysis of similarity between treatment groups on a weekly basis resulted generally in control and chlortetracycline groups showing closest similarity and the tylosin group forming the next level of similarity with this group and the vancomycin group having the least similarity to any groups. When analyzed individually, patterns for each group showed increased and progressive diversity over time (data not shown).

A representative DGGE dendrogram of band pattern percentage similarity coefficient (bar) comparisons of broiler chick cecal bacteria during the 6-week grow-out. Chlor, chlortetracycline; Con, control; DGGE, denaturing gradient gel electrophoresis; Tyl, tylosin; Van, vancomycin.
Discussion
The establishment of a healthy, competitive digestive microbiota for the prevention or elimination of enteropathogen (e.g., Salmonella and Escherichia coli) colonization is an accepted concept (Nurmi and Rantala, 1973; Nisbet et al., 1993; Corrier et al., 1995). Disruption of that protective microbiota weakens microbial defenses against enteropathogen colonization. Thus, it is important to understand the impact of antibiotics on the intestinal microbiota and on the risk for colonization by pathogens such as VRE. In the current study, we found that 3-day-old broiler chickens given crop inoculations of human- and poultry-derived VRE isolates, but no antibiotic treatment, developed cecal colonization that persisted, respectively, for 4–5 weeks or through to week 6 of the grow-out period. Vancomycin, but not chlortetracycline or tylosin, promoted persistent high-density intestinal colonization with the poultry-derived and human VRE strains.
Our results are consistent with previous studies in suggesting that selective pressure from glycopeptide antibiotics facilitated emergence and dissemination of VRE in Europe. Oral vancomycin has been shown to suppress anaerobic intestinal microbiota and promote persistent high-density colonization by VRE in mice and humans (Donskey et al., 1999, 2004), similar to our findings in chickens. However, our findings are also consistent with recent reports demonstrating that VREs often persist in broiler chickens in the absence of glycopeptide selective pressure (Borgen et al., 2001; Heuer et al., 2002b; Nilsson et al., 2009; Bortolaia et al., 2015). We demonstrated that VRE colonization persisted for several weeks in chickens in the absence of glycopeptide treatment, but that concentrations decreased to a low level by the end of the grow-out period. Similarly, in previous studies, the fecal concentration of VRE in broilers was low with detection only by selective enrichment (Bortolaia et al., 2015).
In contrast to our findings, some previous studies have reported that antimicrobial feed additives such as tylosin and chlortetracycline may cause significant changes in the composition of the microbiota of chickens (Torok et al., 2011; Lin et al., 2013). The differences may be related to use of different methods to analyze the microbiota or other differences in study design. In our study, it is important to note that the lack of impact of tylosin and chlortetracycline on DGGE patterns and on SCFA profiles correlates with the failure of these agents to promote colonization with VRE in comparison with nonantibiotic-treated controls. It is possible that these agents may cause alterations of the microbiota that can be detected using sensitive methods without altering the ability of the microbiota to provide colonization resistance against VRE.
Several factors may play roles in the pathogenesis of VRE colonization in chickens. First, our data suggest that disruption of the indigenous intestinal microbiota or exposure before establishment of a diverse microbiota may facilitate colonization by VRE. Although anaerobes play a critical role in inhibiting VRE colonization in mice and humans (Donskey, 2004), additional studies are needed to determine the component of the microbiota that inhibits VRE colonization in chickens. Second, overgrowth in the intestinal tract in the presence of vancomycin may be associated with extensive environmental contamination and repeated ingestion of VRE (Heuer et al., 2002b; Donskey, 2004). Such repeated exposures to pathogens may facilitate colonization even in the presence of an intact indigenous microbiota. In accordance with normal agricultural growth conditions, our chicks were maintained on the same litter from the beginning to the end of each trial. So, repeat inoculations from contact with contaminated litter likely occurred. However, in spite of repeated exposure, the human isolate reached undetectable levels by the end of grow-out in all but the vancomycin group. Culturing cecal contents in VRE enrichment broth and subsequent culturing on selective medium failed to detect the human VRE in these chicks at 6 weeks. Third, it has been suggested that VRE strains may exhibit host specificity with isolates from poultry and hospitalized patients falling into different amplified fragment length polymorphism genogroups (Willems et al., 2000). Host specificity could potentially explain the fact that the poultry-derived isolate persisted throughout the 6-week grow-out, while the human isolate did not. However, studies with more strains are needed to confirm this hypothesis. Finally, although vancomycin treatment caused significant reductions in acetic acid and butyric acid, previously, we demonstrated that SCFAs do not inhibit growth of VRE in vitro and at the concentrations typically present in the cecum of mice (Pultz et al., 2004). Individual and mixtures of anaerobic bacteria have signature SCFA profiles (Krausse and Ullmann, 1987; Corrier et al., 1995). Concentrations of SCFA have been used as indicators of the relative health of digestive microbiota and SCFA production can be bacteriostatic and bactericidal (Goepfert and Hicks, 1969; Cherrington et al., 1992). Reductions in cecal acetic and butyric acids following vancomycin treatment are a reflection of reductions in bacteria responsible for producing these SCFAs. Therefore, it appears that development and maintenance of a healthy, digestive bacterial population are crucial for protection against VRE colonization. There is potential for replacement of affected bacteria by identifying those bacteria susceptible to vancomycin and formulation of prophylactic or therapeutic bacterial cultures (Donskey et al., 2001; Poole et al., 2001). These replacement cultures would need to contain bacteria resistant to vancomycin and, possibly, producing similar SCFA profiles.
Contrary evidence to our proposal of cecal replacement cultures to reestablish a protective microbiota may be presented by the experimentally infected poultry-derived VRE in the current study and a previous study involving older chickens supplemented with antibiotics (Robredo et al., 1999). The previous study differed from ours, in that most of the chickens were naturally colonized with vanA-positive Enterococcus hirae strains, the chickens were 4 weeks old when antibiotic treatment was initiated, no exogenous VRE was administered, and only five chickens were included in each treatment group. Vancomycin-resistant E. faecium strains emerged during treatment and when the chickens were 9 weeks old in 5 of 5 (100%), 1 of 4 (25%), and 2 of 5 (40%) chickens in the avoparcin, tylosin, and control groups, respectively. These data provide additional evidence that avoparcin, but not tylosin, treatment may be associated with emergence of vancomycin-resistant E. faecium strains in chickens. Additionally, 1 month after the cessation of antibiotic treatment, only one of the five chicks in the avoparcin group was still positive for VRE, while two each of the control and tylosin-treated chicks remained positive. The poultry-derived VRE in our artificially infected chicks continued to be isolated from all groups at 6 weeks. It is possible that poultry-derived VREs are better adapted at coexisting with other cecal bacteria than the human isolates and therefore are more resistant to conditions in the ceca hostile to lesser adapted strains.
Although VRE has not been isolated from food animals in the United States, it is plausible that these animals could become a reservoir for VRE in the future (Dunne et al., 1996; Schwalbe et al., 1999). The isolation of vanB-type vancomycin-resistant E. faecium from chicken feed in the United States (Schwalbe et al., 1999) suggests the possibility that food animals may occasionally be exposed to VRE. Our data demonstrate that young chickens may develop transient high-density colonization with human VRE isolates even in the absence of antimicrobial selective pressure. Overgrowth of these strains in the intestinal tract could result in extensive environmental contamination. Although tylosin and chlortetracycline did not promote VRE colonization, additional research is needed to evaluate the effects of other therapeutic and nontherapeutic antimicrobials that are used in food animals in the United States.
Footnotes
Disclosure Statement
No competing financial interests exist.