
Abstract
Salmonella enterica subsp. enterica can colonize tomato fruit as it interacts with fruit surface compounds. The exometabolome of tomato fruit contains a mixture of compounds, including fatty acids, which could affect Salmonella fitness. Fatty acids detected in fruit exudates were investigated for Salmonella inhibition. Pelargonic, lauric, myristic, palmitic, margaric, stearic, and oleic acids were suspended in water dissolved in dimethyl sulfoxide (DMSO) or emulsified in water and quillaja saponin to assess how bioavailability impacted Salmonella growth. The minimum inhibitory concentrations of fatty acids were determined using a resazurin assay. Quillaja saponin emulsion and DMSO solution of pelargonic acid were inhibitory to Salmonella at 31.25 mM. Lauric and myristic acid emulsions inhibited growth at 1 M concentrations in quillaja emulsions and 62.5 mM in DMSO. Lauric and myristic acids significantly affected growth of Salmonella Newport, Javiana, and Typhimurium (p ≤ 0.05). Growth curve analysis using the Baranyi model revealed reduced maxima populations for all treatments (p ≤ 0.001) and shorter lag phase durations for Salmonella Newport with lauric acid (p < 0.01) and Salmonella Javiana with lauric (p < 0.001) and myristic (p < 0.001) acids. Salmonella Newport and Javiana exhibited an accelerated growth rate with lauric acid (p < 0.001) as a result of early stationary phase transition (shorter log phase). In myristic acid-amended media, Salmonella Javiana also displayed a faster growth rate (p < 0.001). Pelargonic acid (31.25 mM) treatment of Salmonella cells resulted in a drop in culturable cells to below detection in an hour. Microscopic analysis with Cyto-dye and propidium iodide of bacterial cells treated with pelargonic acid indicated a mixture of live and dead cells, with cell lysis of some cells. A subset of cells exhibited elongation—possibly indicating filament formation, a known antibiotic stress response. The results suggest that fatty acids present in tomato fruit surface exudates may exert a restrictive effect on Salmonella growth on fruit.
Introduction
T
Following contamination of tomato fruit with S. enterica, phenotypic expression, survival, and persistence of this pathogen could be influenced by metabolites present on the tomato surface. The exometabolites on fruit surfaces include sugars, amino acids, organic acids, a variety of fatty acids (Han and Micallef, 2016), and breakdown products of fatty or amino acids (Jadhav et al., 1972). Sugars and amino acids can serve as nutrients for bacterial growth resulting in pathogen population increases, while fatty acids might influence colonization and persistence of Salmonella enterica serotypes during fruit maturation (Noel et al., 2010). Fruit exometabolite profiles were shown to vary among tomato cultivars, and this variation was correlated with differences in S. enterica growth among cultivars, with a number of fatty acids being associated with depressed growth (Han and Micallef, 2014, 2016).
Fatty acids exuded to the tomato fruit surface vary in their bioavailability and concentrations. Fatty acids may play a role in plant protection against human pathogen colonization. Linoleic acid in green tomatoes caused an upregulation of the fadH promoter, required for the metabolism of unsaturated fatty acids in S. enterica (Noel et al., 2010). Medium chain fatty acids reduced growth and pathogenicity of S. enterica through influencing the expression of pathogenicity genes (Van Immerseel et al., 2004). Linoleic, oleic, palmitic, and stearic acids resulted in inhibition of the quorum sensing molecule AI-2 (Auto Inducer 2) required for cell to community and cell to cell communication (Widmer et al., 2007). Fatty acids can have bacteriostatic activity resulting in their use as food preservatives. Their mode of action against bacteria might involve interaction with the bacterial cell membrane and their acidic pH could disrupt proton gradients across bacterial cell membranes (Ricke, 2003).
Fatty acid profiles in tomato exometabolites were not only cultivar specific but also changed over the course of plant development and differed among different plant organs (Han and Micallef, 2016). Since we have previously shown that the presence of fatty acids correlated with reduced growth of S. enterica in tomato surface compounds (Han and Micallef, 2016), a better understanding of the impact of fatty acids on the growth of S. enterica could provide data that may be used in tomato breeding programs or in cultivar selection. In this study, we investigated the susceptibility of three environmental isolates of S. enterica recovered from tomato farms (Micallef et al., 2012) against seven fatty acids previously identified in tomato fruit exudates.
Materials and Methods
Bacterial strains
The three Salmonella enterica subsp. enterica serotypes used in this study were Salmonella Newport, Javiana, and Typhimurium. The isolates used were obtained from Mid-Atlantic tomato farms—Salmonella Newport and Javiana isolated from irrigation pond water and Salmonella Typhimurium isolated from soil (Micallef et al., 2012). Frozen isolates maintained at −80°C in Brucella Broth (BD, Sparks, MD) containing 15% glycerol were revived in tryptic soy broth (TSB; BD) by incubating for 18 h at 37°C. Turbid cultures were streaked on Xylose Lysine Tergitol 4 Agar (XLT4; BD), and typical colonies were used to create overnight cultures for further experiments.
Fatty acids and stock preparation
The fatty acids tested were the saturated pelargonic (C9:0) (TCI, Tokyo, Japan), lauric (C12:0) (TCI), myristic (C14:0) (TCI), palmitic (C16:0) (Sigma, St. Louis, MO), margaric (C17:0) (Acros Organics, New Jersey), and stearic (C18:0) (Alfa Aesar, Ward Hill, MA) acids and the unsaturated oleic acid (18:1) (TCI). The fatty acids were tested in their free form (nonesterified), although in tomato fruit they may also be present in their esterified form. Two molar concentrations of stock solutions of fatty acids were prepared in sterilized deionized water (SDW) autoclaved at 121°C for 15 min, or SDW containing a final concentration of 0.0001% (w/v) filter-sterilized quillaja tree bark saponin (Sigma). The fatty acids were suspended in water by placing the samples in a 75°C water bath for 15 min until the fatty acids melted. The melted fatty acids were then vortexed before use to obtain an even suspension. Fatty acid solutions were also prepared by vortexing each fatty acid in dimethyl sulfoxide (DMSO) (BDH; VWR, West Chester, PA). The maximum concentration of fatty acid-DMSO solutions that was tested was 62.5 mM as DMSO concentrations above 3.125% (v/v) was found to be inhibitory toward Salmonella growth.
Minimal inhibitory concentration determination
A modified resazurin assay was used based on the protocol described by Sarker et al. (2007). The indicator medium for bacterial growth consisted of Iso-Sensitest broth (Oxoid, Basingstoke, UK) with a resazurin dye (Acros Organics) indicator. The pH values of each fatty acid at 1 M concentration, except for pelargonic acid at a concentration of 31.25 mM in Iso-Sensitest Broth, were measured with a Thermo Scientific Orion 2-star Benchtop pH meter (Thermo Fisher, Waltham, MA) (Table 1). The fatty acid stock solutions were diluted in broth to obtain final concentrations of 1 M, 500 mM, 250 mM, 125 mM, 62.5 mM, and 31.25 mM and 90 μL aliquoted into wells of a 96-well plate. The wells were then inoculated with a final concentration of 5 log colony-forming unit (CFU)/mL of Salmonella cells, following the recommended concentration for the broth dilution method to assess the minimum inhibitory concentration (MIC) of antimicrobials (Wiegand et al., 2008). The plates were loosely sealed using cling wrap and placed in an incubator at 37°C for 24 h. The 96-well plates were examined after 24 h for change in color from blue to pink, indicating bacterial growth. Wells that did not change color from blue to pink were indicative of an inhibitory concentration of the fatty acid. Fatty acids that changed the hue of the dye, but did not completely change color, were further evaluated for their effect on Salmonella growth over a 24 h period.
The pH of unamended Iso-Sensitest Broth is 7.20 and unamended TSB is 7.12.
Indicates that fatty acid/broth mixture was not used in this study. Solubility and pKa values are from National Center for Biotechnology Information. PubChem Compound Database; margaric acid pKa is from ChEMBL Database.
TSB, tryptic soy broth.
Growth of Salmonella in the presence of fatty acids
Salmonella Newport, Typhimurium, and Javiana were inoculated into TSB containing 1 M lauric acid (TSBL), 25 mM pelargonic acid (TSBP), or 1 M myristic acid (TSBM). TSBM and TSBL were incubated statically at 45°C in a VWR Digital Heat Block. Growth of the pathogen was monitored over a period of 24 h by plating at 0, 4, 8, 12, 18, and 24 h. Plating was conducted through appropriate serial dilutions in 0.1% peptone using the Miles and Misra method, and plate counts were presented as log CFU/mL. The pH of pelargonic acid at 31.25 mM and lauric and myristic acids at 1 M in TSB was measured (Table 1).
Live/Dead staining
Live/Dead staining was performed using Calbiochem® Live/Dead Double Staining Kit (EMD Chemicals, Inc., Gibbstown, NJ), according to manufacturer's instructions. Briefly, an overnight culture of Salmonella Typhimurium was treated with 30 mM of pelargonic acid for 30 min. The cells were then pelleted by spinning in a centrifuge at 16,100 × g for 3 min (Sorvall Legend Micro 21R, Bremen, Germany). The supernatant was discarded and resuspended in 1 mL of SDW by vortexing to a final concentration of 7 log CFU/mL. To the cells, 150 μL of the membrane permeable green fluorescent Cyto-dye and membrane impermeable red fluorescent propidium iodide dye were added. The cells were incubated in the dark at 37°C for 15 min, following which the cells were placed under a cover slip and viewed using a Zeiss 710 confocal laser scanning microscope system (Carl Zeiss Microscopy GmbH, Germany) with a 40x oil immersion objective. To detect green fluorescence of live cells, a 488-nm argon laser with limits between 493 and 550 nm was used. To detect dead cells emitting red fluorescence, a 561-nm diode-pumped solid-state laser with limits between 616 and 656 nm was used. Dual lasers were used simultaneously to observe the differentially stained bacteria along with 4x magnification to detect changes in cell morphology.
Experimental design and statistical analysis
The resazurin plate assay was replicated thrice with three repeats performed. Populations of Salmonella cells were enumerated at different time points over 24 h, and an average of three repeats for each time point was represented as log CFU/mL. Significant difference between bacterial population at different time points for each serotype in the presence of lauric and myristic acids was evaluated using a mixed model with repeated measures. Fixed effects were serotype, fatty acid, and time, while subject was treated as a random effect, due to repeated measures and nonindependence of the observations. Statistically significant effects at p < 0.05 were further analyzed with Tukey's Honestly Significant Difference using JMP® Pro version 11 (SAS Institute, Inc., Cary, NC). The USDA Integrated Pathogen Modeling Program (IPMP 2013) software (Huang, 2014) was used to model bacterial growth rates under constant temperature based on the Baranyi model (Baranyi and Roberts, 1994). This model can be described by the equation:
where Y 0, Y max, and Y(t) are the S. enterica population in ln CFU/mL at initial, maximum, and time, t, h 0, a dimensionless parameter quantifying the initial physiological state of the cells being studied, and μmax, the maximum growth rate. The Baranyi model was applied in two steps. Each growth curve was analyzed using the model to determine h 0 and μmax. When all growth curves were generated for each treatment, an average of h 0 was calculated to be used in a second application of the model to obtain a new value of μ max. The lag phase duration λ was calculated using the equation λ = h 0/μmax.
Results
MIC values of fatty acids against Salmonella
Water suspension, DMSO solution, and quillaja saponin emulsions of fatty acids were evaluated to assess differences in bioavailability and the effect on MIC of fatty acids against Salmonella enterica subsp. enterica serotypes. No difference in the sensitivity of Salmonella serotypes to fatty acids was detected. The pH of the Iso-Sensitest broth decreased from 7.2 ± 0.03 to 4.99 ± 0.08 and 6.67 ± 0.17 when fatty acids were added, as shown in Table 1. The suspension of pelargonic acid in water resulted in Salmonella growth inhibition at a concentration of 125 mM, but no antimicrobial activity of lauric, myristic, palmitic, margaric, oleic, and stearic acids at a concentration of 1 M was observed (Table 2). The MIC of pelargonic acid suspended in quillaja saponin was 31.25 mM, while inhibition was also noted for lauric acid at a concentration of 1 M against all three Salmonella serotypes tested (Table 3). Myristic acid caused a color change from dark blue to mauve, but not pink after 24 h of incubation. Quillaja saponin based emulsions of palmitic, stearic, margaric, and oleic acids did not result in inhibition of bacterial growth at a concentration of 1 M (Table 3). The modified resazurin dye assay indicated that DMSO at a concentration of 3.125% did not inhibit bacterial growth. Hence, the concentration of fatty acids dissolved in DMSO which could be tested without the solvent inhibiting bacterial growth was 62.5 mM for fatty acids. A MIC of 31.25 mM was observed for pelargonic acid dissolved in DMSO, but no inhibitory activity was seen with lauric, stearic, margaric, myristic, lauric, and oleic acids at a concentration of 62.5 mM (Table 4).
ND- No MIC detected at a fatty acid concentration of 1 M; color change from blue to pink was observed.
Color change from blue to mauve was observed. MIC >1 M denotes that no MIC was detected at a fatty acid concentration of 1 M.
MIC >62.5 mM denotes that no MIC was detected at a fatty acid concentration of 62.5 mM; color change from blue to pink was observed.
Factors affecting S. enterica growth
Presence of the fatty acids lauric and myristic acids (p < 0.001), Salmonella serotype (p = 0.006), and time (p < 0.001) were factors for Salmonella growth (Fig. 1). As expected, all populations increased over the 24 h period, with Salmonella Javiana maintaining a significantly larger population overall compared to Salmonella Typhimurium (p = 0.004). Although serotype was a significant factor, variation was slight and this factor interacted with time (p < 0.001), with differences between these two serotypes ranging from 1.2 log CFU/mL at the 8 h time point to 0.1 log CFU/mL at the 18 h time point. Growth of Salmonella overall was highest in TSB, which differed significantly from growth in TSBL (p < 0.001) and TSBM (p < 0.001). A statistically significant difference was also observed between growth in the presence of lauric and myristic acid (p < 0.001).
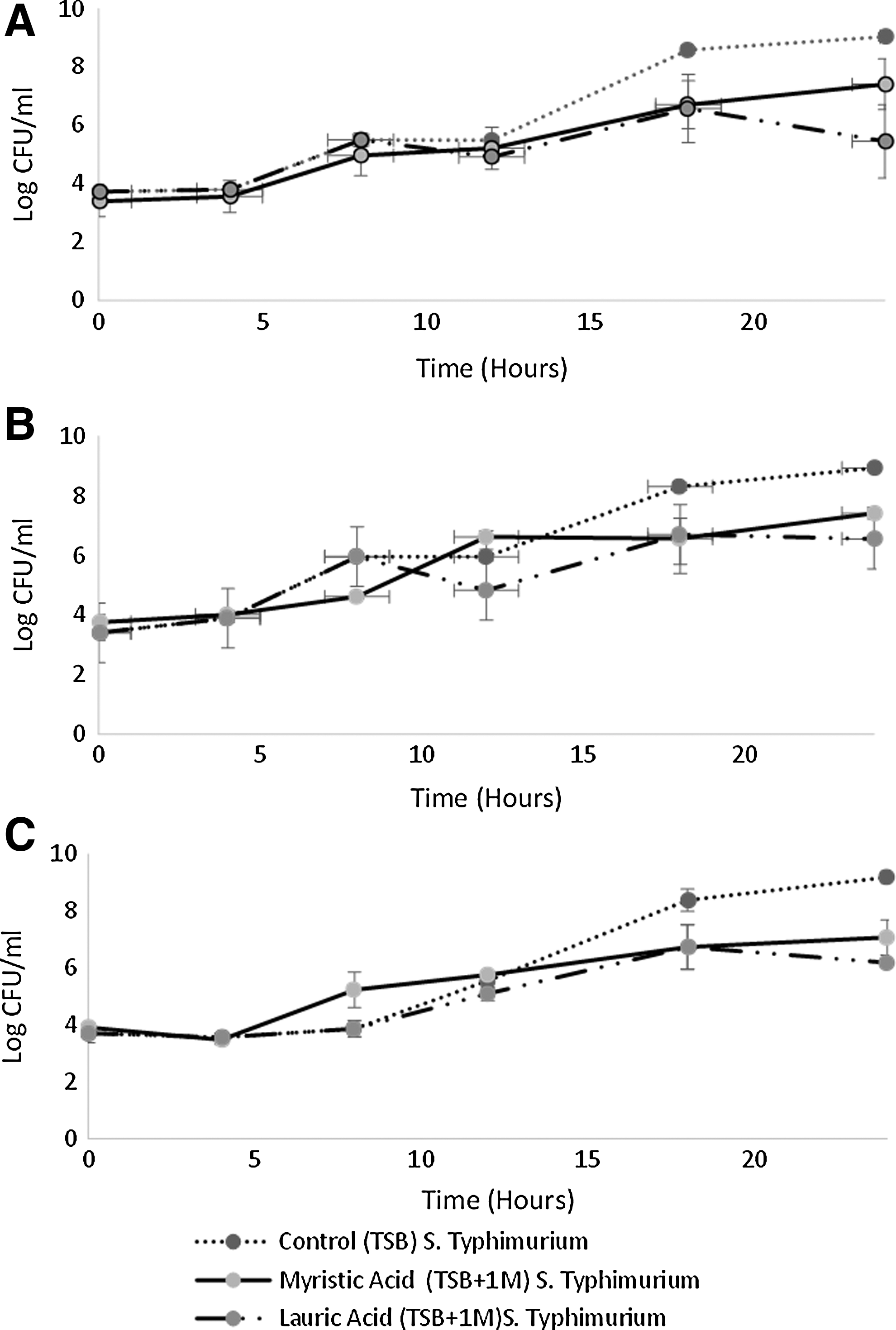
Growth curves of
Effect of lauric acid on S. enterica growth
Lauric acid had an inhibitory effect on all serotypes analyzed (p < 0.001) (Fig. 1). The addition of 1 M lauric acid changed the pH of TSB from 7.12 ± 0.02 to 5.19 ± 0.05 (Table 1). In TSB culture in the absence of fatty acids, the populations of Salmonella Newport, Javiana, and Typhimurium had a mean increase of 5.3 ± 0.1, 5.5 ± 0.1, and 5.5 ± 0.5 log CFU/mL over 24 h. Salmonella Newport, Javiana, and Typhimurium had an average population increase of 1.7 ± 1.2, 3.2 ± 0.5, 1.6 ± 0.2, and 2.5 ± 0.3 log CFU/mL, respectively, in TSBL over 24 h. Statistically significant differences in Salmonella Newport population grown in TSB and TSBL were measured at 18 h (p = 0.004) and 24 h (p < 0.001), with population differences of 2.0 ± 1.2 log CFU/mL and 3.6 ± 1.2 log CFU/mL, respectively. For Salmonella Javiana, statistically significant variation in growth in lauric acid unamended and amended TSB was detected at the 12 h (p = 0.030), 18 h (p < 0.001), and 24 h (p < 0.001) time points with discrepancies of 1.1 ± 0.3, 1.6 ± 0.7, and 2.4 ± 0.2 log CFU/mL, respectively. Salmonella Typhimurium growth also exhibited a statistically significant reduction at 18 h (p = 0.002) and 24 h (p = 0.002) with a difference of 3.0 ± 0.2 and 1.6 ± 0.4 log CFU/mL, respectively.
Growth curves were fitted to the data for Salmonella Newport. Salmonella Javiana and Typhimurium growth in TSBL in the presence of lauric acid and in TSB had no fatty acid controls (Fig. 2; Table 5). Lag phase durations were shorter in TSBL compared to control for Salmonella Newport (p < 0.01) and Javiana (p < 0.001) (Table 5). The maximum population density Y max was lower for all three Salmonella serotypes tested in the presence of lauric acid (p < 0.001) (Table 5). The maximum growth rate for Salmonella Newport and Javiana in the presence of lauric acid was higher than the TSB control (p < 0.001) (Table 5), attributed to early transition into stationary phase before 10 h incubation, as fit by the Baranyi model (Fig. 2E, H). Salmonella Typhimurium did not exhibit differences in lag phase duration or growth rate in the presence of lauric acid.
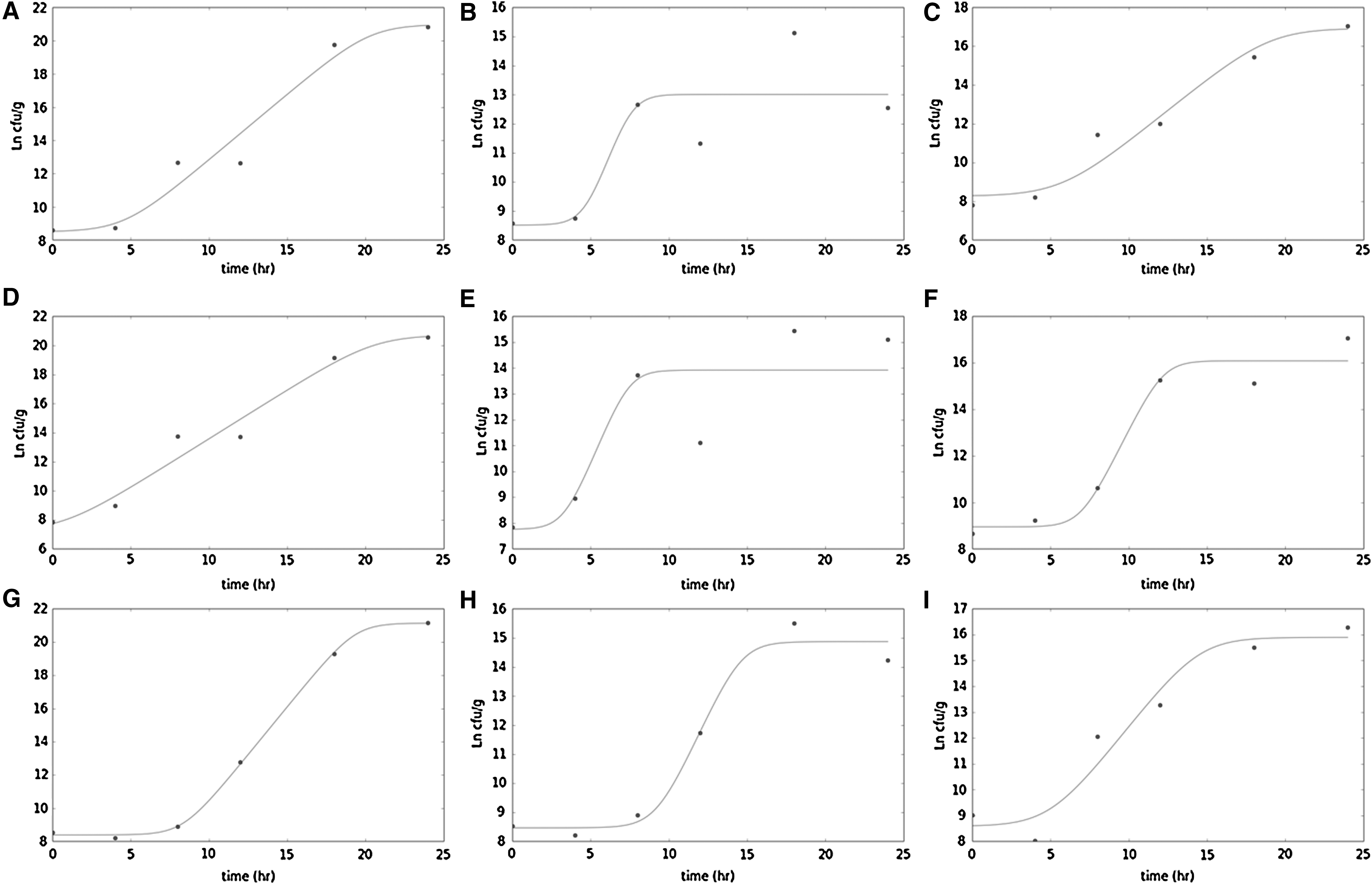
Fitting of Baranyi model to experimental data (shown in Ln CFU/mL) for Salmonella Newport
Superscript letters a,b denote statistically significant differences among treatments within a serotype at p < 0.01. The performance of the model was further assessed using the statistical parameters, the mean of the SSE and the RMSE.
CFU, colony-forming unit; MSE, mean square error; RMSE, root mean square error; SSE, sum of squared error.
Effect of myristic acid on S. enterica growth
Presence of myristic acid in TSBM resulted in significant differences in growth at 45°C among all three Salmonella serotypes tested over 24 h in comparison to growth in TSB unamended with myristic acid (Fig. 1). The addition of 1 M myristic acid changed the pH of TSB from 7.12 ± 0.02 to 5.07 ± 0.18 (Table 1). In the presence of 1 M myristic acid, Salmonella Newport, Javiana, and Typhimurium had an average 4.0 ± 0.4, 3.7 ± 0.5, and 3.1 ± 0.5 log CFU/mL increase in population over 24 h. Statistically significant differences in Salmonella Newport populations grown in TSB and TSBM were measured at 24 h (p = 0.040), with population differences of 1.63 ± 0.2 log CFU/mL. For Salmonella Javiana, statistically significant variation in growth in TSB unamended and amended with myristic was detected at the 8 h (p = 0.004), 18 h (p < 0.001), and 24 h (p < 0.001) time points with discrepancies of 1.3 ± 0.1, 1.7 ± 1.03, and 1.5 ± 0.1 log CFU/mL, respectively. Salmonella Typhimurium growth also exhibited a statistically significant reduction at 8 h (p = 0.020), 18 h (p = 0.002), and 24 h (p < 0.001) with a difference of 1.4 ± 0.90, 1.64 ± 0.4, and 2.1 ± 0.8 log CFU/mL, respectively.
Growth curves fitted to the data revealed differences in Ymax for all three serotypes (p = 0.001) (Fig. 2; Table 5). Lag phase duration varied by serotype, with a significantly shorter lag phase measured for Salmonella Javiana (p < 0.001), while a longer lag phase was detected for Salmonella Typhimurium (p = 0.077) (Table 5). Salmonella Newport did not exhibit any difference in lag phase duration in the presence of myristic acid. Salmonella Javiana displayed a significantly faster growth rate in the presence of myristic acid (p < 0.001), while Salmonella Typhimurium exhibited a slower growth rate (p = 0.067) (Table 5).
Effect of pelargonic acid on S. enterica growth
When 32.5 mM of pelargonic acid was added to TSB, the population of viable Salmonella cells dropped below the detection limit (100 cells) after 1 h. When the medium was tested at 24 h, no cells were enumerated on TSA. To further explore the effect of pelargonic activity of Salmonella Newport, a Live/Dead staining assay was performed on cells after 30 min of exposure to 30 mM pelargonic acid. Imaging using confocal microscopy indicated presence of lysed dead cells along with live bacterial cells (Fig. 3). Average cell length was measured at 5 μm. Elongated cells ranged between 9.74 and 22.65 μm in length, possibly indicating filament formation. Lysed dead cells had red colored granules and did not display yellow fluorescence for typical dead cells.

Confocal laser scanning micrographs of Live/Dead dye treated Salmonella Typhimurium cells after incubation in TSB containing 30 mM pelargonic acid for 30 min. Panel A is at 40x magnification and indicates the presence of
Discussion
The results of this study indicate that S. enterica growth can be affected by the presence of certain fatty acids that were previously identified in tomato fruit surface exudates (Han and Micallef, 2016). Fatty acids could have an antimicrobial effect based on their concentration present in the soluble phase and their bioavailability. Fatty acids are a part of the cuticular matrix of tomatoes and their concentrations and bioavailability could vary based on tomato cultivar and maturity of the fruit. Since their solubility in a solvent could affect bioavailability, in this study three different systems were studied; fatty acids in water which offers limited dissolution, fatty acids in water and a saponin which aids emulsification and, therefore, enhances dissolution, and fatty acids in DMSO which offers higher solubility, but is limited by the concentration of DMSO, which is in itself toxic to Salmonella.
Fatty acids in the soluble phase would be more comparable to fatty acids found in the exometabolome present on the fruit surface. Actual concentrations in the fruit exometabolome are not known; our study mainly utilizes saturated solutions of fatty acids, although the fruit surface may contain lower concentrations. Testing the fatty acids in solution, as an emulsion and as a suspension to understand the impact of bioavailability of fatty acids on Salmonella enterica subsp. enterica growth, we observed that the fatty acid emulsions impacted bacterial growth better than suspensions, suggesting improved bioavailability. Emulsions were created using a saponin as tomatoes contain saponins such as α-tomatine and esculoside A (Weltring et al., 1998; Nohara et al., 2010). Quillaja saponin based emulsions of pelargonic acid resulted in the same MIC as DMSO-dissolved pelargonic acid. While stearic, palmitic, margaric, and oleic acids were not inhibitory at 1 M in all three systems evaluated, pelargonic, lauric, and myristic acids resulted in significant changes in bacterial growth when suspended in quillaja saponin. S. enterica colonization of tomato, therefore, may be impacted not only by the presence and concentration of certain fatty acids but also other compounds present that may enhance the bioavailability of fatty acids that exhibit an antibacterial effect. Interestingly, the bacterial inhibition we observed was seen in the three shortest chained acids we tested, the two medium-chain pelargonic (C9:0) and lauric (C12:0) acids, and a less marked effect by the shortest chained long-chain myristic acid (C14:0). This may be suggestive of decreasing bioavailability with increasing chain length.
Another characteristic that could impact bacterial fitness is the pKa of fatty acids, which affects acid dissociation in a pH-dependent manner. In fact, addition of fatty acids to growth medium resulted in a lowering of its pH approaching the pH of tomato fruit exudates (pH ≈5.5). Only palmitic and pelargonic acid-amended broths had a pH exceeding the pKa of the fatty acid, leading to acid dissociation. However, neither one of these fatty acids exhibited inhibition to Salmonella growth at the concentrations tested, which should have resulted in saturated solutions of the fatty acids in broth. All other fatty acids had approximately equivalent pH-pKa values, such that the concentrations of dissociated and undissociated fatty acids would be in equilibrium.
The modified resazurin assay provided a platform to assay different fatty acids for their ability to inhibit Salmonella growth colorimetrically. The resazurin assay has been previously used to determine the MIC of essential oils and phytochemicals (Mann and Markham, 1998; Sarker et al., 2007). The assay was performed using Iso-Sensitest medium as it is pH buffered. The color change from blue to pink indicated bacterial growth. It was determined through the modified resazurin assay that the concentration at which quillaja saponin was used for this study did not inhibit Salmonella growth. In contrast, concentrations of DMSO above 3.125% exhibited inhibitory activity against Salmonella and limited the concentrations of fatty acid solutions tested in this way. When myristic and lauric acids were evaluated using the Iso-Sensitest medium, the color of the blue resazurin dye became lighter, but did not transition to a bright pink, possibly indicating partial inhibition. These were hence further evaluated by conducting growth curve experiments.
Of the fatty acids tested, pelargonic acid has the lowest molecular weight (158.23 g/mol) followed by lauric (200.31 g/mol) and myristic acids (228.37 g/mol). Pelargonic acid has the lowest melting point at 12°C followed by oleic acid (14.5°C). Microscopic analysis of S. enterica using membrane permeable and impermeable dyes revealed cell damage. Treatment with pelargonic acid appeared to show red staining in and surrounding the cells with the membrane impermeable dye propidium iodide. This observation suggests cell lysis due to cell membrane damage and leakage of cell contents. Pelargonic acid salts are used as herbicides that can disrupt plant cuticle and cell membranes (USDA, 2015). Elongated cells were also observed indicating possible filament formation. Filamentous cell formation is a strategy used by bacteria during DNA damage, antibiotic associated stress, and environmental stress (Bos et al., 2015; Yang et al., 2016).
Microscopic examination of cells treated with 1 M lauric or myristic acids was not possible because the fatty acids solidified at room temperature preventing the isolation of fatty acid treated cells from the medium. Attempts to isolate cells from a 1 M concentration of fatty acids resulted in a film formation due to fatty acid solidification on the microscopic slide. Lauric acid has demonstrated antimicrobial activity against lactic acid bacteria, Propionibacterium acnes, and foodborne pathogens such as S. enterica (Hoffman et al., 2001; Nakatsuji et al., 2009). Myristic acid has demonstrated activity against Gram-positive bacteria and antifungal activity by obstructing the activity of N-myristoyl-transferase (Parang et al., 1996). Other fatty acids such as palmitic acid that were detected in tomato surface washes have also been known to demonstrate antimicrobial activity (Ricke, 2003), but did not result in a color change when the resazurin assay was used.
The growth of Salmonella serotypes was evaluated in TSB for lauric and myristic acids as they demonstrated potential inhibitory activity at a concentration of 1 M. The growth curve was performed at 45°C as the melting points of lauric acid and myristic acid are 43.2°C and 64.4°C, respectively. Although 45°C is a temperature that is higher than what would be encountered in the field, harvested tomatoes may have pulp temperatures as high as 35°C, which can cool slowly in large stacks (Vigneault et al., 2000). Myristic and lauric acids restricted the maximum populations of Salmonella attained after 24 h growth compared to the populations of Salmonella in unamended TSB for all three serotypes tested. Evaluation of growth rates and lag phase duration parameters obtained by the Baranyi model implemented explained some of these population levels. A shorter lag phase and faster growth rate were observed for Salmonella Newport and Javiana in the presence of lauric acid and Salmonella Javiana in the presence of myristic acid. The accelerated growth rate appeared to reflect an earlier entry into stationary phase (shorter exponential phase), a possible effect of toxicity, although some net positive population gains were observed at later time points. These growth curves (Fig. 2B, E, F) were the most poorly fit.
In contrast, Salmonella Newport in myristic acid exhibited a slower growth rate, but no statistically significant difference in lag phase duration, but still attained a smaller population compared to the control. Myristic acid caused a prolonged lag phase duration but decelerated growth rate. Although fatty acids impacted population maxima of all serotypes, differences in growth parameters were serotype and fatty acid specific, suggesting that different serotypes possess different uptake, detoxification, or tolerance capabilities to various fatty acids. This may further explain some of the cultivar-dependent variation in epiphytic S. enterica populations previously measured on fruit of various tomato cultivars (Han and Micallef, 2014). Differences in lag phase between Salmonella serotypes and treatments could have also been affected by the pH of the broths amended with fatty acids. In the case of both lauric and myristic acid, however, the pH of the solution approached the pKa of the fatty acid, which means that the concentrations of dissociated and undissociated acid were equivalent.
Occurrence of S. enterica in the crop growing environment in the Mid-Atlantic region of the United States has been reported repeatedly and certain serotypes may be endemic to specific regions (Greene et al., 2008; Micallef et al., 2012; Angelo et al., 2015; Marine et al., 2015; Maurer et al., 2015). The association of certain serotypes with tomato farm environments and tomato plants may be indicative of adaptation to survive in agricultural environments and association with tomato plants and its edible fruit (Shi et al., 2007). Finding alternative strategies that may be used to enhance tomato protection and reduce tomato contamination may be a viable approach. This study provides evidence that pelargonic acid, as well as lauric and myristic acids, naturally found in tomato exometabolites may impact S. enterica fitness, in a serotype-independent manner. Hence more exploration is required to understand the interaction of S. enterica with the tomato surface and cuticle to develop a better understanding of how fatty acids in tomato may be used as a way to enhance the microbial safety of this fruit.
In conclusion, the ability of fatty acids present in the tomato carposphere to impair fitness or reduce population size of S. enterica points to a need to better understand the composition of the tomato fruit cuticle, the exometabolome, and the interaction of these with epiphytic bacteria. Exploiting naturally occurring compounds on fruit surfaces with inhibitory effects on enteric bacteria could lead to the possibility of developing tomato cultivars that might be more resistant to Salmonella colonization. Selection of tomato cultivars with enhanced resistance to contamination by S. enterica could improve consumer trust in raw produce and reduce the economic burden caused by tomato associated S. enterica outbreaks.
Footnotes
Acknowledgments
This work was supported by University of Maryland start-up funds to Shirley A. Micallef. The authors thank Dr. Wendy Peer for assistance with confocal laser scanning microscopy.
Disclosure Statement
No competing financial interests exist.