
Abstract
The aim of this study was to investigate the prevalence of fosfomycin resistance gene fosA3 and characterize plasmids harboring fosA3 among CTX-M-producing Escherichia coli from chickens in China. A total of 234 CTX-M-producing E. coli isolates collected from chickens from 2014 to 2016 were screened for the presence of plasmid-mediated fosfomycin resistance genes (fosA, fosA3, and fosC2). Clonal relatedness of fosA3-positive isolates was determined by pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing (MLST). The genetic environment of fosA3 was analyzed by polymerase chain reaction (PCR) mapping. Plasmids were studied by using conjugation experiments, PCR-based replicon typing and plasmid MLST. Sixty-four (27.4%) fosA3-positive E. coli isolates were identified in this study. The gene bla CTX-M-55 (31/64) was predominant among these strains, followed by bla CTX-M-14 (18/64) and bla CTX-M-65 (14/64). Various PFGE patterns and sequence types (STs) indicated that these isolates were clonally unrelated. Seven different genetic environments of fosA3 were identified and two new combinations (ISEcp1-bla CTX-M-65-ΔIS903D-IS26-fosA3-orf1-orf2-Δorf3-IS26 and IS26-ISEcp1-bla CTX-M-3-orf477-bla TEM-1-IS26-fosA3-orf1-orf2-Δorf3-IS26) were discovered for the first time. Conjugation experiments were successful for 47 isolates and 33 transconjugants harbored a single plasmid. Plasmids carrying fosA3 belonged to incompatibility group IncFII (17/33), IncI1 (2/33), IncHI2 (3/33), and IncB/O (1/33). F33:A-:B- plasmids carrying bla CTX-M-55, IncHI2/ST3 plasmids carrying bla CTX-M-65, and F2:A-:B-plasmids carrying bla CTX-M-55 were found in E. coli isolates from different provinces. Our results revealed a considerable prevalence of fosA3 gene among CTX-M-producing E. coli with clonal diversity from chickens in China. The transmission of different kinds of plasmids is responsible for the dissemination of fosA3 in chicken farms in China.
Introduction
T
For Escherichia coli, resistance to fosfomycin is uncommon and mainly mediated by chromosomal mutation of gene encoding fosfomycin transporters and its target enzyme (MurA) (Oteo et al., 2009; Takahata et al., 2010). However, plasmid-borne fosfomycin resistance genes, fosA3, fosC2, and fosA, which encode fosfomycin-modifying enzymes, were also discovered (Wachino et al., 2010; Ho et al., 2013a). Fosfomycin has been approved for clinical application for many years in China, but use in animals is prohibited (Hou et al., 2012; Yang et al., 2014). However, the gene fosA3 has been found spreading among fosfomycin-resistant E. coli from pets, livestock, and food animals, and even occurred in Salmonella isolates of food origin (Hou et al., 2012; Ho et al., 2013b). At present, fosA3 has mainly been found in E. coli isolated from Asian countries: Japan, mainland China, Korea, and the Hong Kong region (Lee et al., 2012; Sato et al., 2013; Chan et al., 2014). However, occasional occurrence in other countries, such as Bolivia and Germany, is also reported.
Prevalence of fosA3 among E. coli isolates from animals is believed to be related to the genetic linkage between fosA3 and other resistance determinants (bla CTX-M, rmtB, and floR), especially bla CTX-M, which made coselection possible when cephalosporins were extensively used in animal production (Hou et al., 2013). Besides, multiple transferrable plasmids coharboring fosA3 and bla CTX-M seem to play a major role in dissemination of fosA3 (Yang et al., 2014). However, currently, there is little information on prevalence of plasmid-mediated fosfomycin resistance genes among CTX-M-producing E. coli. Therefore, the aim of this study was to investigate the prevalence of plasmid-mediated fosfomycin resistance gene fosA3 among CTX-M-producing E. coli isolates from chickens collected in China, and to characterize fosA3-harboring plasmids.
Materials and Methods
Bacterial isolates
A total of 234 CTX-M-producing E. coli isolates were recovered from chicken feces and diseased or dead chickens from seven provinces and municipalities in China (Anhui, Hubei, Henan, Sichuan, Tianjin, Hebei, and Ningxia) between September 2014 and June 2016. All isolates were identified by using BD Phoenix-100 Automated Microbiology System (BD Diagnostic Systems, Sparks) with identification panels NMIC/ID-4 (catalog no. 448505; 45 reactions/51 wells). Fosfomycin-resistant E. coli isolates were selected on Mueller–Hinton (MH) agar plates supplemented with 64 μg/mL fosfomycin (Macklin Biochemical Co., Ltd., Shanghai, China) and 25 μg/mL glucose-6-phosphate (Solarbio Science & Technology Co., Ltd., Beijing, China), and subjected to further study.
Antimicrobial susceptibility testing
Antimicrobial susceptibility test was performed by the disk diffusion method on MH agar plates and evaluated according to the guidelines in Clinical and Laboratory Standards Institute (CLSI, 2016), except for florfenicol (CLSI, 2013). The zone diameter interpretive standard for each drug is listed in Supplementary Table S1 (Supplementary Data are available online at
Molecular detection
All selected fosfomycin-resistant or fosfomycin-insensitive E. coli isolates were subjected to screening for the plasmid-mediated fosfomycin resistance genes fosA3, fosC2, and fosA by polymerase chain reaction (PCR) amplification using primers described previously (Hou et al., 2012). Positive products were confirmed by sequencing. The regions surrounding fosA3 were investigated by PCR mapping and sequencing using primers listed in Supplementary Table S2. Florfenicol resistance gene floR and 16S rRNA methyltransferase gene rmtB were also detected as described before (Maynard et al., 2003; Doi and Arakawa, 2007). The genotype of bla
CTX-M was determined by sequencing products amplified using specific primers (Zhang et al., 2014) and analyzing online using BLAST (
Strain typing and phylogenetic group analysis
Pulsed-field gel electrophoresis (PFGE) of XbaI (TaKaRa, Dalian, Liaoning, China)-digested genomic DNA was performed using the CHEF-MAPPER System (Bio-Rad Laboratories, CA) as described previously (Yu et al., 2015). Salmonella serovar Braenderup H9812 was used as a standard size marker. PFGE patterns were analyzed with BioNumerics software (Applied Maths) using the unweighted pair group method with arithmetic averages based on Pearson similarity coefficient. Isolates with Pearson similarity ≥85% were regarded as clonally related. Multilocus sequence typing (MLST) was performed according to the protocols published before (Wirth et al., 2006) and sequence types (STs) were obtained by submitting amplicon sequence to
Conjugation experiments
The transferability of fosA3 gene was studied by conjugation experiments using rifampin-resistant E. coli C600 as the recipient strain. Transconjugants were selected on MH agar plates supplemented with fosfomycin (25 μg/mL), rifampin (400 μg/mL), and glucose-6-phosphate (25 μg/mL), and finally confirmed by ERIC-PCR (Mohapatra et al., 2007). The antimicrobial susceptibility test was performed as described above for all transconjugants. The minimum inhibitory concentrations of cefotaxime (CTX) and fosfomycin (FOS) were determined by the agar dilution method for donors and transconjugants according to CLSI guidelines. E. coli ATCC 25922 was used as the quality control strain. The presence of fosA3, bla CTX–M, rmtB, and floR in transconjugants was also detected by PCR as mentioned above.
Plasmid analysis
The size and number of plasmids in all transconjugants were determined by S1 nuclease PFGE (S1 PFGE) and transconjugants with a single plasmid were used for plasmid analysis. Whole genomic DNA was digested with S1 nuclease (Takara Biotechnology, Dalian, China) and subjected to PFGE as described previously (Barton and Harding, 1995). Lambda PFG Ladder (New England Biolabs, Beverly, MA) was used as the size standard. PCR-based replicon typing was performed to classify different incompatibility groups as described by Carattoli et al. (2005). In addition, replicon sequence typing for IncF plasmids, plasmid multilocus sequence typing (pMLST) for IncI1 plasmids, and double locus sequence typing (pDLST) for IncHI2 plasmids were carried out to distinguish plasmid subtypes as described previously (Garcia-Fernandez et al., 2008; Garcia-Fernandez and Carattoli, 2010; Villa et al., 2010). Allele numbers and STs were obtained through submitting the amplicon sequence to the plasmid MLST database (
Nucleotide sequence accession numbers
The nucleotide sequences of new genetic environments surrounding fosA3 have been deposited in the GenBank database under the accession numbers KX495605 and KX578603.
Results
Detection of antimicrobial resistance genes
Of the 234 CTX-M-producing E. coli, 64 (27.4%) isolates showed resistance or insensitivity to fosfomycin. All of them were positive for fosA3, but negative for fosA and fosC2. Gene detection and sequencing confirmed that genotype of gene bla CTX-M included bla CTX-M-55 (n = 31), bla CTX-M-14 (n = 18), bla CTX-M-65 (n = 14), bla CTX-M-15 (n = 2), bla CTX-M-3 (n = 1), bla CTX-M-64 (n = 1), and bla CTX-M-123 (n = 1), and 4 isolates carried two different bla CTX–M genes (bla CTX-M-55: bla CTX-M-14; bla CTX-M-55: bla CTX-M-65 (n = 2); bla CTX-M-15: bla CTX-M-14). In addition, 9 and 45 isolates harbored rmtB and floR genes, respectively (Fig. 1).
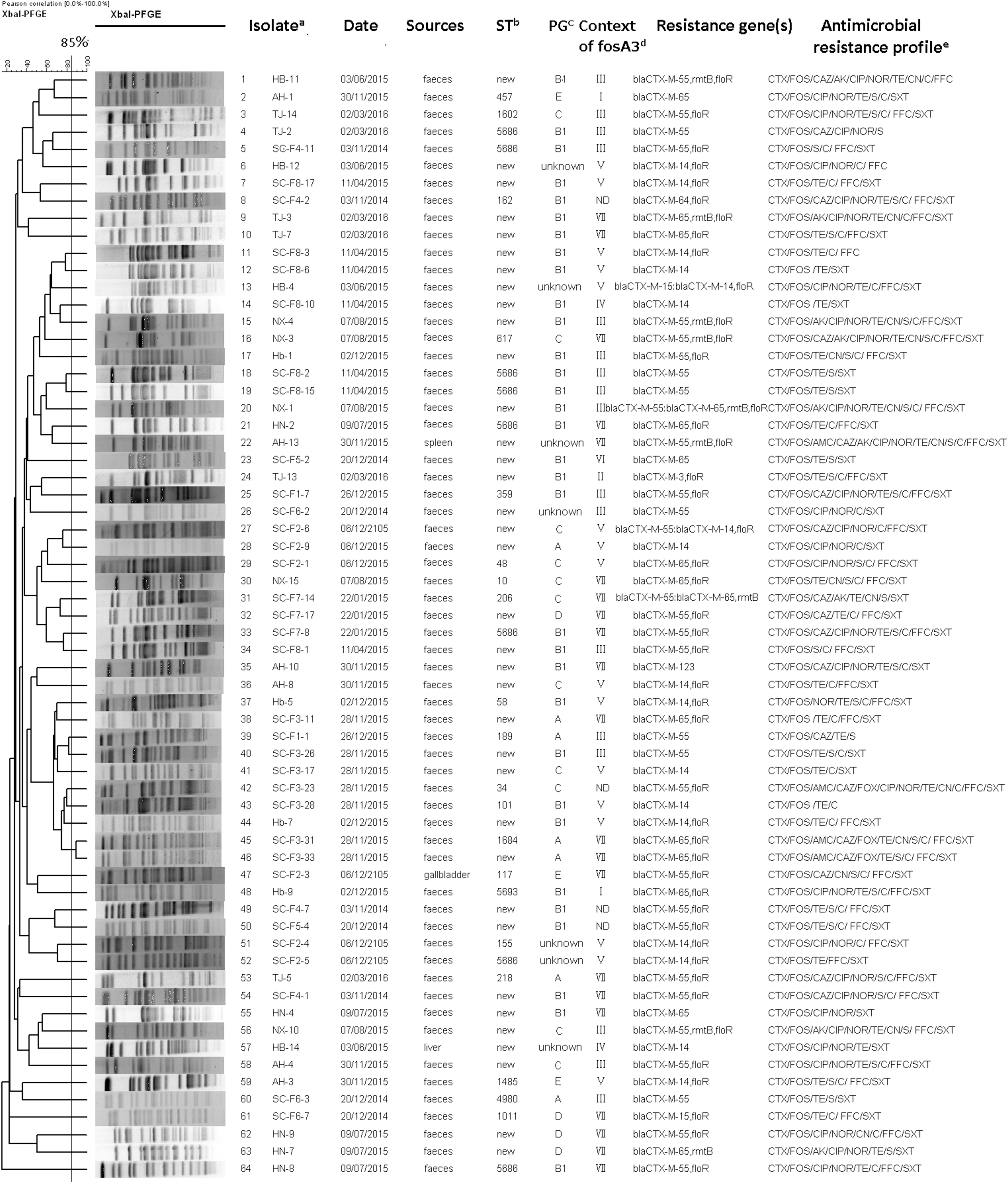
XbaI-PFGE dendrogram and detailed information about fosA3-positive E. coli isolates. aAbbreviations for provinces and municipality: HB, Hebei; Hb, Hubei; AH, Anhui; TJ, Tianjin; SC, Sichuan; NX, Ningxia; HN, Henan; F1–F8, farm1–farm8; bnew ST numbers were not obtained and only allele numbers were provided in Supplementary Table S3; cPG, phylogenetic group; dND, not determined; I–VII represent genetic environments (Type I–Type VII) of fosA3 in Figure 2; eAbbreviations for antimicrobial drugs: AMC, amoxicillin/clavulanic acid; CTX, cefotaxime; CAZ, ceftazidime; CIP, ciprofloxacin; NOR, norfloxacin; FOX, cefoxitin; AK, amikacin; FOS, fosfomycin; CN, gentamicin; S, streptomycin; C, chloramphenicol; TE, tetracycline; SXT, sulphamethoxazole/trimethoprim; FFC, florfenicol. PFGE, pulsed-field gel electrophoresis; ST, sequence type.
Genetic environments of fosA3
The regions adjacent to fosA3 were determined by PCR mapping. Seven different genetic environments were identified and arbitrarily designated as type I to VII (Fig. 2). In most isolates, fosA3 were located between two IS26 elements oriented in the opposite direction. The downstream region of fosA3 was an IS26 element in 60 isolates and it could not be identified for other five isolates. The length of the spacer regions between the 3′ end of fosA3 and IS26 was variable (536, 1222, and 1758 bp). In five structures (Type I, II, III, VI, VII), an IS26 element was located upstream of fosA3 (n = 46). The length of the upstream IS26/fosA3 intergenic space was 322 bp. The other two structures (Type IV, V) were from isolates harboring gene bla CTX-M-14. In these isolates, the upstream IS26 elements were replaced by two similar structures, ISEcp1-bla CTX-M-14-IS903-465bp-fosA3 and IS26-ISEcp1-bla CTX-M-14-ΔIS903-258bp-fosA3. Two new genetic environments surrounding fosA3 were identified in this study (Type I and II). Structure ISEcp1-bla CTX-M-65-ΔIS903D-IS26-fosA3-orf1-orf2-Δorf3-IS26 (accession no. KX495605) from isolate Hb-9 was similar with that on plasmid pV408 (accession no. LN735560.1) from E. coli in Bolivia, except that a putative iron outer membrane receptor gene (iroN) and its flanking regions were deleted from this structure. Genetic structure IS26-ISEcp1-bla CTX-M-3-orf477-bla TEM-1-IS26-fosA3-orf1-orf2-Δorf3-IS26 (accession no. KX578603) from isolate TJ-13 was identical with that on plasmid pHNAH9 from chicken E. coli in China (accession no. KJ668699.1), except for Δorf3 region, which only shared 90% identity with the corresponding region.
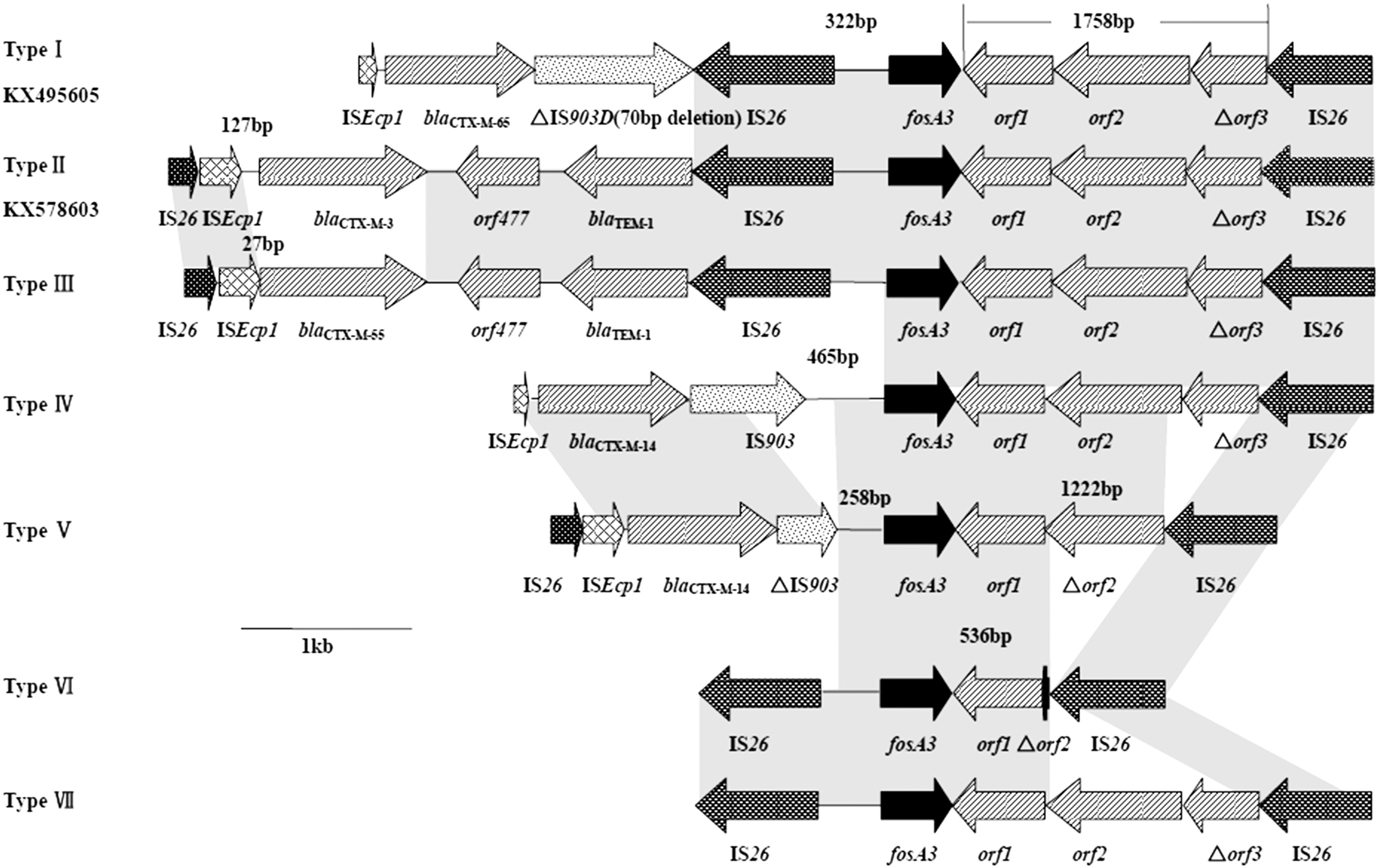
Schematic representation of the genetic environments of fosA3. Gray areas indicate regions with 100% nucleotide sequence identity; the arrows represent the position and orientation of the depicted genes; Black bar in type VI structure denotes truncated Δorf2 (14 bp).
Characterization of fosA3-positive isolates
Phylogenetic group analysis showed that 64 isolates were distributed into six groups (A, B1, C, D, E, and unknown; Fig. 1). Group B1 was predominant (32, 50.0%), followed by C (11, 17.2%), A (7, 11.0%), D (4, 6.25%), and E (3, 4.7%). Seven isolates could not be assigned to any of the groups.
Dendrogram of XbaI-PFGE patterns reflected clonal diversity of the 64 fosA3-positive isolates (Fig. 1). A total of 64 different PFGE patterns were obtained and there was no predominant cluster. Only one cluster with a similarity index ≥85% containing two isolates was identified and they were recovered from the same farm in Sichuan Province. Clonal unrelatedness was not only observed between isolates from different provinces but also between isolates from the same province, even from the same farm (SC-F8-1, SC-F8-2, and SC-F8-17).
MLST revealed 30 new STs for 36 isolates and 21 known STs for the remaining 28 isolates (Fig. 1). The allele profiles of new STs were provided in Supplementary Table S3. The known STs in this study fell into seven clonal complexes (CCs), in which CC10 was predominant, and nine singletons. Each ST had only one isolate, except eight ST5686 isolates.
Conjugation experiments and plasmid analysis
The gene fosA3 was successfully transferred from 47 donors to the recipients by conjugation experiments. S1 PFGE demonstrated that 33 transconjugants harbored single plasmid and they were used for plasmid analysis (Table 1). The plasmids present in transconjugants were assigned to the following incompatibility groups: IncFII (n = 17), IncHI2 (n = 3), IncI1 (n = 2), and IncB/O (n = 1). Interestingly, multiple replicons were detected in 10 transconjugants, which might be fused plasmids. For five plasmids with multiple replicons (pAH-3, pHB-4, pHN-4, pSC-F7-17, and pSC-F6-7), the plasmid sizes were much larger than that in donor strains, indicating that the fusion happened in conjugation process. By replicon sequencing, IncF II plasmids were divided into F33:A-:B- (n = 8), F18:A-:B- (n = 1), F18:A-:B1 (n = 4), F2:A-:B- (n = 3), and F24:A-:B1 (n = 1) subtype. All F33:A-:B- plasmids coharbored gene fosA3 and bla CTX-M-55. IncI1 plasmids were assigned to two STs (ST71 and ST136) by pMLST. All IncHI2 plasmids belonged to ST3 by pDLST. At least one antimicrobial resistance was cotransferred in all transconjugants with a single plasmid. Resistance to CTX was always observed with the transfer of fosA3, except plasmid pSC-F4-2. PCR results also confirmed that these plasmids coharbored gene bla CTX-M, except this one. Moreover, cotransfer of gene rmtB and/or floR with fosA3 was observed in 17 plasmids (51.5%).
Abbreviations for provinces and municipality: HB, Hebei; Hb, Hubei; AH, Anhui; TJ, Tianjin; SC, Sichuan; NX, Ningxia; HN, Henan; F1–F8, farm1–farm8.
Abbreviations for antimicrobial drugs: CTX, cefotaxime; AK, amikacin; FOS, fosfomycin; CN, gentamicin; S, streptomycin; C, chloramphenicol; TE, tetracycline; SXT, sulphamethoxazole/trimethoprim; FFC, florfenicol.
MIC, minimum inhibitory concentration.
Discussion
This study investigated the prevalence of plasmid-mediated fosfomycin resistance genes among CTX-M-producing E. coli from chickens in China, and several representative plasmids carrying fosA3 were characterized. To our knowledge, this was the first report to study the prevalence of fosA3 among CTX-M producers of chicken origin. Gene fosA3 has been detected in E. coli isolates recovered from clinical samples, pets, pigs, wild animals, and other food animals and its prevalence rate was at a rather low level (1.1–9.0%) (Hou et al., 2012; Ho et al., 2013a, 2013b). However, we revealed a high prevalence of fosA3 (27.4%) in CTX-M producers of chicken origin, which was much higher than that reported in isolates from clinical samples in China (0.32%) and Korea (3.0%) (Lee et al., 2012; Ho et al., 2013a) and in isolates from chickens in China (8.8%) reported before (Yang et al., 2014). It could be explained by the fact that all strains in our study were CTX-M producers and fosA3-carrying strains always coharbored bla CTX-M in previous studies (Hou et al., 2012; Sun et al., 2012; He et al., 2013; Ho et al., 2013b). Genetic environment detection confirmed that diverse bla CTX-M genes (bla CTX-M-3, bla CTX-M-55, bla CTX-M-14, and bla CTX-M-65) had genetic linkage with gene fosA3 through mobile elements (IS26, IS903; Fig. 2). A variety of structures linking fosA3 and bla CTX-M have been reported (Lee et al., 2012; Sato et al., 2013) and we discovered two new structures surrounding fosA3 (accession nos. KX495605 and KX578603).
Furthermore, fosA3 is always found to be associated with rmtB and floR (Pan et al., 2014, 2016; Yang et al., 2016). In this study, rmtB was coharbored on one plasmid with fosA3 in four isolates, while floR was coharbored with fosA3 in 15 isolates. The association with other resistance determinants is likely to facilitate the dissemination and maintenance of fosA3 by coselection, since cephalosporins, aminoglycosides, and florfenicol have been frequently used as treatment or feed additives in animal husbandry in China (Lu et al., 2011; Deng et al., 2013; Yang et al., 2015). That may be the reason why increasing fosfomycin resistance and prevalence of fosA3 were observed even though fosfomycin was forbidden in animal production in China (Ho et al., 2013b; Yang et al., 2014). However, it is still confusing why fosA3 is more prevalent in isolates of animal origin than in isolates of human clinical origin, where fosfomycin is often prescribed in the treatment of urinary tract infections (Raz, 2012; Tseng et al., 2015).
In this study, fosA3-positive E. coli isolates were collected from seven provinces and municipalities in China, together with previous report that fosA3 has been found in E. coli isolates from Guangdong, Guangxi, Shandong province (Yang et al., 2014), suggesting that fosA3 has spread in chicken farms all over our country. Based on PFGE information, we disclosed a high clonal diversity of fosA3-positive isolates. Cluster analysis showed that almost all isolates were clonally unrelated. It could be inferred that the spread of gene fosA3 among E. coli isolates of chicken origin in China was not attributed to clonal expansion. This conclusion was also supported by MLST results that 21 known STs plus 30 new allele combinations were identified. A study from Hong Kong demonstrated clonal diversity of E. coli isolates carrying fosA3 from livstock and other animals, and six CCs were found based on MLST analysis (Chan et al., 2014). By contrast, we showed a higher diversity with seven CCs even though the unregistered STs were not included. This finding highlights the possibility that fosA3 is likely to spread by clonal expansion in the future, although, which has not been observed so far.
Previous studies found that plasmids carrying fosA3 were distributed into incompatibility group IncF II, IncN, IncI1, IncB/O, IncHI2, and untypeable (Ho et al., 2013a, 2013b; Hou et al., 2013; Yang et al., 2014). These plasmids are also significant platforms for horizontal transfer of gene bla CTX-M (Zhao and Hu, 2013). Conjugative plasmids of multiple incompatibility groups were found in our study, including IncF II, IncHI2, IncI1, and IncB/O. IncB/O plasmid carried fosA3 with no bla CTX-M content in a previous study (Ho et al., 2013b). However, in our study, bla CTX-M-65 was located on this plasmid. Consistent with this, F18:A-:B1 plasmid has been found to be associated with bla CTX-M in E. coli, but there was no report that fosA3 was located on this plasmid before (Matsumura et al., 2013; Ben Sallem et al., 2014). We identified fosA3 on this plasmid for the first time. This suggested that gene fosA3 might integrate with bla CTX-M on one plasmid in the process of dissemination, which was a good explanation to the origin of fosA3-carrying plasmids. IS26 element might play a major role in this activity, for IS26 element was found upstream and downstream of both gene fosA3 and bla CTX-M. Many studies have clarified that IS26 plays a key role in disseminating antibiotic resistance genes in Gram-negative bacteria (Harmer et al., 2014; He et al., 2015). This phenomenon is worrisome when we take account of the increasing prevalence and diversity of gene bla CTX-M in animals in China (Xu et al., 2015), which improves possibility and complexity of fosfomycin resistance transfer.
More importantly, F33:A-:B- plasmids carrying bla CTX-M-55, IncHI2/ST3 plasmids carrying bla CTX-M-65, and F2:A-:B- plasmids carrying bla CTX-M-55 were found in E. coli isolates from different provinces. In E. coli isolates from pets and food animals in China, F33:A-:B- plasmids often coharbor fosA3 and bla CTX-M-65 (Hou et al., 2012, 2013). All F33:A-:B- plasmids encoded bla CTX-M-55. Similar plasmids were also found in E. coli isolates of chicken origin in Guangdong and Shandong provinces (Yang et al., 2014), indicating chicken farms were hot spots for dissemination of F33:A-:B- plasmids carrying bla CTX-M-55 in China. The combination of fosA3 and bla CTX-M-55 on F2:A-:B- plasmid had never been reported before; we identified this plasmid in two provinces (Sichuan and Tianjin) for the first time. Actually, it is bla CTX-M-3 that often coexists with fosA3 on this plasmid in E. coli isolates from pets, chickens, pigs, and human samples (Hou et al., 2012; Ho et al., 2013b; Sato et al., 2013). IncHI2/ST3 plasmid carrying bla CTX-M-65 and fosA3 was originally identified in E. coli isolates of chicken origin in China (Guangdong and Shandong) (Yang et al., 2014). We identified this plasmid again in two different provinces (Tianjin and Sichuan), implying that this plasmid was in certain dissemination in chicken farms in China. Therefore, these plasmids seemed to be capable of spreading very efficiently and indicated that the prevalence of fosA3 was ascribed to the dissemination of plasmids carrying fosA3. This result reinforced the perspective that the transfer of several important plasmids was the primary reason for dissemination of fosA3 in chicken farms in China (Yang et al., 2014).
Conclusions
Our study revealed a considerable prevalence of fosA3 among CTX-M-producing E. coli with clonal diversity from chickens in China. The dissemination of fosA3 might possibly be explained by the horizontal transfer of various fosA3-carrying plasmids, which comes from integration events mediated by IS26 element. These plasmids often coharbored other resistance genes, such as bla CTX-M, rmtB, and floR. Even though fosfomycin was not used in chicken farms in China, it was possible that fosA3 was coselected when other antimicrobial agents (cephalosporins and florfenicol) were used. It may be not enough to control the generation of fosfomycin resistance and spread of fosA3 just by forbidding the usage of fosfomycin in animal husbandry without a general more rational policy on antibiotics.
Footnotes
Acknowledgments
This work was supported by National Key Basic Research Program of China (No. 2013CB127200). We thank all colleagues who made contributions to this work.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.