
Abstract
Endophytes are microorganisms that inhabit plant tissues without causing disease. Some endophytes help their hosts to combat pathogens. Here we explored the hypothesis that the plant-derived foods consumed by humans and other animals host endophytes that also antagonize foodborne pathogens or food-rotting agents. Our laboratory previously cultured a library of bacterial endophytes from different members of the maize/corn family (Zea) including wild relatives. Here, 190 of these endophytes were screened for their ability to antagonize four foodborne pathogens (Escherichia coli O157:H7, Listeria monocytogenes, Clostridium perfringens, and Salmonella enterica Newport) and a food spoiling agent (Pseudomonas fluorescens) using dual culture assays. Two Paenibacillus polymyxa endophytes (strains 3C6 and 3G11) were found to inhibit the growth of all five deleterious strains on agar. Using conserved polymerase chain reaction primers and sequencing, both beneficial endophytes were found to encode polymyxin genes, suggesting a potential antibacterial mechanism of action. Polymyxin production by both strains was confirmed using enzyme-linked immunosorbent assay. Strains 3C6 and 3G11 originated, respectively, from the seeds of the wild Central American maize species Zea diploperennis, and the wild ancestor of modern maize, Zea mays ssp parviglumis (Parviglumis). As the latter is the direct ancestor of modern maize, we discuss the role its endophyte(s) may have played in promoting crop domestication by suppressing foodborne pathogens and/or food-spoilage agents.
Introduction
E
Plant–endophyte associations sometimes form symbiotic (mutualistic) relationships by which the endophytes gain access to a protected habitat and nutrients, whereas the host gains benefits from the endophytes in terms of growth promotion, nutrient acquisition, or antagonism against plant pathogens (Rosenblueth and Martínez-Romero, 2006; Ryan et al., 2008; Johnston-Monje and Raizada, 2011). Although endophytes can be beneficial, some toxic alkaloid-producing fungal endophytes in grasses such as tall fescue and perennial ryegrass have been linked to health disorders (fescue toxicosis) in cattle and horses that consume these grasses (Guerre, 2015).
An unexplored hypothesis is whether the seeds and other foods selected by humans and other animals possess endophytes that not only help their plant hosts but also indirectly benefit human and animal health by antagonizing foodborne pathogens or food-spoilage organisms, either in the source grain/food or in foods consumed alongside (e.g., meat). Endophytes would be viable in raw foods, whereas some spores may survive cooking.
Many bacteria are causative agents of foodborne illness, of which Escherichia coli, Salmonella enterica, Listeria monocytogenes, and Clostridium perfringens are especially problematic, whereas Pseudomonas fluorescens causes spoilage to foods (Bach et al., 2002; Brynestad and Granum, 2002; Pitout et al., 2003; You et al., 2006; Allen et al., 2016; Lin et al., 2016). E. coli is a Gram-negative bacterium commonly found in the human intestine as a commensal organism, but some strains of E. coli are pathogenic such as enterohemorrhagic E. coli (EHEC). The most common strain of EHEC is E. coli O157:H7 (Callaway et al., 2009), which is a very important foodborne pathogen found in feed, water, soil, and manure (Bach et al., 2002). In humans, this pathogen causes diarrhea, hemorrhagic colitis, and hemolytic uremic syndrome.
S. enterica strains, including Salmonella enterica serotype Newport, are Gram-negative bacteria that cause salmonellosis in humans as well as in animals and is associated with gastroenteritis and bacteremia. Disease outbreaks have been associated with consumption of contaminated poultry, eggs, pork, beef, seafood, milk, cheese, chocolate, peanuts, cereal, sprouts, mangoes, and other fruits and vegetables (Van Beneden et al., 1999; Pitout et al., 2003; Sivapalasingam et al., 2003; Zhao et al., 2003).
L. monocytogenes is a Gram-positive, nonspore forming, rod-shaped bacterium that can be found in water, dairy products, meats, egg products, seafood, and vegetables (Farber and Peterkin, 1991; Embarek, 1994; Lawrence and Gilmour, 1994; Novoslavskij et al., 2015). Infection symptoms range from moderate gastrointestinal illness to severe septicaemia and meningitis, with a mortality rate of 20–30% in the case of invasive listeriosis, which makes it a major food safety concern (Carpentier and Cerf, 2011; Cossart, 2011; Allen et al., 2016). Listeriosis affects cattle, sheep, and goats with the disease associated with encephalitis, septicaemia, and uterine infections and abortion (Low and Donachie, 1997).
C. perfringens is an anaerobic, Gram-positive, rod-shaped, spore-forming bacterium that causes two types of food poisoning in humans: a mild, self-limiting type A diarrhea and the more severe type C necrotic enteritis. In animals, C. perfringens causes diarrhea, enterotoxemia, and necrotic enteritis (Brynestad and Granum, 2002; Uzal and Songer, 2008; García and Heredia, 2011; Silva and Lobato, 2015; Silva et al., 2015), whereas in poultry, it can cause avian malignant disease, gangrenous dermatitis, or avian necrotic enteritis, the most severe bacterial poultry enteric disease (Cooper and Songer, 2009).
Finally, the food spoiler P. fluorescens is a Gram-negative, rod-shaped bacterium that causes major economic losses (Andreani et al., 2015). P. fluorescens causes food spoilage by producing extracellular lipases and proteases; the bacterium is common in dairy products, fish products, and leafy greens (Miller et al., 1973; Olanya et al., 2015; Lin et al., 2016).
Our laboratory has previously isolated ∼200 bacterial endophytes from seeds, roots, and shoots of different genotypes of the maize/corn family, Zea, including wild, ancient, and modern varieties (Johnston-Monje and Raizada, 2011; Johnston-Monje et al., 2014). Among these varieties were wild Parviglumis teosinte (Zea mays ssp parviglumis), the direct, living ancestor of modern maize from Mexico (Piperno et al., 2009), and Zea diploperennis, a wild perennial Central American maize (Iltis and Doebley, 1980). Endophytes from this collection, in particular from wild and ancient maize, were recently found to host microbes with antifungal and nutrient acquisition abilities (Mousa et al., 2015; Shehata, 2016, Shehata et al., 2016a, b, c), perhaps selected by their host plants that grow without synthetic pesticides and fertilizers.
In this study, as a first step toward testing our hypothesis, we ask whether bacterial endophytes from the maize family, including those isolated from grain, can antagonize, in vitro, the mentioned bacterial foodborne pathogens as well as the food spoiler P. fluorescens.
Materials and Methods
Bacterial strains
The bacterial endophytes used in this study (Fig. 1A; Supplementary Table S1; Supplementary Data are available online at
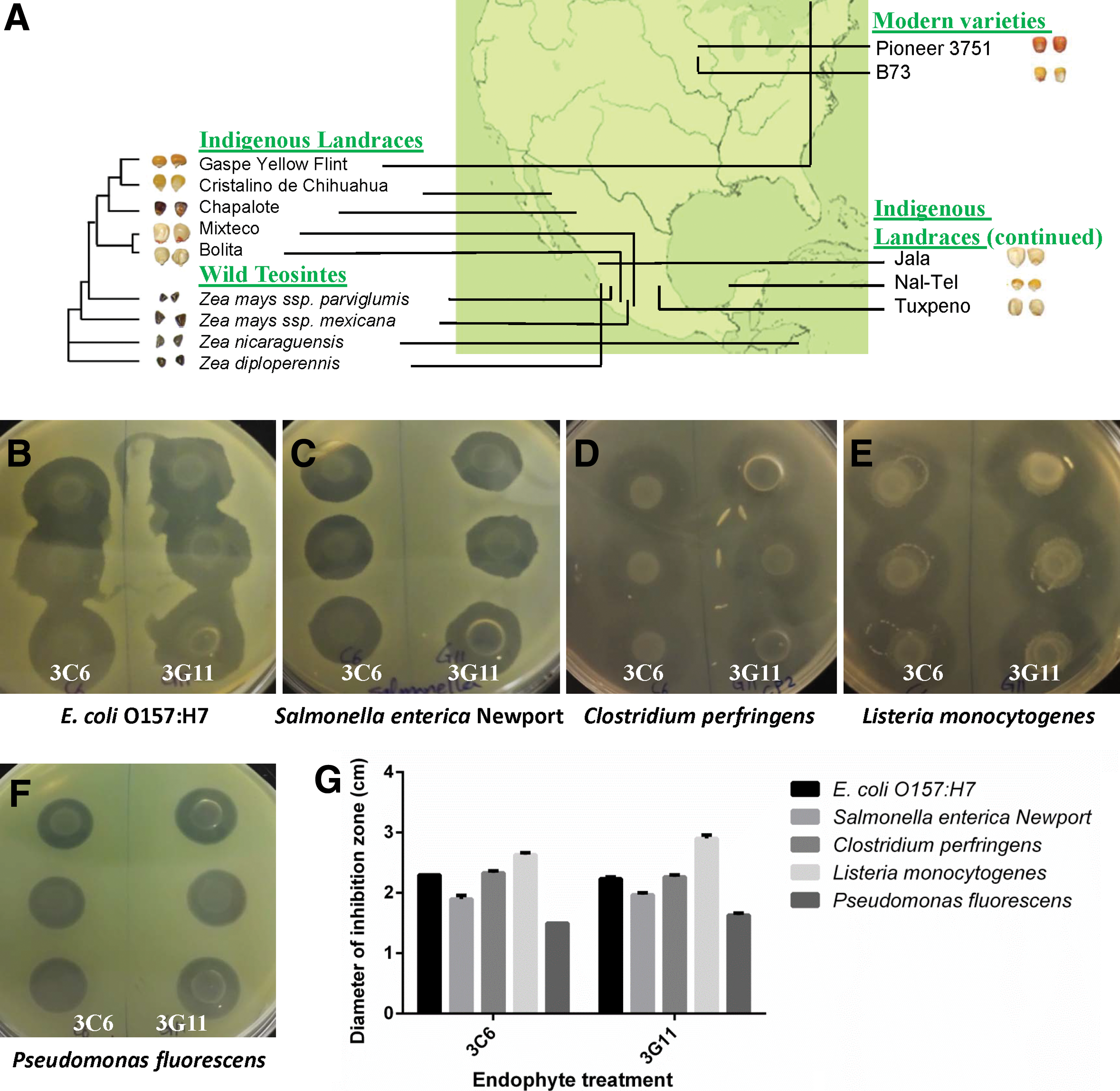
Screening of maize endophytes for their ability to suppress foodborne pathogens and a food spoiling agent.
Foodborne pathogens, Escherichia coli O157:H7 (C899), Listeria monocytogenes (LJH391), Clostridium perfringens (C1354), and Salmonella enterica Newport (C407), and food spoiler, Pseudomonas fluorescens (C942), were obtained from the Canadian Research Institute in Food Safety culture collection at the University of Guelph.
In vitro screening for antagonistic activity against foodborne pathogens
Bacterial endophytes were cultured in Luria-Bertani (LB) medium overnight at 37°C with shaking at 250 rpm. From each bacterial endophyte culture (OD595 = 1.0), 15 μL were spotted onto 10 cm Petri plates containing 15 mL of solidified LB agar, in triplicate, allowed to dry, then the plates were inverted and incubated overnight at 37°C to allow colonies to develop. In parallel, the foodborne pathogenic strains were cultured in Tryptic Soy Broth overnight at 37°C with shaking at 200 rpm. C. perfringens was cultured in Brain Heart Infusion in an anaerobic chamber at 37°C. From each pathogenic strain culture, 10 μL (OD595 = 1.0) (200 μL for C. perfringens, OD595 = 0.6) were mixed with 10 mL of melted and cooled LB agar, and then poured over LB agar on which the endophyte colonies were growing. After solidification, the plates were incubated at 37°C (in an anaerobic chamber for C. perfringens). Inhibition zone diameters were recorded (Bevilacqua et al., 2003).
Taxonomic identification of antagonistic strains based on 16S rRNA sequencing
To identify the antagonistic endophytes, full length 16S rRNA sequences were used. DNA was extracted using a Bacterial Genomic Miniprep Kit (NA2110; Sigma), and then quantified using a NanoDrop ND-1000 (Thermo Scientific, USA). Polymerase chain reactions (PCRs) were conducted using universal 16S rRNA primers (Frank et al., 2008; Ghyselinck et al., 2013) according to a previously described protocol (Shehata et al., 2016a) and then Basic Local Alignment Search Tool-Nucleotide searched against the 16S ribosomal sequence database at National Center for Biotechnology Information. A phylogenetic tree was constructed using reference 16S sequences from GenBank to confirm strain identity the phylogeny tool
Genomic fingerprinting of antagonistic strains
To test whether strains 3C6 and 3G11 are identical or distinct, BOX-PCR followed by gel electrophoresis was conducted. Each reaction consisted of 20 μL of GoTaq® Green Master Mix (M712C; Promega, Madison, WI), 2 μM BOX-A1R primer with sequence CTACGGCAAGGCGACGCTGACG, 50 ng of DNA, and double-distilled water up to 40 μL. BOX-PCR conditions were 7 min at 95°C, then 30 cycles (1 min at 95°C, 1 min at 53°C, and 8 min at 65°C), and a final extension at 65°C for 15 min (Cottyn et al., 2001). The PCR products were run on a 1% agarose gel in Tris-acetate-EDTA at 75 V for 11 h. Banding patterns were visualized and extracted using Image Lab 5.1 software.
Detection of polymyxin genes in candidate antagonistic strains
Primers for polymyxin genes pmxA, pmxB, pmxC, pmxD, and pmxE (Supplementary Table S2) were designed using Primer 3 software based on the reference sequence for Paenibacillus polymyxa strain PKB1 (GenBank: JN660148.1). PCR mixtures contained 10 μL of GoTaq Green Master Mix (M712C; Promega), 50 ng of DNA, 0.5 μL of each of the forward and reverse primers (10 μM), and double-distilled water up to 20 μL. The same reaction conditions and gel purification methods were used as in the 16S gene amplification already described. Retrieved sequences were searched using BLASTN against the Genbank nr/nt collection.
Detection of polymyxin production
Strains 3C6 and 3G11 were cultured in glucose starch calcium carbonate (GSC) medium for 3 days at 37°C with shaking at 200 rpm. The GSC medium consisted of, per liter, 20 g each of glucose, starch, and ammonium sulfate, 10 g of yeast, 2.6 g of K2HPO4, 0.1 g of FeSO4.7H2O, 0.5 g of MgSO4.7H2O, 0.25 g of sodium chloride, and 9 g of calcium carbonate (Shaheen et al., 2011). The GSC medium was previously found to be optimal for polymyxin production (Niu et al., 2013). Two milliliters from each culture were centrifuged at 10,000 rpm for 5 min. Two experiments were conducted using GSC culture supernatants. The first experiment was to evaluate the antibacterial activity of the cell-free supernatant. Culture supernatants were passed through 0.22 μm filters and from the resultant cell-free supernatants; 15 μL were then spotted on LB agar plates and assayed for activity against E. coli as already described. Polymyxin standards prepared from commercial polymyxin solution (81271; Sigma) at concentrations of 500, 250, 125, 62.5, 31.25, 15.6, and 7.8 ng/μL were spotted on agar in parallel. The second experiment was to assay for polymyxin production. In this experiment, the GSC culture supernatants were mixed with methanol at a ratio of 1:4 and allowed to incubate at 4°C for 30 min. The mixtures were then centrifuged at 10,000 rpm for 5 min. The resultant supernatants were used to assay for polymyxin production using a Colistin and Polymyxin ELISA Kit (DEIA042; Creative Diagnostics, USA). E. coli DH5-Alpha strain was used as a negative control. The test was run using 50 μL of each methanol extract, in triplicate, according to the manufacturer's recommendations. Polymyxin standards (0, 2.5, 7.5, 22.5, 67.5, and 202.5 ppb) were run in parallel, in triplicate, to construct a semilogarithmic standard curve (A% against the log concentration). The absorbance was measured at 450 nm (A450). The relative absorbance (A%) was calculated as the mean A450 of the sample or A450 of the standard x (100/A450 of the 0 ppb standard).
Statistical and graphical analyses
Prism 6 (GraphPad Software, USA) was used for graphical displays and statistical analysis. Pearson r was used to measure the linear correlation between the banding patterns of strains 3C6 and 3G11.
Results
In vitro screening for antagonistic activity against foodborne pathogens and a food spoiler
Out of 190 bacterial endophytes tested from the maize family (Fig. 1A) for antagonistic activity against four foodborne pathogens, 2 endophytes (strain 3C6 and strain 3G11) were found to show zones of growth inhibition in vitro of Escherichia coli O157:H7, Listeria monocytogenes, Clostridium perfringens, and Salmonella Newport; the same two endophytes also inhibited the food spoiling agent, P. fluorescens (Fig. 1B–G).
Taxonomic identification of candidate endophytes based on 16S rRNA
The two candidate strains were identified based on their full 16S rRNA sequences along with their positions on a phylogenetic tree constructed using reference 16S rRNA sequences. Strains 3C6 and 3G11 were found to resemble Paenibacillus polymyxa (100% and 99% similarity, respectively) (GenBank: KX015879 and KX015880) and to cluster with this species in a phylogenetic tree (Fig. 2A, B). Genomic fingerprinting showed that 3C6 and 3G11 were very closely related (Pearson correlation of banding pattern, r = 0.941), but distinct given the extra band in the genomic fingerprint of strain 3G11 (Fig. 2C–E). Strain 3C6 originated from seeds of the wild Central American maize species Zea diploperennis, whereas 3G11 originated from seeds of the direct, wild ancestor of modern maize, Zea mays ssp parviglumis (Parviglumis teosinte) (Fig. 2B).
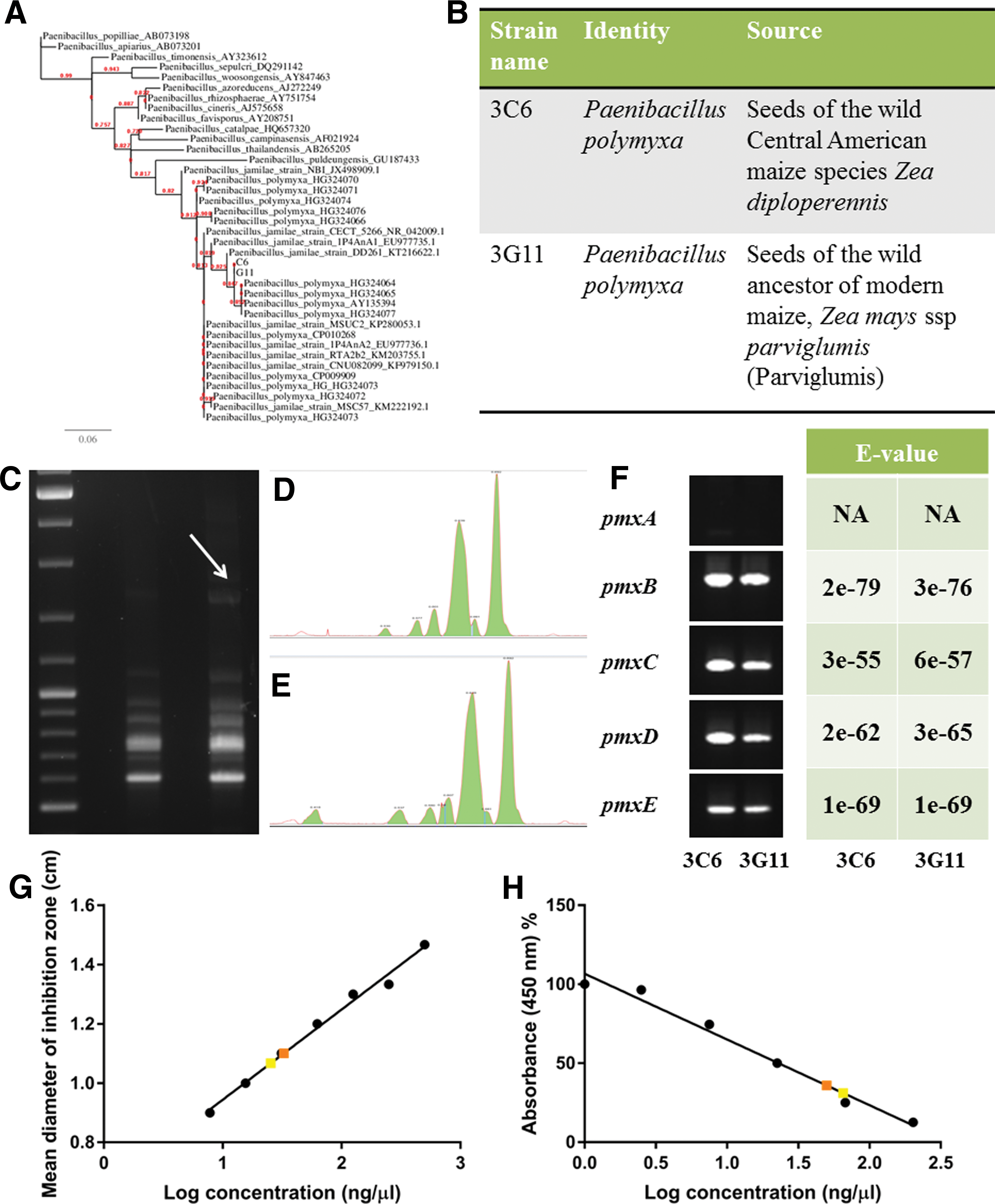
Taxonomic identification of candidate endophytes and detection of polymyxin.
Detection of polymyxin genes and polymyxin metabolite(s) in candidate antipathogen strains
As polymyxins are well-known antibiotics of previously reported P. polymyxa strains (Yahav et al., 2012), strains 3C6 and 3G11 were tested for the presence of polymyxin genes. An operon encoding polymyxin synthetase genes was previously identified from a plant growth-promoting Paenibacillus polymyxa strain E681 as having five open reading frames, pmxA, pmxB, pmxC, pmxD, and pmxE (Choi et al., 2009). Whereas pmxA, pmxB, and pmxE were found to encode polymyxin synthetases, pmxC and pmxD resembled genes that encode transport proteins. Here strains 3C6 and 3G11 were tested using PCR primers for pmxA-E followed by sequencing and BLAST analysis of amplicons of the correct size. The genomes of both strains contained sequences with high similarity to pmxB, pmxC, pmxD, and pmxE, but apparently not for pmxA (Fig. 2F; Supplementary Table S3).
To confirm the production of polymyxin from strain 3C6 and 3G11, first, cell-free supernatants were assayed for antibacterial activity. Cell-free supernatants of 3C6 and 3G11 were found to demonstrate antibacterial activity equivalent to 32.7 and 25.4 ng/μL of polymyxin, respectively (Fig. 2G). Furthermore, polymyxin was detected in methanol extracts of culture supernatants using a polymyxin ELISA Kit. Strains 3C6 and 3G11 were found to secrete 50 and 65.3 ng/μL of polymyxin, respectively, under the conditions used (Fig. 2H).
Discussion
Seeds of two wild maize genotypes, Zea diploperennis and Parviglumis teosinte, were found to possess bacterial endophytes that can suppress four foodborne bacterial pathogens and one food spoiling agent, in vitro. It is especially intriguing that Parviglumis teosinte seeds possess an endophyte (strain 3G11, predicted to be P. polymyxa) that can suppress the growth of human and livestock pathogens, including those transmitted by fecal matter. Parviglumis (also known as Balsas teosinte) is the extant, wild, direct ancestor of modern maize, selected and domesticated beginning ∼10,000 years ago in the Balsas river valley of Mexico (Matsuoka et al., 2002).
One theory of crop domestication holds that nutritious seeds were brought to temporary human settlements by animals, foraging for human food scraps, and left behind in their feces (Diamond, 1997). The animal manure was a rich nutrient source that subsequently promoted germination and growth of the seed-bearing plants. In another theory, crop domestication promoted human settlements that led to food waste, which attracted wild animals, ultimately leading to animal domestication and increased human–animal contact (Larson and Fuller, 2014). Exciting but speculative questions arise from these theories.
First, as it is known that wild animals select foods, in part, for their medicinal value (Huffman, 2003), then perhaps animals in the Balsas river valley habituated to eating Parviglumis teosinte seeds, in part, because their endophytes helped to protect the animals from dangerous bacterial pathogens that originated from other animals or increasing human populations (Harper and Armelagos, 2013). Balsas teosinte seeds are surrounded by a hard fruit case that can help them to escape damage after ingestion. Archaeological evidence has revealed the presence of animal bone fragments, including mammals, near human settlements in this region (Piperno et al., 2007; Ranere et al., 2009). Mammals are the major reservoir of E. coli strains such as strain O157:H7 (Bach et al., 2002; Delaquis et al., 2007; Franz and van Bruggen, 2008; Callaway et al., 2009), whereas listeriosis caused by L. monocytogenes affects mammals (Low and Donachie, 1997). C. perfringens is the most serious toxin-producing clostridia that affects both humans and animals (García and Heredia, 2011), and its spore-forming ability imparts longevity and persistence (Brynestad and Granum, 2002).
Alternative to the hypothesis that Parviglumis seed endophyte(s) may have helped ancient animals in the Balsas region of Mexico, perhaps the endophytes protected local humans from bacterial pathogens after direct animal to human transmission or through contaminated meat. Archaeological evidence has revealed that the ancient humans of the Balsas river valley were hunters and that animal protein was abundant (Piperno et al., 2007; Ranere et al., 2009). L. monocytogenes is a human and animal foodborne pathogen that can be transmitted to humans directly from infected animals or contaminated food (Farber and Peterkin, 1991; Embarek, 1994; Lawrence and Gilmour, 1994; Carpentier and Cerf, 2011; Cossart, 2011; Novoslavskij et al., 2015). By contrast, P. fluorescens is a food spoiler (Andreani et al., 2015), and hence Balsas teosinte grain may have had the potential to protect ancient peoples when added to foods.
Alternative to the mentioned food-related hypothesis, if humans temporarily penned small animals, leading to a buildup of dangerous manure, then perhaps P. polymyxa protected the animals that were fed Parviglumis grain. Salmonella enterica serotype Newport is known to be transmitted through infected manure used in agricultural fields, or from manure to soil to plants (You et al., 2006). In general, it is thought that many current human pathogens have their origins in agricultural domestication, in part because human populations increased, which promoted disease transmission, and in part because humans were in contact with domesticated animals, which were the sources of these pathogens (Diamond, 2002; Pearce-Duvet, 2006; Harper and Armelagos, 2013). The problem with such a theory is that there appears to be no evidence for animal domestication in this region at this time; turkeys were domesticated elsewhere in Mexico thousands of years later (Thornton et al., 2012; Larson and Fuller, 2014). However, humans at the time of maize domestication may have lived with dogs in ancient Mexico (Zeder et al., 2006), and dog-to-human transmission of pathogens has been shown, as has pathogen transmission from rodents that gather at food storage sites (Pearce-Duvet, 2006; Harper and Armelagos, 2013). The corollary theory would be that there might have been a buildup of human feces at the first settlements, which could have led to widespread outbreaks of the bacterial pathogens suppressed by P. polymyxa.
Finally, it may be that Parviglumis-associated P. polymyxa provided no direct benefits to ancient humans and animals associated with them, but rather that the endophyte, by providing resistance to a wide spectrum of bacteria including plant pathogens, protected its host and thus contributed to making Parviglumis a hardy plant that was attractive for domestication. In fact, here we uncovered the presence of an antibacterial Paenibacillus polymyxa endophyte from Zea diploperennis, a wild perennial teosinte from Central America (Iltis and Doebley, 1980). We also previously demonstrated that both wild and modern maize possess other Paenibacillus polymyxa endophytes that combat diverse fungal pathogens of crops (Mousa et al., 2015). Combined, these data are suggestive of long-term mutualism between the maize family (Zea) and its Paenibacillus polymyxa endophytes.
In terms of the antibacterial mechanism of action, our data show that strain 3G11 and strain 3C6 from Z. diploperennis, both predicted to be P. polymyxa strains, encode polymyxin genes and secrete polymyxin in vitro. Polymyxins are cationic lipopeptide antibiotics, well known for their antibacterial activity especially against Gram-negative bacteria (Choi et al., 2009; Naghmouchi et al., 2011). The mechanism of action depends on the detergent-like properties of polymyxins; they can bind membrane-associated lipopolysaccharides, resulting in disruptions to the integrity of cytoplasmic membranes and outer membranes (Naghmouchi et al., 2011; Yahav et al., 2012). Polymyxin antibiotics were used as antibiotics to treat bacterial infections, but because of their side effects, their use is currently limited to topical use on surface injuries or wounds, or used systemically in multidrug-resistant infections (Choi et al., 2009; Yahav et al., 2012). Interestingly, Polymyxin E was found to work synergistically with bacteriocins to control foodborne pathogens (L. monocytogenes and E. coli) (Naghmouchi et al., 2011). The involvement of polymyxin genes in suppressing the five harmful microbes in this study will need to be tested experimentally.
Several other antimicrobials were reported to be produced by P. polymyxa strains. Among these antimicrobials are paenibacillin, a peptide antibiotic that is active against Gram-positive bacteria (He et al., 2007), fusaricidins, a group of peptide antibiotics with activity against Gram-positive bacteria (Kajimura and Kaneda, 1997), and polyxin, a bacteriocin-like compound, with activity against Gram-positive and Gram-negative foodborne pathogens (Piuri et al., 1998).
Conclusions
Here we hypothesized that the maize family possesses endophytes that can suppress foodborne pathogens and food spoiling agents. We demonstrated, in vitro, that seeds of two wild maize relatives possess bacterial endophytes that inhibit the growth of five such harmful microbes, and we have proposed a candidate mechanism of action. One of these wild maize relatives is the direct ancestor of modern maize, raising interesting questions about the role of its endophyte(s) in promoting crop domestication by ancient Mexican farmers. More experiments in the future are needed to test the various hypotheses proposed in this article. With respect to future applications, as both endophyte strains were found to produce polymyxin, they may not be ideal for use as natural food or feed preservatives, as they may contribute to the spread of antimicrobial resistance.
Footnotes
Acknowledgments
The authors thank William Lachowsky for training and facilitating the work at the Canadian Research Institute in Food Safety at the University of Guelph. This research was supported by a grant to M.N.R. from Natural Sciences and Engineering Research Council of Canada. H.R.S. was supported by a generous scholarship from the Egyptian Government.
Disclosure Statement
No competing financial interests exist.
Authors' Contributions
H.R.S. helped to design the study, carried out experiments, and wrote the article. M.W.G. helped to design the study and supplied foodborne pathogens. M.N.R. helped to design the study and to write the article. All authors read and approved the final article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.