
Abstract
Punicalagin, one of the main active compounds in pomegranate peel, has been reported to possess many properties, including antioxidant, antimicrobial, antiviral, and immunosuppressive activities. The aim of this study was to investigate the antibacterial effect of punicalagin against Staphylococcus aureus and possible mode of action. Growth inhibition activity was examined by the agar diffusion method. Then agar dilution method was adopted to determine the minimum inhibitory concentration (MIC). The effects of punicalagin on cell membrane were assessed by measuring potassium efflux. Morphological changes of S. aureus were assessed by scanning and transmission electron microscopy. Crystal violet assay was applied to investigate antibiofilm activity of punicalagin. Punicalagin exhibited good antistaphylococcal effect with an MIC of 0.25 mg/mL. An increase of potassium efflux was observed when cells were treated with punicalagin at 2 × MIC. Punicalagin induced morphological damages to the cell membrane. Moreover, punicalagin exerted a remarkable inhibitory effect on biofilm formation of S. aureus. These findings suggest that punicalagin has antimicrobial and antibiofilm activities against S. aureus and may have potential application to control S. aureus contamination in food industry.
Introduction
S
Pomegranate has been known for hundreds of years as a healing food. Pomegranate and its peel are widely used in folk medicine for the treatment of parasite infection, diarrhea, ulcers, hemorrhage, and respiratory diseases (Tanveer et al., 2015). Punicalagin was proved to be an important bioactive phytochemical in pomegranate peel. It has been reported to possess antioxidant, antimicrobial, antiproliferative, antiviral, and immunosuppressive activities (Aqil et al., 2012; Lin et al., 2013; Danesi et al., 2014; Li et al., 2014b). Endo et al. (2010) found that punicalagin had strong antimicrobial activities against Candida albicans and Candida parapsilosis with minimum inhibitory concentrations (MICs) of 3.9 and 1.9 μg/mL, respectively. Glazer et al. (2012) reported that the growth rates of three rot fungi were negatively correlated with the levels of total polyphenolic compounds in pomegranate peel extract and particularly with punicalagin. Li et al. (2014b) revealed that punicalagin showed moderate inhibitory effect on Salmonella with an MIC of 0.25–1 mg/mL. In addition, punicalagin inhibits Salmonella virulence factors and has antiquorum-sensing potential when used at subinhibitory concentrations.
Although punicalagin has been tested against certain microorganisms for its antimicrobial potential, the information about its effects on S. aureus is limited in the literature. Therefore, the aim of this study was to investigate the antimicrobial and antibiofilm activities of punicalagin against S. aureus. The findings of this study would elucidate the antistaphylococcal and antibiofilm properties of punicalagin and provide theoretical basis for its future potential use in the food industry as a natural preservative.
Materials and Methods
Bacterial strains and culture conditions
S. aureus American Type Culture Collection (ATCC) 25923 and ATCC 29213 were obtained from ATCC. The other isolates were taken from our laboratory strain collection, including one raw milk isolate, two chicken isolates, and two dumplings isolates. All seven strains were used in MIC assay. S. aureus ATCC 25923 was used in antimicrobial mechanism study, including inhibition zone measurement, growth curves, potassium efflux, and electron microscopy. ATCC 29213 was chosen for antibiofilm assay because of its relatively high biofilm formation capacity. Strains were preserved at −80°C in trypticase soy broth (TSB; Land Bridge Technology Co. Ltd., Beijing, China) containing 25% glycerol. Before each experiment, the bacteria were streaked onto tryptic soy agar (TSA) for activation. Then one colony was inoculated into TSB and incubated at 37°C with orbital shaking to mid-log phase. Then cells were washed twice and resuspended in sterile phosphate buffered saline (PBS, pH 7.0). The bacterial suspension was adjusted using fresh PBS to OD600 = 0.5, which corresponded to ∼108 CFU/mL.
Inhibition zone measurement
S. aureus growth inhibition by punicalagin was determined using the agar diffusion method (Yi et al., 2010) by measuring the size of inhibition zones. Bacterial suspension prepared as already described was diluted 100-fold and was spread onto TSA plates (100 μL per plate). Three sterilized Oxford cups (stainless cylinders with an outside diameter of 8.0 mm, inner diameter of 6.0 mm, and height of 10.0 mm) were placed on each plate, followed by filling each cup with 200 μL of distilled water containing 0, 1, or 2 mg/mL punicalagin (≥98%, CAS 65995-63-3, Must Bio-Technology Co. Ltd., Chengdu, China). Plates were incubated at 37°C for 24 h before the clear inhibition zones were manually measured.
Agar dilution for MIC determination
The MIC of punicalagin against S. aureus was determined using the agar dilution method recommended by European Committee for Antimicrobial Susceptibility Testing (2000) with limited modifications. Punicalagin was mixed with warm (50°C) melted Mueller Hinton agar (MHA, Land Bridge Technology Co. Ltd., Beijing, China) in a 24-well plate to achieve a concentration of 4 mg/mL. The mixture was further serially twofold diluted with melt MHA. The final concentration of punicalagin was 4, 2, 1, 0.5, 0.25, and 0.125 mg/mL, respectively. MHA without punicalagin worked as control. After hardening, each well of MHA containing different concentrations of punicalagin was inoculated with a droplet (2 μL) of bacterial suspension (∼106 CFU/mL) of the tested S. aureus strain. The spots were left to dry and the plates were incubated at 37°C for 24 h. MIC was defined as the lowest concentration of punicalagin resulting in no growth of S. aureus.
Growth curves
The antimicrobial effect of punicalagin against S. aureus was assessed by examining the growth kinetics (Bharitkar et al., 2014). The log-phase bacterial suspension (100 μL, OD600 = 0.5) was diluted 100-fold into TSB containing punicalagin at concentrations of 0, 1, 2, and 4 × MIC and incubated at 37°C with orbital shaking (150 rpm). An aliquot of the culture was removed at different times of growth (0, 2, 4, 6, 8, and 24 h) and plated on TSA after adequate serial decimal dilutions. The plates were incubated at 37°C for 24 h and colonies counted manually. A kinetic growth or inactivation curve was constructed for each treatment from the viable cell counts.
Potassium efflux
Overnight culture of S. aureus in TSB was harvested by centrifugation and washed twice in saline solution (0.85% NaCl). The cells were resuspended in saline and cell density was adjusted to OD600 = 0.5. Punicalagin was added to 4 mL of the cell suspension at 2 × MIC, followed by incubation at 37°C. Controls were cell suspension without punicalagin. The supernatant was collected at 30 min intervals by centrifugal removal (8000 × g, 10 min) of cells from an aliquot of 4 mL. Extracellular potassium concentrations were measured in the supernatant using an atomic absorption spectrophotometer (Solaar, Thermo Electron Corporation) (Lou et al., 2011).
Electron microscopy
Field emission scanning electron microscopy (FESEM) and transmission electron microscopy (TEM) were employed to examine the morphological changes of S. aureus cells after punicalagin treatment.
FESEM assays were carried out as previously described (Li et al., 2014a) with minor modification. Cells were treated with punicalagin at concentrations of 0, 2, and 4 × MIC for 4 h at 37°C. The cells were harvested by centrifugation at 2500 × g for 10 min at 25°C and washed twice with PBS. Then they were resuspended in PBS containing 2.5% glutaraldehyde and kept at −4°C for 10 h for fixation. After two washes in PBS and air drying, the samples were immobilized on a support, and sprayed with Au-Pd under vacuum before examination under an FESEM (S-4800, Hitachi, Tokyo, Japan).
For TEM observation, S. aureus cells treated with punicalagin as mentioned were immobilized in agar and fixated with 2.5% glutaraldehyde for 10 h followed by 1% osmic acid for 2 h. After washing with PBS, the samples were dehydrated in acetone solution and embedded with resin. Ultrathin sections were double stained with saturated lead citrate–uranyl acetate. (Yi et al., 2010) The slices were observed with a Hitachi model HT7700 TEM.
Biofilm formation
Punicalagin was tested for its potential effect on biofilm formation of S. aureus ATCC 29213. A static biofilm formation assay was performed in 96-well polystyrene plates as previously reported (Lee et al., 2014). In brief, a twofold serial dilution was prepared containing TSB with 2% sucrose (w/v), and different concentrations of punicalagin ranging from 0 to MIC. Wells containing TSB with 2% sucrose only were used as negative control. Wells containing bacteria grown in TSB with 2% sucrose but without punicalagin treatment were used as positive control. After incubation at 37°C for 24 h, cell growth was measured through ODs at a wavelength of 630 nm using a microplate spectrophotometer (model 680; Bio-Rad). Then the planktonic cells were removed by washing three times with PBS. Cells in biofilm were fixed with methanol for 15 min and stained with 1% crystal violet. The plates were then washed with distilled water, and 250 μL of 33% acetic acid was added to each well to dissolve the residual crystal violet inside the biofilm matrix. After 5 min incubation, the absorbance (570 nm) was measured. Specific biofilm formation index (Garcia-Heredia et al., 2016) was calculated by attaching and stained bacteria (OD570) normalized with cell growth (OD630).
Statistical analysis
Statistical analyses were performed using the GraphPad software (GraphPad Software, Inc., La Jolla, CA). Data are presented as the mean values ± standard deviation. The statistical significance of the differences was calculated using the unpaired Student's t-test or one-way ANOVA. Differences were considered significant at p < 0.05.
Results
Antistaphylococcal effect of punicalagin
The antimicrobial effect of punicalagin on S. aureus was first examined by measuring the inhibition zone on agar plates. The inhibition zone diameter was 11.1 (±0.4) mm in the presence of 1 mg/mL of punicalagin, which expanded significantly to 13.0 (±0.5) mm at 2 mg/mL of punicalagin (data not shown).
Using agar dilution assay, the MIC of punicalagin against S. aureus was determined. It was shown that punicalagin exerts good inhibitory effects against seven S. aureus strains with the MIC of 0.25 mg/mL.
Growth kinetics
The growth or inactivation kinetics of S. aureus in the presence of punicalagin was examined to determine whether punicalagin inhibited the growth of S. aureus through bacteriostatic or bactericidal effects (Fig. 1). At the minimal inhibitory concentration (1 × MIC), punicalagin reduced viable S. aureus cell counts by approximately half log within 2 h of exposure. The surviving cells remained viable for several hours, and were reduced by approximately another 1 log at the end of 24 h incubation. Treatment of S. aureus by higher concentrations (2 × MIC and 4 × MIC) of punicalagin resulted in significantly more precipitous loss of viable cell counts after initial contact. At the end of treatment for 24 h, viable cell counts were reduced to less than the detection limit. These observations demonstrated that punicalagin had a strong bactericidal effect on S. aureus.

Effect of punicalagin on the growth of Staphylococcus aureus. Bacterial cells were grown in TSB with 0, 1, 2, and 4 × MIC of punicalagin at 37°C. The error bars represent standard deviation of three replicates. MIC, minimum inhibitory concentration; TSB, trypticase soy broth.
Release of potassium
Loss of cell viability is often associated with deterioration of cytoplasmic membrane integrity. To investigate the effect of punicalagin on cytoplasmic membrane, the permeability of the membranes to intracellular K+ was assessed. Figure 2 shows K+ efflux from S. aureus treated with punicalagin for different periods of time at 2 × MIC (0.5 mg/mL). In the absence of punicalagin, there was a very slow and nearly linear increase of K+ efflux over a period of 90 min. In the presence of punicalagin, the K+ efflux significantly accelerated at contact and continued to increase for the period of 90 min, resulting in approximately threefold increase in extracellular K+ concentration.
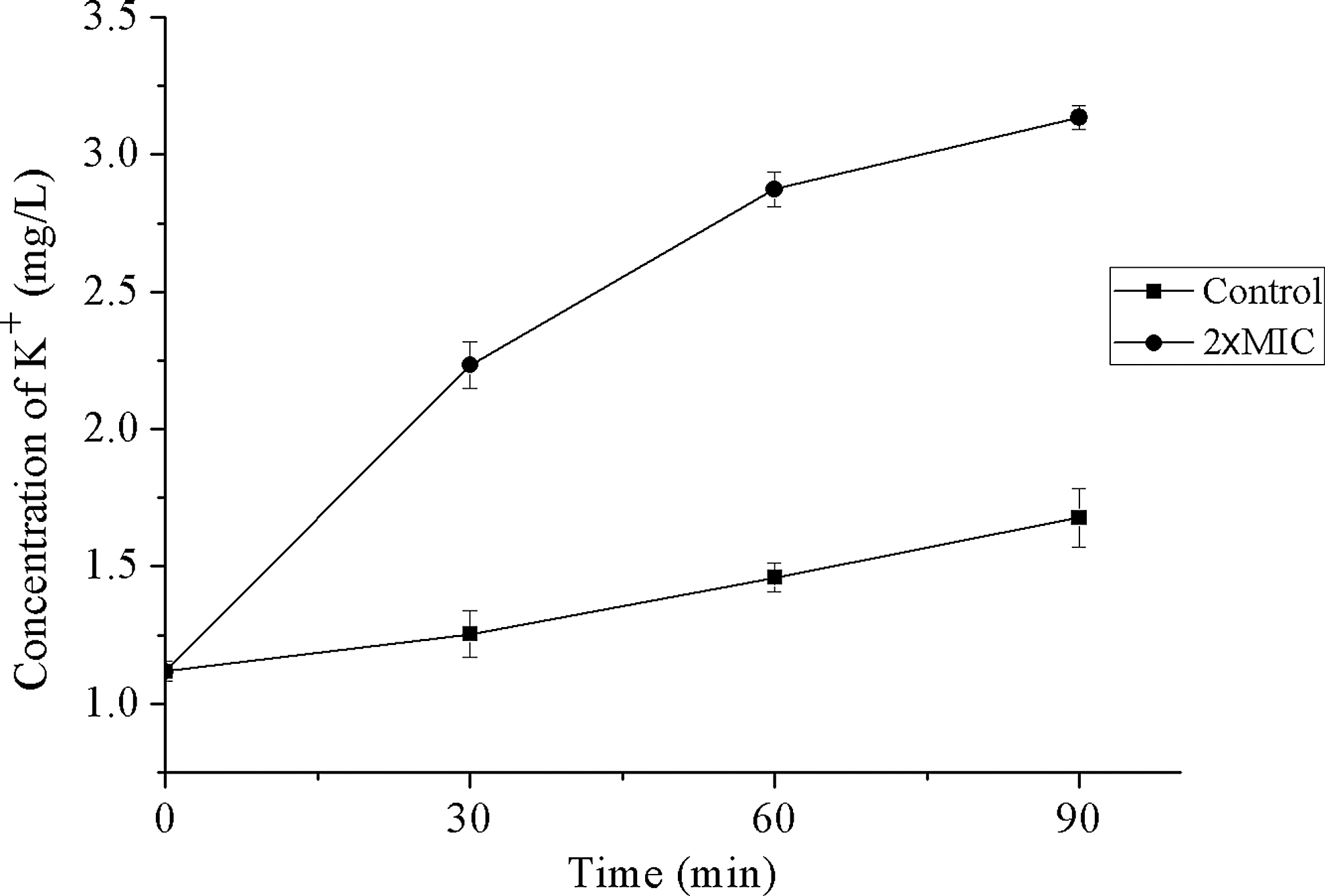
Effects of punicalagin on the extracellular concentrations of potassium ions. (■) Control. (●) Treated with 2 × MIC of punicalagin.
Microscopic observation
The effect of punicalagin on S. aureus cells was observed using electron microscopy (Fig. 3). Bacterial cells treated with punicalagin were examined by FESEM after 4 h exposure. S. aureus cells without treatment exhibited sphere contour, smooth surface, and were more uniform in size (∼0.8 μm in diameter). After treatment with punicalagin, the cells were visibly enlarged and less uniform in size and had a rough surface. Cells treated with higher concentrations of punicalagin exhibited higher degree of deformation. When examined by TEM, punicalagin-treated cells exhibited a wavy contour of cytoplasmic membrane, suggesting significant shrinking of cytoplasm. But cell lysis was not observed. We also observed a higher proportion of dividing cells, suggesting that punicalagin may interfere with cell wall synthesis or cell division.
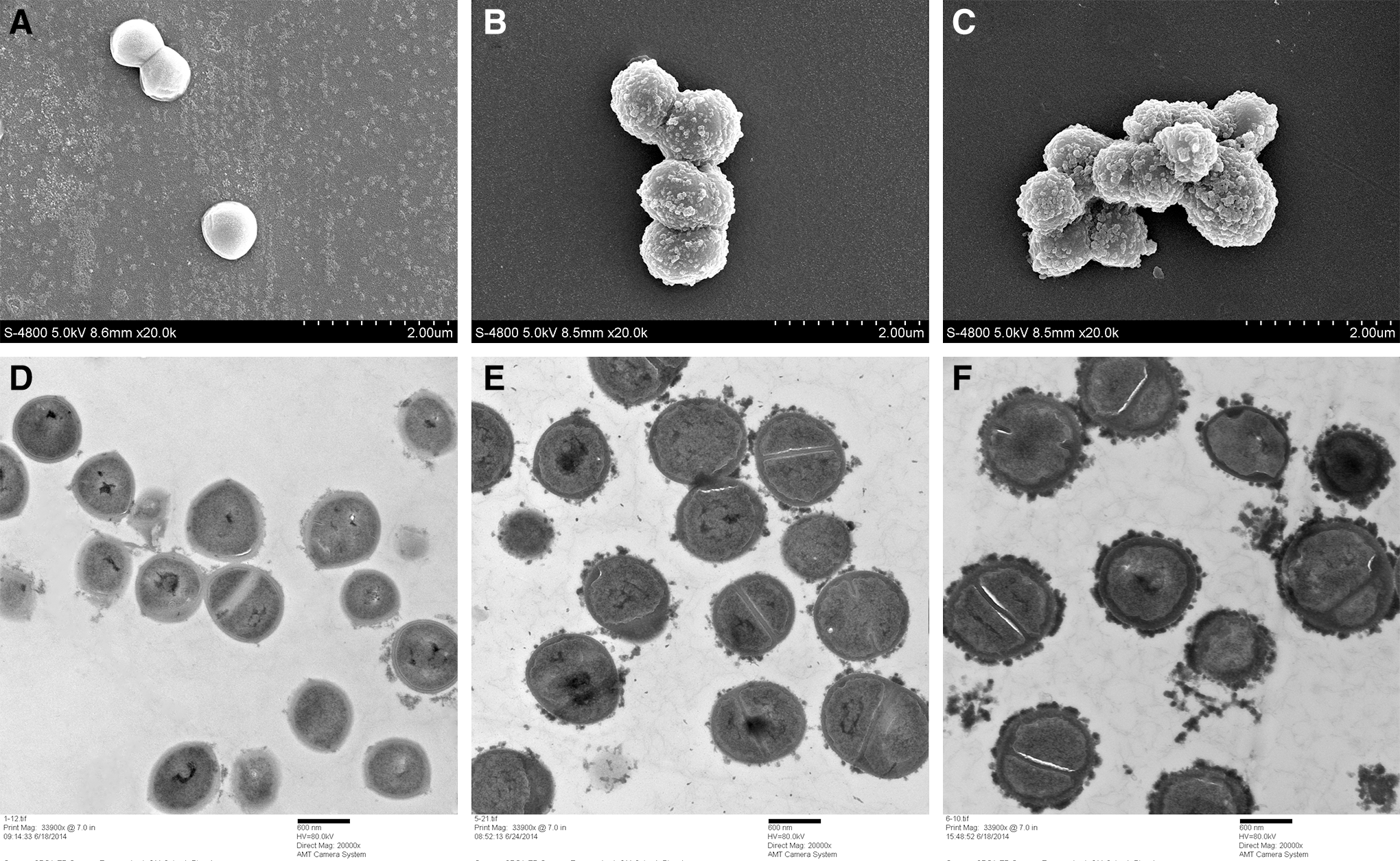
Scanning electron micrographs
Antibiofilm effect
Crystal violet staining assay was used to evaluate the effect of punicalagin on the biofilm formation of S. aureus. As presented in Figure 4, punicalagin exhibited a statistically significant inhibitory effect on the biofilm formation of S. aureus at various concentrations (p < 0.05). The biofilm formation index of the control group was close to 3.7. When treated with 1/64 × MIC of punicalagin, biofilm formation was inhibited by 47%. More than 90% of the biofilm was inhibited when the concentration of punicalagin was larger than 1/32 × MIC.

Antibiofilm activity of punicalagin against S. aureus. Biofilm formation index was determined in the presence of punicalagin at the concentrations of 1/64, 1/32, 1/16, 1/8, 1/4, 1/2, and 1 × MIC in 96-well plates. Control was bacteria grown in TSB with 2% sucrose, but without punicalagin treatment. Different letters indicated significant differences at p < 0.05.
Discussion
Although pomegranate peel exact has been widely studied for its antimicrobial activity, the antimicrobial potential of its main active component, punicalagin, has not been extensively explored. Here, we proved that punicalagin has good antimicrobial effects against S. aureus and exerts its effect through causing damage in cell membrane. In addition, punicalagin showed very strong inhibitory effect on biofilm formation of S. aureus.
The cytoplasmic membrane is the target of many antimicrobial agents. The antimicrobial activities of many compounds are generally believed to partly stem from their ability to selectively destroy the structure and function of membranes of various microorganisms (Bouhdid et al., 2009; Yao et al., 2012). It was shown that punicalagin induced immediate and massive efflux of K+ from S. aureus. The large increase of K+ release after treatment indicated that punicalagin acted on the cytoplasmic membrane by increasing its permeability or by interfering with cross-membrane proton gradient. This observation is in good agreement with other previous reports. Hada et al. (2003) showed that the total number of K+ ions lost from S. aureus cells increased with increasing concentration of tea tree oil. According to de Souza et al. (2010), the efflux of K+ ions from S. aureus cells occurred immediately after exposure to essential oils followed by a slow but steady loss.
The effect of punicalagin on S. aureus morphology was similar to that reported by Ifesan et al. (2009), who showed that Eleutherine americana extract resulted in enlargement of the S. aureus cells and increased thickness of the cell membrane. However, this result was different from the disintegrated membrane caused by many reported plant-derived compounds and other antibiotics, which can kill the microorganisms by pore formation in the cell membrane. Stojanović-Radić et al. (2012) reported a variety of morphological changes in S. aureus caused by Inula helenium L. root essential oils. Six hours after the treatment with the essential oils, cell debris can be observed after the cells collapsed. Lv et al. (2011) reported that treatment with plant essential oils resulted in cell rupture and deformations in Escherichia coli, S. aureus, and Saccharomyces cerevisiae, which were accompanied by deficiencies in cell wall structures. More severe deformation occurred in treated Bacillus subtilis cells, showing rupture and lysis of the membranes. Although cell lysis was not observed after punicalagin treatment, punicalagin led to membrane damage and increased permeability to cytoplasmic membrane in S. aureus.
Biofilms can be formed on all types of surfaces in the food industry. They are often resistant to normal sanitation procedures and are difficult to be removed. Formation of biofilms can lead to product spoilage, loss of production efficiency, and food safety problems. Many natural products have been studied for their antibiofilm activities in past several years. Chen et al. (2016) found that baicalein reduced cell attachment and biomass of S. aureus in a concentration-dependent manner. Wang et al. (2011) reported magnolol repressed biofilm formation of S. aureus and inhibited the expression of most of the genes involved in biofilm production. Punicalagin showed a significant inhibitory effect on in vitro biofilm formation of S. aureus. Although bacterial growth was not affected under the subinhibitory concentrations, punicalagin at these concentrations could probably affect expressions of many genes, including those involved in S. aureus biofilm formation.
Plant extracts have drawn wide attention for their application as natural preservatives in foods because consumers are pursuing more natural food products. Punicalagin can be extracted in a large scale from pomegranate peel, which is a byproduct in food industry. In terms of its safety, Patel et al. (2008) showed that oral intakes of up to 5 g/kg bodyweight pomegranate fruit extract (which corresponds to 1.5 g/kg punicalagins) showed no sign of acute toxicity and there was no adverse effects with 600 mg extract/kg bodyweight (180 mg punicalagins/kg bodyweight) over 90 days in rats and mice models. Punicalagin-rich pomegranate peel extract was demonstrated to be stable after thermal treatment, which may indicate the stability of punicalagin that is the main bioactive component in the extract (Qu et al., 2014). Its low toxicity, stability, together with its good antibacterial properties make punicalagin a good candidate to be developed as an alternative food preservative, which could be used in combination with other preservatives or with other food technologies. Admittedly, only a few S. aureus strains were tested in this study and more strains from different sources need to be tested to assess punicalagin's efficacy. Moreover, before its real application in the food industry, issues including interaction with various food components that may affect its antimicrobial efficacy, more cost-effective extraction from pomegranate peel, and its synergistic effect when used with other preservative technologies should be fully explored in future research.
Conclusion
In summary, punicalagin exhibited good antibacterial and antibiofilm effects against S. aureus. It affected the cell membrane, which was demonstrated by leakage of potassium along with microstructural changes. These findings indicated that punicalagin has good potential to be applied in food industry as an alternative or complementary preservative.
Footnotes
Acknowledgments
This work was supported, in part, by the Twelve-fifth Science and Technology Support Program (No.2015BAD16B08) and New Century Excellent Talent Support Plan (NCET-13-0488).
Disclosure Statement
No competing financial interests exist.