
Abstract
This study aimed to investigate the effect of sub-minimum inhibitory concentrations (sub-MICs) of three food-grade biocides (benzalkonium chloride -BZK-, trisodium phosphate -TSP-, and sodium hypochlorite -SHY-) on Salmonella biofilms. The structural parameters and bacterial viability of the biofilms formed by a S. Typhimurium isolate from poultry was investigated by means of confocal laser scanning microscopy after staining with SYTO9 and propidium iodide. The MIC values for Salmonella cells before exposure to subinhibitory concentrations of biocides were 8.0 μg/mL (BZK), 18.0 mg/mL (TSP), and 6.0 mg/mL (SHY). The cultures exhibited a stable acquired tolerance to BZK and SHY. The maximum concentrations of biocides that allowed bacterial growth after several passages through gradually higher concentrations of such compounds were 30.4 μg/mL (BZK) and 10.1 mg/mL (SHY). The architecture and viability of S. Typhimurium biofilms varied in response to sub-MICs of different biocides. Previous adaptation to SHY enhanced (p < 0.001) biofilm formation (average biovolume in the observed field -14,161 μm2-: 139,856.15 ± 155,213.27 μm3) relative to unexposed cells (53,779.05 ± 55,535.62 μm3) and cells previously exposed to BZK (58,216.97 ± 58,644.45 μm3) or TSP (30,052.13 ± 28,290.56 μm3). This was particularly marked when biofilm was grown in the absence of biocides or in the presence of sub-MICs of SHY. The highest percentage of dead cells was shown by biofilms formed by cultures previously exposed to TSP relative to the other conditions tested (34.08% ± 13.74% vs. 23.70% ± 16.16%; p < 0.001). The importance of maintaining higher than MICs of SHY during sanitizing procedures to fight foodborne infections by Salmonella biofilms is highlighted.
Introduction
S
Many foodborne outbreaks have been associated with biofilms (Díez-García et al., 2012). As a result, a great deal of effort is devoted to maintaining efficient hygiene in all food industries and to disrupting the formation of biofilms. Consequently, a variety of biocides, such as benzalkonium chloride (BZK), trisodium phosphate (TSP), and sodium hypochlorite (SHY) are commonly employed in food environments (Dynes et al., 2009; Capita et al., 2014).
Subinhibitory concentrations of biocides could occur as a consequence of improper application (using an inappropriate concentration or failing to spread adequately), unsuitable storage of the formulations (resulting in a decrease in the effective concentration of biocide), or the presence of excessive amounts of organic matter, which is known to inactivate several chlorine-based compounds (Capita et al., 2014). Recent studies have shown that some biocides, when present at concentrations below the minimum inhibitory concentration (MIC), can induce significant biofilm formation in a variety of bacterial species (Houari and Di Martino, 2007; Redelman et al., 2012; Capita et al., 2014; Ortiz et al., 2014; Luther et al., 2015). However, it would appear that BZK, TSP, and SHY have not as yet been tested against Salmonella for this purpose. A study of this sort would provide useful information from the point of view of food safety.
This research aimed to examine whether exposure of S. Typhimurium to subinhibitory concentrations of three food-grade biocides commonly used in food environments (benzalkonium chloride, trisodium phosphate, and sodium hypochlorite), and consequent adaptation to them, could influence the ability of this bacterium to form biofilms. For this purpose, the structural parameters (biovolume, surface coverage, maximum thickness, and roughness) and the bacterial viability of the biofilms produced by a S. Typhimurium strain of food origin were quantitatively assessed by confocal laser scanning microscopy (CLSM).
Materials and Methods
Bacterial strain and culture conditions
A Salmonella enterica serotype Typhimurium strain (S175) previously isolated from poultry was used. The strain was cryopreserved at −80°C in tryptone soya broth (TSB, Oxoid Ltd., Hampshire, England) supplemented with 20% (v/v) glycerol. Before experiments, frozen cells were subcultured twice in TSB at 37°C. Working cultures were kept at 4°C ± 1°C on tryptone soya agar (TSA, Oxoid) plates and were subcultured monthly.
Biocides
Three biocides were tested: benzalkonium chloride (BZK, Fluka, Deisenhofen, Germany), trisodium phosphate (TSP, Merck, Darmstadt, Germany), and sodium hypochlorite (∼10% active chlorine) (SHY, Sigma-Aldrich, Steinheim, Germany). Solutions were prepared aseptically before each experiment in sterile distilled water.
Determination of MICs
The MIC values were established using a micro-dilution broth method (CLSI, 2013). Five colonies of the strain S175 were taken from TSA plates, inoculated into 10 mL of Mueller-Hinton (MH) broth (Oxoid), and incubated at 37°C. Previous experiments had shown that after 24 h of incubation these bacterial cultures contained ∼5 × 108 cfu/mL. For the experiment, 100-well polystyrene micro-well plates (Oy Growth Curves Ab Ltd., Helsinki, Finland) were used. Wells were filled with 20 μL of the chemical solution (a range of concentrations was used for each biocide) and 180 μL of appropriate dilutions (in MH broth) of inoculum to give a final concentration in the well of approximately 5 × 105 cfu/mL. The inoculum concentration was confirmed by plating. The micro-well plates were incubated at 37°C in an automated turbidimetric-based system (Bioscreen C MBR, Oy Growth Curves Ab), which measures absorbance of the culture at regular intervals. Both positive (200 μL of inoculum at 5 × 105 cfu/mL) and negative (180 μL of MH broth +20 μL of chemical solution) controls were included in each experiment. The experiments were replicated five times on different days. The MIC was established as the lowest biocide concentration necessary to prevent growth after 48 h of incubation.
Adaptation to increasing concentrations of biocides
The test was performed in the same manner as described for determining the MIC. The starting concentration of biocide was MIC/2. When growth was observed, 20 μL of the suspension were aseptically transferred to the next well, which contained 160 μL of MH broth and 20 μL of the chemical solution. After the transfer, each well contained a concentration of biocide 1.5 times higher than the previous well. This procedure was continued until no growth was observed after 72 h of incubation at 37°C. The suspension in the last well with recorded growth was streaked on TSA plates with biocide (one-half of the maximum concentration of biocides that supported microbial growth was added to the TSA). Unexposed cells were grown in TSB and subsequently streaked on TSA plates without biocides. After incubation at 37°C for 48 h, agar plates were kept at 4°C ± 1°C for no longer than one week. All groups of cells (unexposed and exposed to BZK, TSP or SHY) were tested simultaneously after the same number of days of storage on agar plates.
Stability of adapted cultures
The stability of adaptive tolerance was determined for each compound by repeated subculturing of S175 cells in nonselective broth (TSB) without biocides. This procedure was repeated every 24 h for 10 days. After 10 days, the ability of strains to grow in the presence of the maximum concentration of biocides that supported microbial growth after exposure to increasing concentrations of biocides was determined.
Biofilm determination
The biofilm study was performed using a method previously described by Capita et al. (2014), with some minor modifications. Briefly, cultures were grown at 37°C for 18 h, and two decimal dilutions in TSB were made to obtain a concentration of ∼106 cfu/mL. TSB with biocide (a concentration of MIC/2 was added) was used for growing and diluting previously exposed cells, while TSB without any biocides was used in the case of unexposed cells. A volume of 250 μL was added to the wells of sterile Matrix® 96-Well Glass Bottom Microplates (Thermo Scientific, New Hampshire). After 60 min of adhesion at 37°C, the wells were rinsed with 150 mM of NaCl to eliminate any nonadherent bacteria before being refilled with 250 μL of culture broth. TSB with biocides (MIC/2) and without biocides was added to all groups of wells (containing unexposed cells and cells previously exposed to BZK, TSP, or SHY). Thus, a total of 16 different conditions were tested. The plate was then incubated for 24 h at 37°C, and the wells were rinsed with 150 mM of NaCl. A volume of 2.5 μL of a 1:1 mixture of SYTO9 (stock 3.34 mM in DMSO) and propidium iodide (PI; stock 20 mM in DMSO) fluorescent dyes from the LIVE/DEAD® BacLight™ Viability Kit (Invitrogen, Carlsbad, CA) was added to 1000 μL of TSB, and 250 μL of this solution were added to each well. The plate was then incubated in the dark at 30°C for 20 min to enable the fluorescent labeling of the bacteria. During CLSM imaging, SYTO9 emits a green fluorescence and is used to identify living microorganisms with intact membranes, whereas PI emits red fluorescence and stains dead bacteria with damaged membranes. Dead and viable cells can hence be identified simultaneously.
CLSM image acquisition was performed using a Nikon Eclipse TE 2000-U confocal laser scanning microscope with EZ-C13.60 software (Nikon Instruments, Inc., New York). The biofilms were observed with a forty magnification (40 × ) lens. Fluorescence was detected by excitation at 488 nm (argon laser), and emissions were collected with a 590/50 (SYTO9) or a 650LP (PI) bandpass filter.
Three stacks of horizontal plane images (512 × 512 pixels corresponding to 119 × 119 μm) with a z-step of 1 μm were acquired for each biofilm at three different regions in the well. Three independent experiments were performed for each condition on different days. Thus, a total of 144 CLSI images were obtained. For image analysis, three-dimensional projections of the structure of the biofilms were reconstructed using the Easy3D function of the IMARIS 7.5 software (Bitplane, Zurich, Switzerland).
The quantitative structural parameters of the biofilms, such as their biovolume, substratum coverage, and roughness, were calculated using the computer program BioRCA 1.4, previously developed by our research group using the Lazarus integrated development environment (IDE), which allowed quantification of the total biofilm population (comprising live and dead bacteria) and the independent subpopulations represented by green (live cells) and red (dead cells) fluorescence. The biovolume represented the overall volume of cells (μm3) in the observation field (14,161 μm2) and provided an estimate of the biomass in the biofilm. Substratum coverage (%) reflected the efficiency of substratum colonization by populations of bacteria. Roughness provided a measure of how much the thickness of the biofilm varied, and is an indicator of biofilm heterogeneity. The maximum thickness (μm) of biofilms was determined directly from the confocal stack images.
Statistical analysis
The quantitative structural parameters of the biofilms were compared for statistical significance using analysis of variance techniques. Mean separations were obtained using Duncan's multiple range test. Data processing was carried out using the Statistica® 8.0 software package (Statsoft Ltd., Tulsa, OK).
Results
Adaptation to biocides
Using a broth microdilution method, the MIC values of Salmonella cells before exposure to subinhibitory concentrations of biocides were 8.0 μg/mL (BZK), 18.0 mg/mL (TSP), and 6.0 mg/mL (SHY). After several passages through gradually increased concentrations of the chemicals, the maximum concentration of biocides that allowed bacterial growth were 30.4 μg/mL (BZK), 13.5 mg/mL (TSP), and 10.1 mg/mL (SHY). Adaptive tolerance to BZK and SHY was stable after repeated subculturing in nonselective broth without biocides (the strain was passed through biocide-free TSB every 24 h for 10 days). No adaptation was observed to TSP.
Biofilm architecture
In biocide-free culture media, a well-established and structurally complex biofilm appeared, with more than 90% of the substratum covered (Fig. 1). Previous adaptation to SHY enhanced (p < 0.001) biofilm formation (biovolume 284,926.63 ± 113,508.33 μm3 in the observed field −14,161 μm2-;) (Fig. 1D) relative to cells not adapted (Fig. 1A) and those previously exposed to BZK (Fig. 1B) or TSP (Fig. 1C) (average biovolume 101,366.59 ± 45,551.57 μm3), and decreased roughness (a negative change in the roughness coefficient indicates a decrease in biofilm heterogeneity).
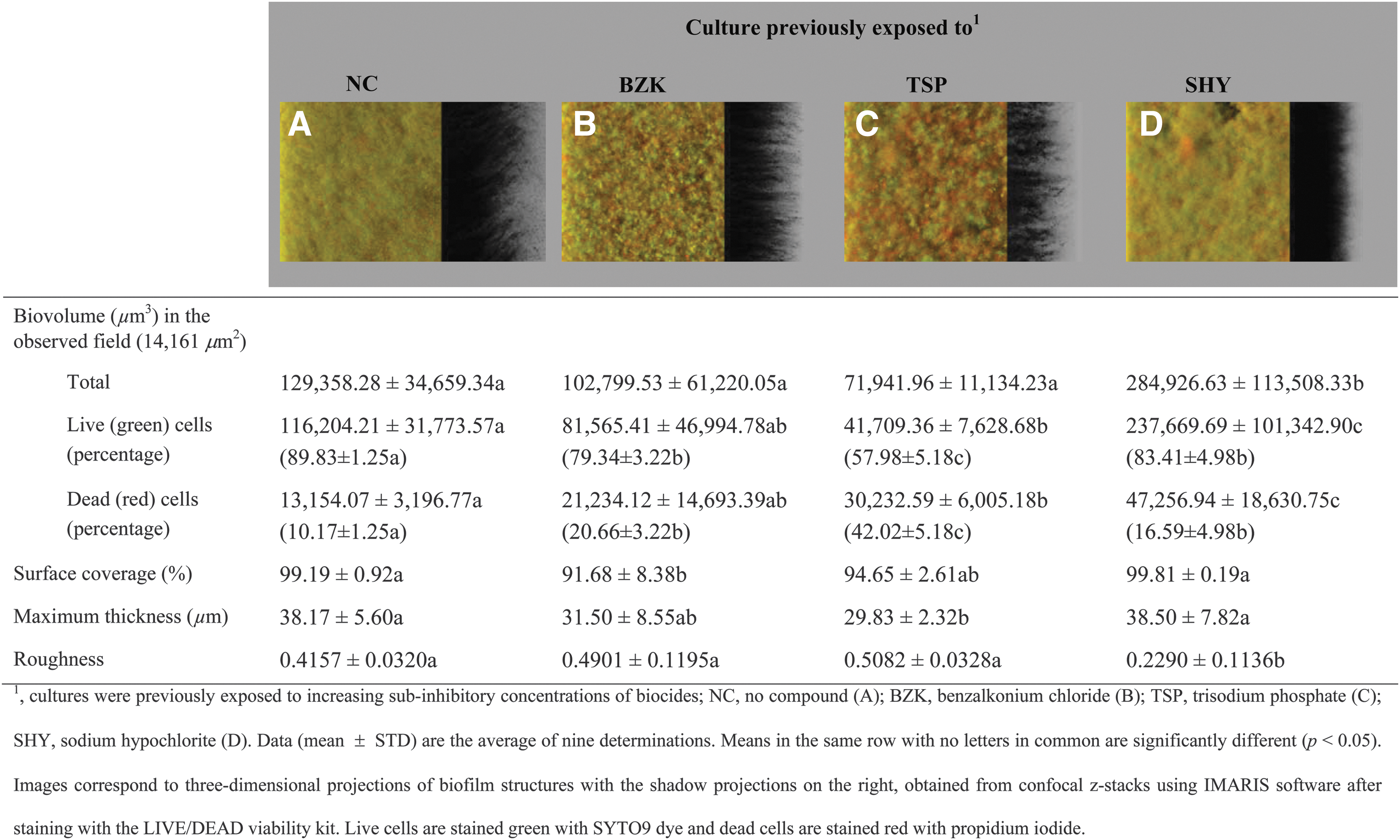
Three-dimensional projections and structural parameters of the biofilms formed by Salmonella Typhimurium S175 in the absence of biocides.
Variation in the three-dimensional structure of the biofilms was observed depending on the compound with which the cells had contacted during the process of biofilm formation. Biofilms grown in the presence of BZK presented a low biomass, with small clusters of live (green) and dead (red) bacteria, regardless of previous adaptation (Fig. 2).
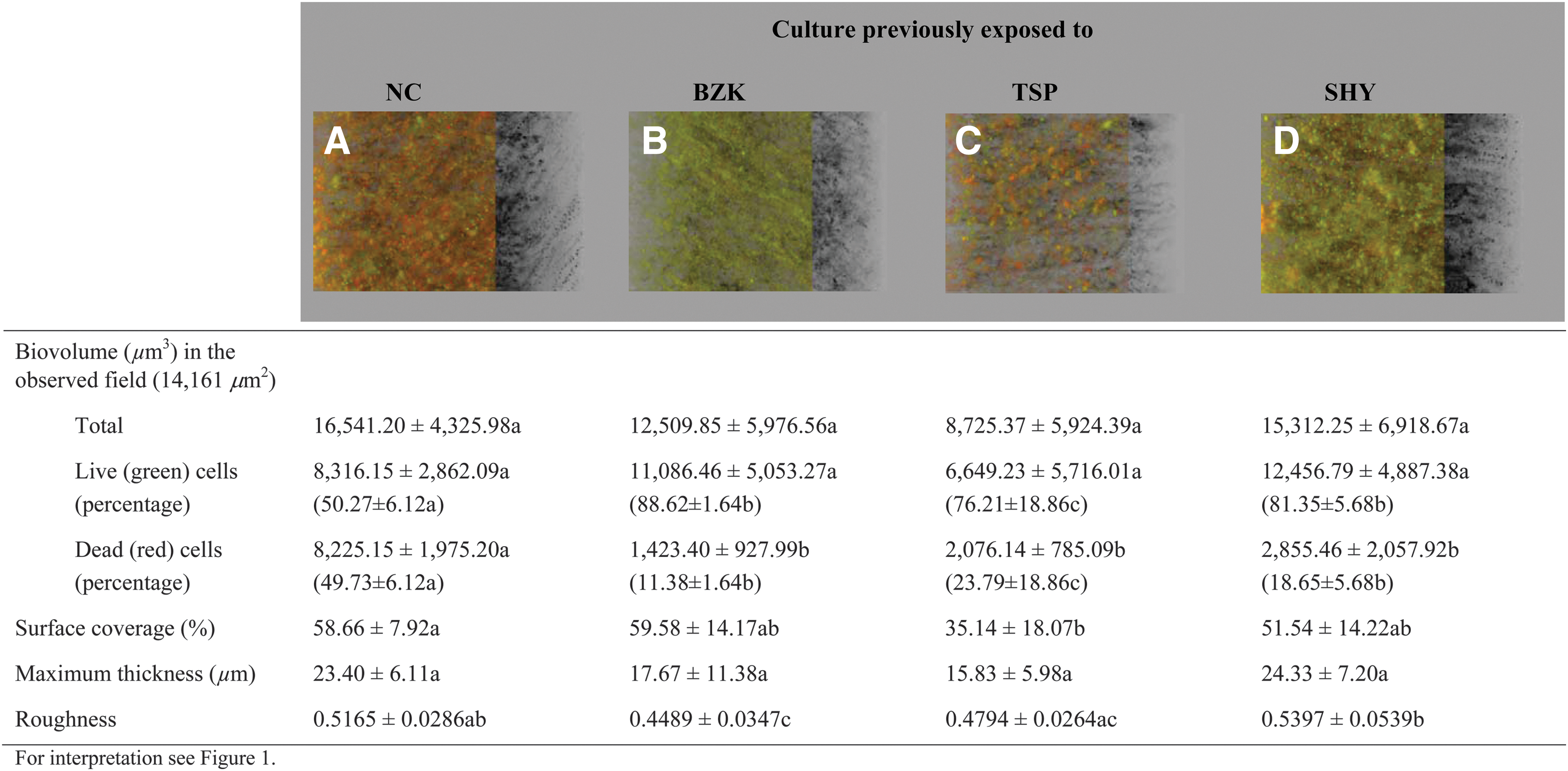
Three-dimensional projections and structural parameters of the biofilms formed by Salmonella Typhimurium S175 in the presence of benzalkonium chloride (MIC/2). MIC, minimum inhibitory concentration.
It was found that when grown in the presence of TSP (MIC/2), unexposed cultures (Fig. 3A) and cultures previously exposed to BZK (Fig. 3B) or TSP (Fig. 3C) formed small micro-colonies of cells that were attached to the glass surface. However, when the cells had previously adapted to SHY (Fig. 3D), the biofilm formed in the presence of TSP covered most of the surface of the observed field, and a larger biovolume (p < 0.01) was found in biofilms from cells adapted to SHY (20,728.15 ± 12,596.98 μm3) than in the other groups of cells, of which the average was 8,831.07 ± 3,981.66 μm3.
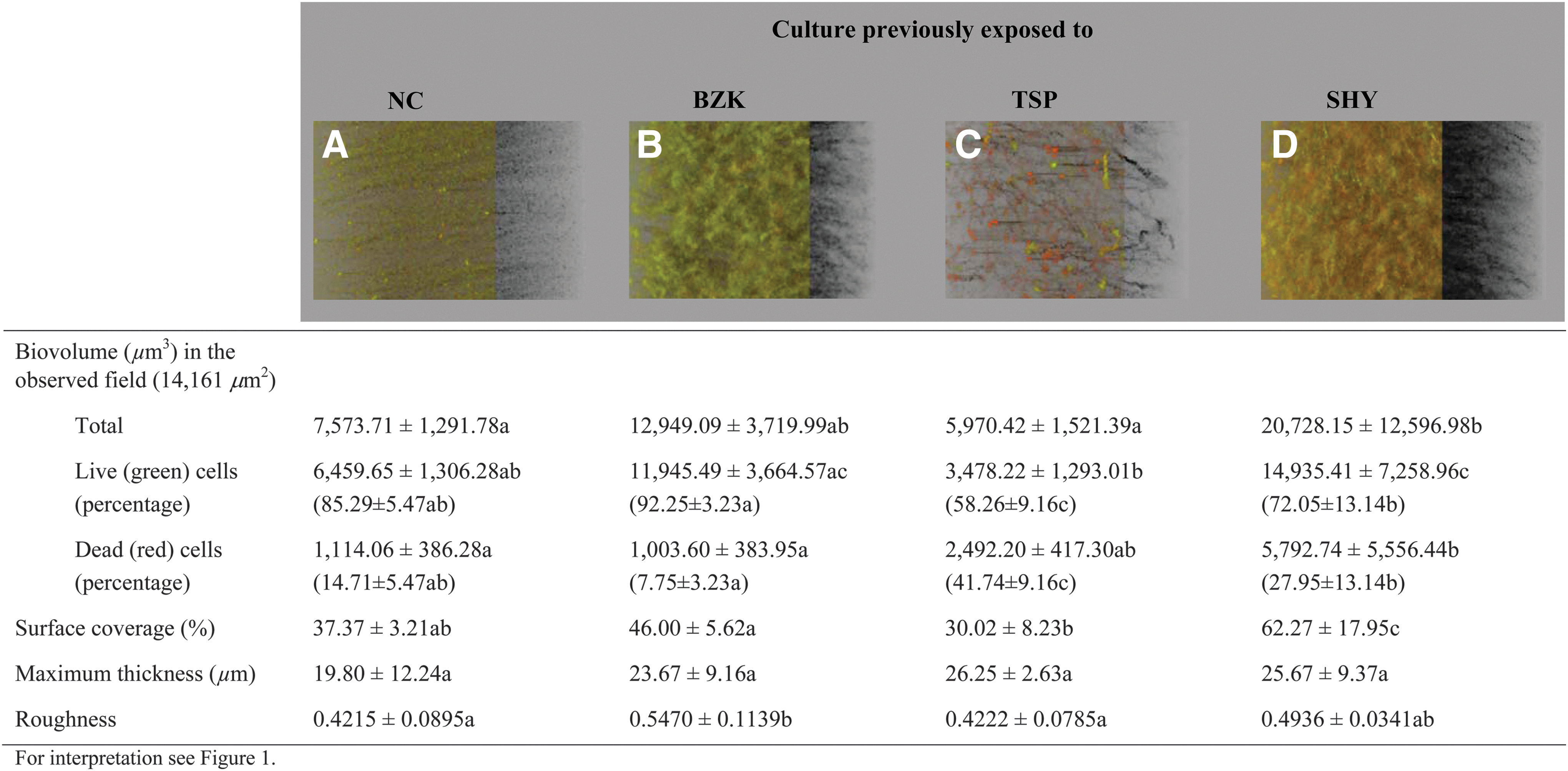
Three-dimensional projections and structural parameters of the biofilms formed by Salmonella Typhimurium S175 in the presence of trisodium phosphate (MIC/2).
The architecture of biofilms growing in the presence of sub-MICs of SHY is shown in Figure 4. Previous adaptation to SHY (Fig. 4D) resulted in an increased (p < 0.001) amount of biomass (238,457.59 ± 158,632.34 μm3) relative to cultures that had no adaptation (Fig. 4A) and to those previously exposed to BZK (Fig. 4B) or TSP (Fig. 4C) (66,607.72 ± 44,065.17 μm3, as average). Previous adaptation to SHY also increased the percentage of surface coverage (97.22% ± 5.65% vs. 80.38% ± 18.42%; p < 0.05), maximum thickness (49.67 ± 11.99 μm vs. 29.45 ± 7.82 μm; p < 0.001), and flattening of the biofilm (0.3904 ± 0.1374 vs. 0.5924 ± 0.1481; p < 0.01).
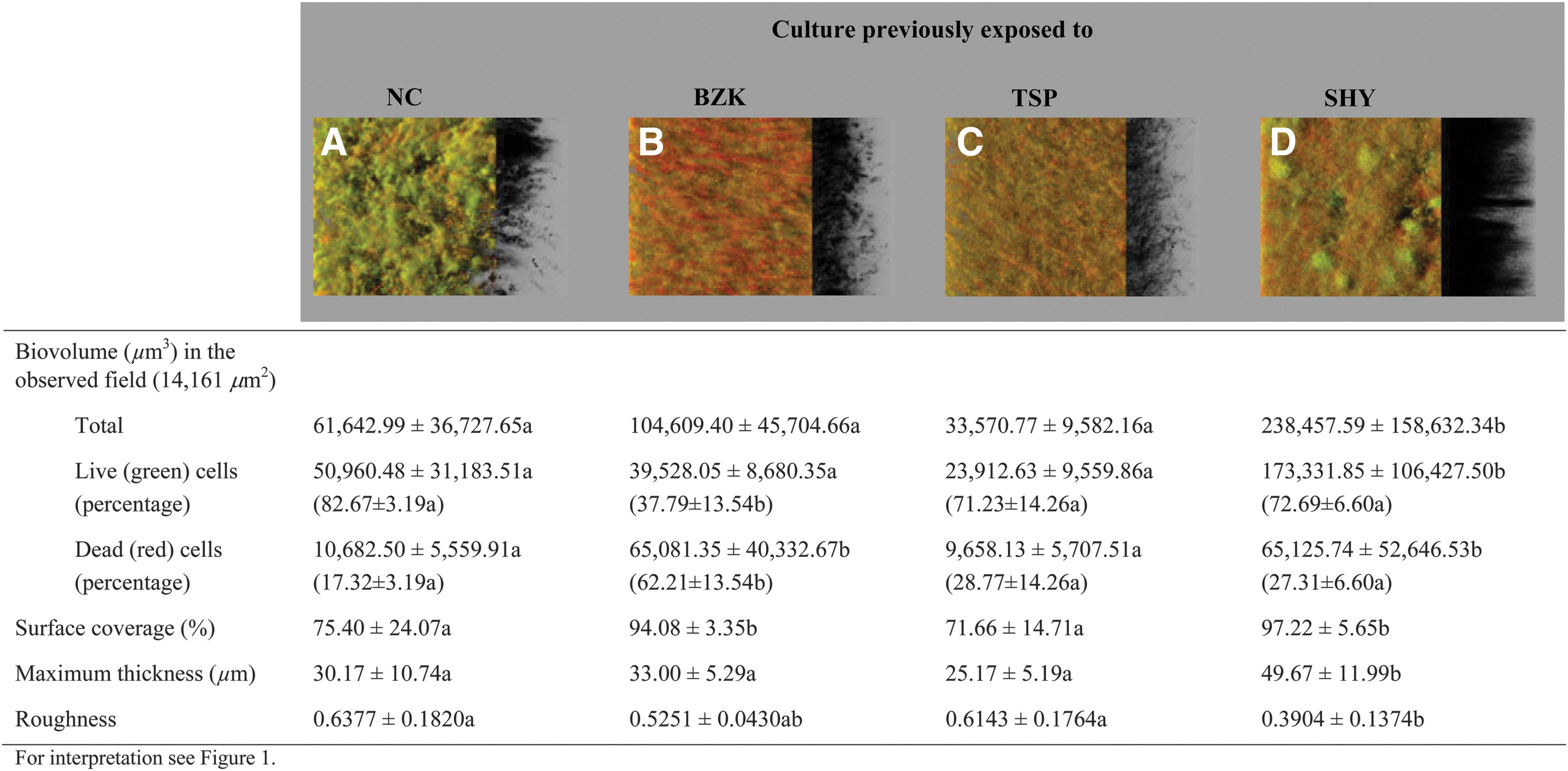
Three-dimensional projections and structural parameters of the biofilms formed by Salmonella Typhimurium S175 in the presence of sodium hypochlorite (MIC/2).
Figure 5 compares the biofilm-forming ability of the strain S175 in the presence of low (MIC/2) concentrations of different biocides. The presence of BZK or TSP significantly reduced biofilm formation by S. Typhymurium, both in unexposed cultures (Fig. 5A) and in cultures previously exposed to BZK (Fig. 5B), TSP (Fig. 5C) or SHY (Fig. 5D). Salmonella cells grown in the presence of SHY had a reduced biofilm-forming capacity relative to cells grown in the absence of biocides in the case of unexposed cultures (Fig. 5A) and those previously exposed to TSP (Fig. 5C). By contrast, cultures previously adapted to BZK (Fig. 5B) or SHY (Fig. 5D) showed a similar (p > 0.05) ability to form biofilms both in the absence of biocides and in the presence of sub-MICs of SHY.
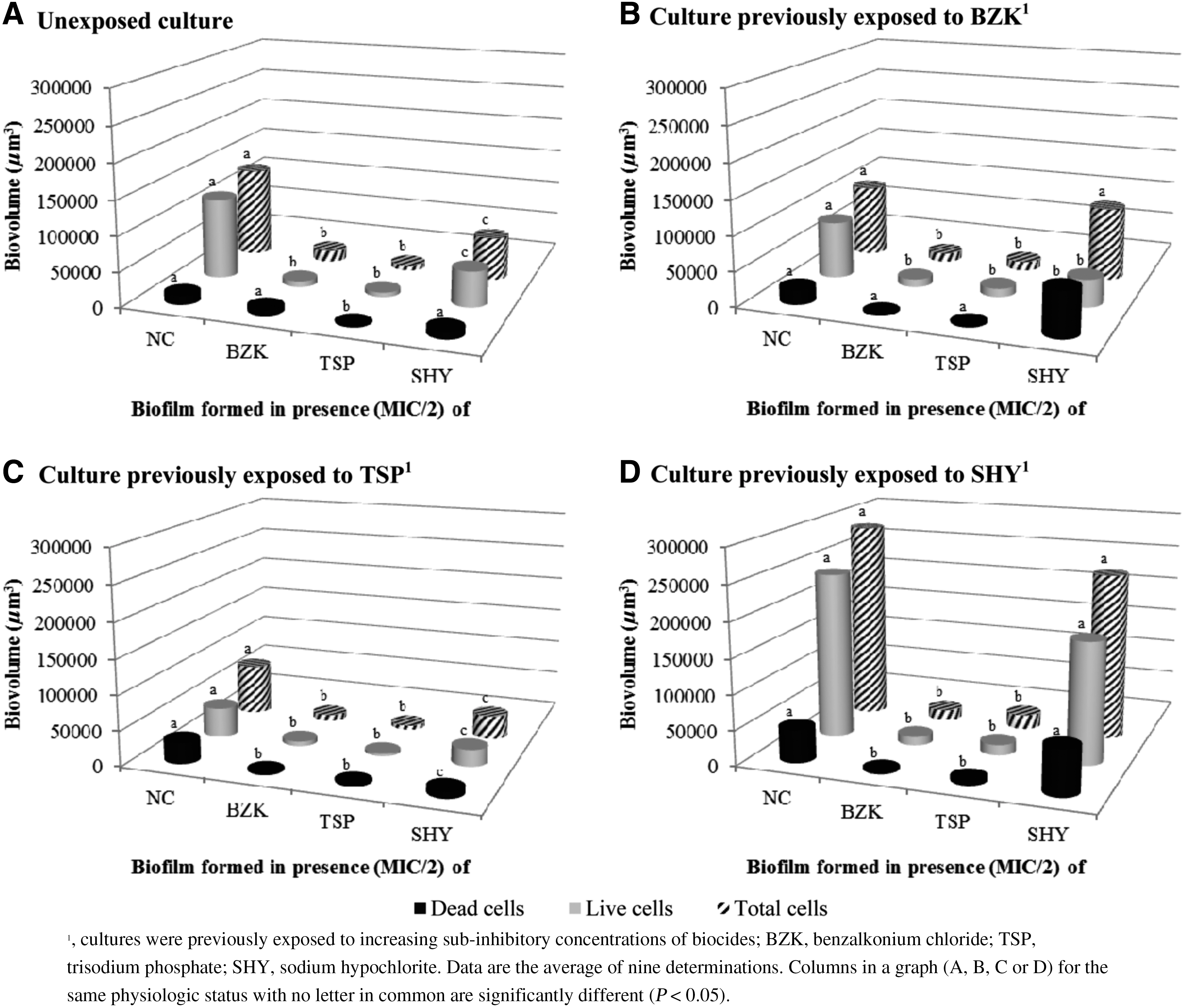
Biovolume of the biofilms formed by Salmonella Typhimurium S175, either previously unexposed
Bacterial viability assessment
Most of the cells in the biofilms were stained with SYTO9 (live cells; 73.70% ± 16.59% of total biovolume). A considerably larger (p < 0.01) percentage of dead cells was observed in cultures previously exposed to TSP (34.08% ± 13.74%) than in unexposed cultures (22.98% ± 16.16%), or in those exposed to BZK (25.50% ± 21.59%) or exposed to SHY (22.63% ± 8.37%). Exposure to biocides did not significantly (p > 0.05) decrease cell viability, with similar percentages of dead cells observed in biofilms growing in the absence of biocides (22.36% ± 12.71%) and in the presence of BZK, TSP, or SHY (average 27.61% ± 17.70%).
Discussion
Salmonella Typhimurium cells undergo adaptation after exposure to increasing subinhibitory concentrations of BZK or SHY. These findings concur with previous observations relating to other genera of bacteria (Dynes et al., 2009; Capita et al., 2014).
In agreement with previous findings (Díez-García et al., 2012), this study demonstrated that S. Typhimurium is able to produce biofilms. Previous adaptation to SHY significantly increased the ability of S. Typhimurium to form biofilms, markedly so when growing in the presence of SHY or in the absence of biocides. This phenomenon is worthy to note, because biofilm formation is thought to play a major role in the persistence of bacteria. It would appear that this is the first report on the effect of sub-MICs of SHY on S. Typhimurium biofilms. The mechanism responsible for increasing biofilm formation in such bacteria after adaptation to SHY remains to be characterized.
As can be seen in Figure 5, the most efficient biofilm formation, both in unexposed cultures and in cultures previously exposed to biocides, was seen for cells grown in the absence of biocides or in the presence of SHY (MIC/2). In contrast, growth in the presence of BZK or TSP decreased the biofilm-forming ability of the strain S175, even though biofilm formation was not completely prevented. Thus, it is suggested that BZK and TSP are in some way able to inhibit bacterial multiplication, attachment, or both, and they might be able to contribute to controlling biofilm formation even at concentrations lower than MIC. In contrast, sub-MICs of SHY were unable to prevent Salmonella from forming biofilms, and even enhanced the biofilm-forming ability of such bacterium. These findings are consistent with previous reports, which showed that sub-MIC doses of SHY stimulate biofilm formation by several bacterial species (Capita et al., 2014; Ebrahimi et al., 2015).
Quantitative analyses of biofilm bacterial viability has been rarely assessed (Fernández-Barat et al., 2012). It would appear that the present research is the first to test the effects of sub-MICs of BZK, TSP, and SHY on the viability of S. Typhimurium biofilms.
The low percentage (27.61% ± 17.70%) of PI-stained cells in biofilms grown in the presence of biocides indicates that membranes of most cells had not been compromised. However, the low biomass observed in the presence of BZK or TSP, as compared with the biofilms grown in the absence of biocides, indicates a reduced ability to form biofilm in the presence of these chemicals. In accordance with other authors (Puga et al., 2016), it is suggested that in the presence of such antimicrobials, bacteria may enter into a dormant state.
Conclusions
The enhancement of biofilm formation after contact with sub-MICs of SHY may have potential implications for food safety, because there are a number of circumstances (e.g., improper use, inappropriate storage, or excessive amounts of organic matter) where concentrations of biocides in food processing environments can be at subinhibitory concentrations. It was highlighted that SHY should be used in adequate concentrations in food industry facilities to avoid sub-MICs of this compound.
Footnotes
Acknowledgments
This work was supported by the Ministerio de Economía y Competitividad (Projects AGL2011-29645 and RTA2014-00045-C03-02).
Disclosure Statement
No competing financial interests exist.