
Abstract
The fern plant Dryopteris uniformis has traditionally been used in herbal medicine and possesses many biological activities. This study was conducted to explore the endophytic bacterial diversity associated with D. uniformis and evaluate their antibacterial potential against foodborne pathogenic bacteria (FPB). Among 51 isolated endophytic bacteria (EB), 26 EB were selected based on their morphological characteristics and identified by 16S rRNA gene analysis. The distribution of EB was diverse in the leaf and the stem/root tissues. When the EB were screened for antibacterial activity against five FPB, Listeria monocytogenes, Salmonella Typhimurium, Bacillus cereus, Staphylococcus aureus, and Escherichia coli O157:H7, four EB Bacillus sp. cryopeg, Paenibacillus sp. rif200865, Staphylococcus warneri, and Bacillus psychrodurans had a broad spectrum of antibacterial activity (9.58 ± 0.66 to 21.47 ± 0.27 mm inhibition zone). The butanol solvent extract of B. sp. cryopeg and P. sp. rif200865 displayed effective antibacterial activity against the five FPB, which was evident from the scanning electron microscopy with irregular or burst cell morphology in the EB-treated bacteria compared to smooth and regular cells in case of the control bacteria. The minimum inhibitory concentration and minimum bactericidal concentration values ranged between 250–500 μg/mL and 500–100 μg/mL, respectively. The above outcomes signify the huge prospective of the selected EB in the food industry. Overall, the above results suggested that D. uniformis contains several culturable EB that possess effective antibacterial compounds, and that EB can be utilized as a source of natural antibacterial agents for their practical application in food industry to control the spread of FPB as a natural antibacterial agent.
Introduction
E
Many studies have shown that endophytes are a novel source of biological products with antagonistic activities that can be utilized by the drug and agricultural industry (Yu et al., 2010; Nair and Padmavathy, 2014; Golinska et al., 2015). Because endophytes remain in a stable environment within the living plant tissue, they have higher bioactivity than microbes isolated from plant surfaces or soil (Selim et al., 2014). Endophytes live inside plants with a great potential source of antimicrobial substances as the plant provides a normal selection system to antibiotics and endophytes (Seo et al., 2010; El-Deeb et al., 2013). To date, a large number of novel biological products with antagonistic activity have been isolated from endophytes (Strobel and Daisy, 2003; Yu et al., 2010). These compounds belong to various structural groups, including steroids, flavonoids, peptides, and terpenoids (Yu et al., 2010). It is widely believed that finding biological products produced by endophytic bacteria (EB) can resolve the problems associated with currently available antibiotics and lead to development of highly effective, low-toxicity, and low environmental impact antibiotics effective against resistant bacterial species (Yu et al., 2010).
Although food is capable of providing nutrients to consumers, it can also support the growth of infectious microorganisms. Indeed, foodborne pathogens comprise one of the most severe threats to human health worldwide (Voravuthikunchai et al., 2006). It has been observed that ferns are not infected by microbial pathogens (Sen and Nandi, 1951; Mandal and Mondal, 2011), and this group of plants has tremendous medicinal and ecological significance. Indeed, ferns are extensively used to treat skin tumefaction, hepatitis, diarrhea, burns, and trauma. The fern plants Polypodium interjectum Shivas, Polystichum woronowii Fomin, Polystichum aculeatum (L.) Roth., Dryopteris affinis (Lowe) Fraser-Jenk, Athyrium filix-femina (L.) Roth, Asplenium scolopendrium L., Asplenium adiantum-nigrum L., and Pteris cretica L. are also known to have antimicrobial, antioxidant, antiviral, antitumor, and even anti-HIV properties (Bahadori et al., 2015). The fern plant Dryopteris uniformis (Makino) Makino is widely known as an ornamental plant, while its young branches are consumed as food in Asia. Although some researchers have identified endophytic fungi from fern plants (Fisher, 1996), the presence of EB internalized in ferns has not yet been investigated in depth (Barros et al., 2010). Moreover, almost no studies have investigated the diversity of EB in ferns or their antibacterial activity. Therefore, this study was conducted to evaluate the diversity of EB in the fern plant, D. uniformis, and to evaluate its antibacterial potential against foodborne pathogenic bacteria (FPB).
Materials and Methods
Collection and isolation of EB
A number of fern plant D. uniformis samples were collected randomly from different locations of the Bohyun Mountain, Yeongchun, Republic of Korea, in May of 2014 and identified by Dr. Seonjoo Park, a plant taxonomist (Department of Life Sciences, Yeungnam University, Republic of Korea). The plant was washed under running tap water, after which 2 g each from leaves (5–10) or stems/roots (5–10 pieces of 1 cm each) were sterilized using 70% ethanol for 60 s followed by 2% sodium hypochlorite for 90 s, 100% ethanol for 30 s, and then three rinses with sterilized distilled water. Sterilized tissues were ground with 6 mL of 0.9% NaCl using a sterile mortar and pestle and incubated to allow the complete release of EB from the host tissues at room temperature for 3 h. For isolation of EB, the tissue extracts were diluted 10- to 100-fold in 0.9% NaCl and plated on Petri dishes having yeast extract and nutrient agar media in three replicates for each dilution (10−1 and 10−2). Finally, samples were incubated for up to 15 days at 28°C.
Identification of EB by morphology and 16S rRNA sequences
The EB isolated from D. uniformis were classified according to their morphological characteristics of the bacterial colony. The EB were finally identified by 16S rRNA gene sequencing. Phylogenetic tree analysis of the isolated EB was conducted using the 16S rRNA sequences. The sequences were aligned and a neighbor-joining tree was constructed. The overhanging ends were detached from both ends to ensure that all sequences were of the same length and the tree was built by the MEGA6 software (version 6) using the neighbor-joining method. Bootstrap tests were performed using 1000 replicates (Tamura et al., 2013; Susilowati et al., 2015).
Screening of antibacterial activity of the isolated EB
Five FPB were investigated in this study, Listeria monocytogenes ATCC 7644, Bacillus cereus ATCC 10876, Staphylococcus aureus ATCC 12600, Escherichia coli O157:H7 ATCC 43890, and Salmonella Typhimurium DT104. These strains were obtained from the American Type Culture Collection (ATCC, Manassas, VA). The evaluation for antibacterial potential was conducted using the patch plate method described by Roh et al. (2009). All the experiments were carried out in triplicate.
Growth kinetics and fractionation by solvent extracts
The EB were cultured in YNB media while shaking at 150 rpm and at 28°C. The absorbance at 600 nm was measured at 24-h intervals from 0 to 96 h using a spectrophotometer (ASP 3700; ACT Gene, Inc., Piscataway, NJ) to evaluate the growth of EB (Araujo et al., 2010). The fractionation of EB extracts was conducted successively using different polarity-based solvents (n-hexane, chloroform, ethyl acetate, and butanol) based on the standard protocol (Zubair et al., 2011). All separated extracts were dried in a rotary evaporator at 55°C, after which 5% Dimethyl sulphoxide (DMSO) was added to the extracts at 0.1 g/mL.
Antibacterial activity of solvent extracts
The antibacterial activity of different solvent extracts was evaluated using the standard disc diffusion method (Diao et al., 2013). The butanol extracts of the two selected EB Bacillus sp. cryopeg and Paenibacillus sp. rif200865 were evaluated for their minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) against five different pathogenic foodborne bacteria by the twofold dilution method (Kubo et al., 2004).
Analysis by scanning electron microscopy
The effects of butanol fraction of the two selected EB, B. sp. cryopeg and P. sp. rif200865, on the morphology of the five FPB were determined by scanning electron microscopy (SEM) (Patra et al., 2015). The foodborne bacteria were then treated with 5% DMSO as the control or with the MIC of butanol faction as the treatment. Immediately before SEM analysis, the specimens were sputter coated with platinum using an ion coater for 2 min before analysis and then examined under a scanning electron microscope (S-4100; Hitachi, Japan).
Statistical analyses
The results of this study are expressed as mean ± standard deviation (SD). Statistical analyses consisted of one-way ANOVA and Duncan's multiple range tests, with p < 0.05 considered to indicate significance. Statistical Analysis Software (SAS) version 9.4 (SAS, Inc., Cary) and Molecular Evolutionary Genetics Analysis (MEGA) version 6.06 were used for all analyses.
Results
Distribution of EB isolated from different tissues of D. uniformis
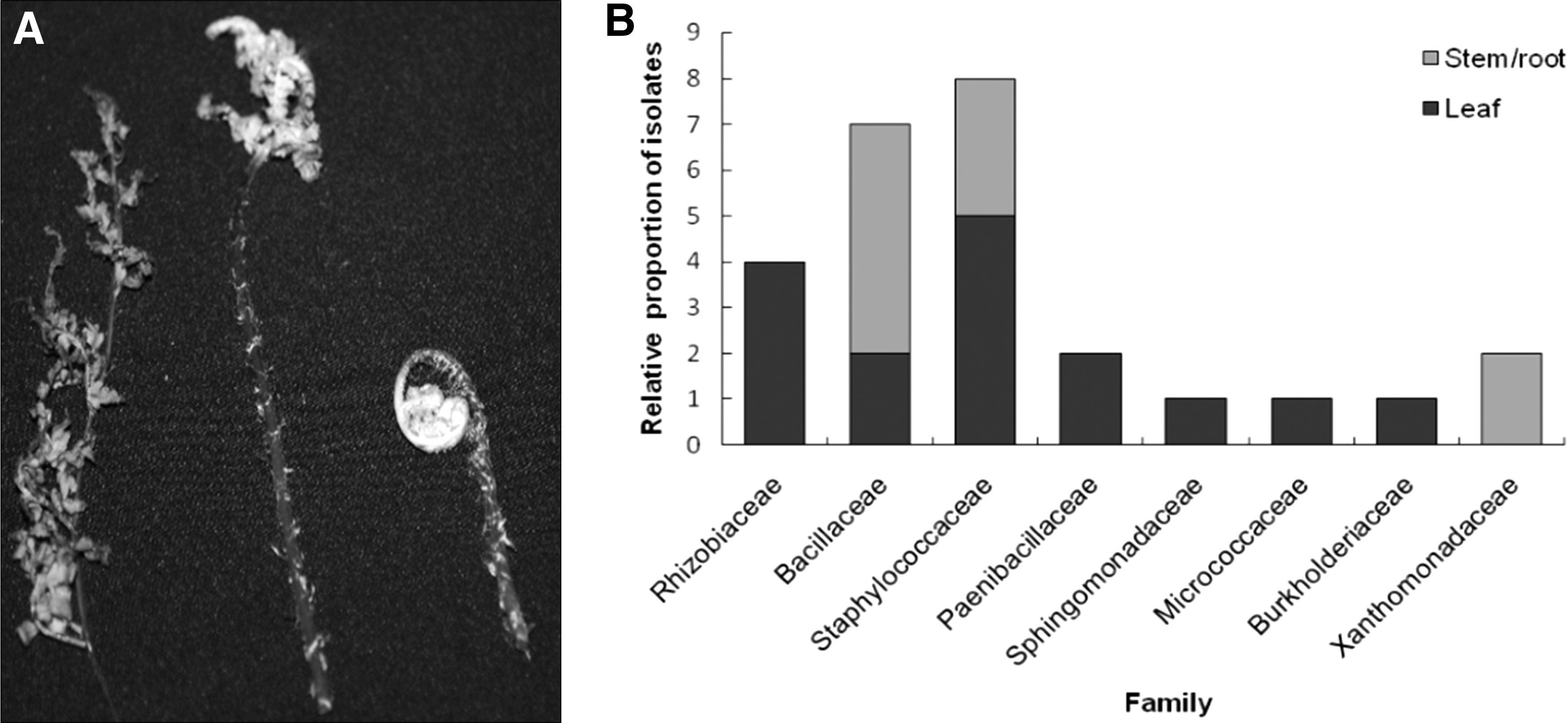
A total of 51 types of EB were isolated from different samples of the plants D. uniformis collected from different locations of Bohyun Mountain, Yeongchun, Republic of Korea (Fig. 1A), based on their morphological characteristics. Out of these, 29 and 22 bacteria species were isolated from leaves and from stems/roots, respectively. The morphological characteristics of isolated bacteria are described in detail in Supplementary Table S1 (Supplementary Data are available online at
Identity%, highest percent of similarity between the query and subject sequences over the length of the coverage area. Maximum score, the highest alignment score (bit-score) between the query sequence and the database sequence segment.
Evaluation of antibacterial activity
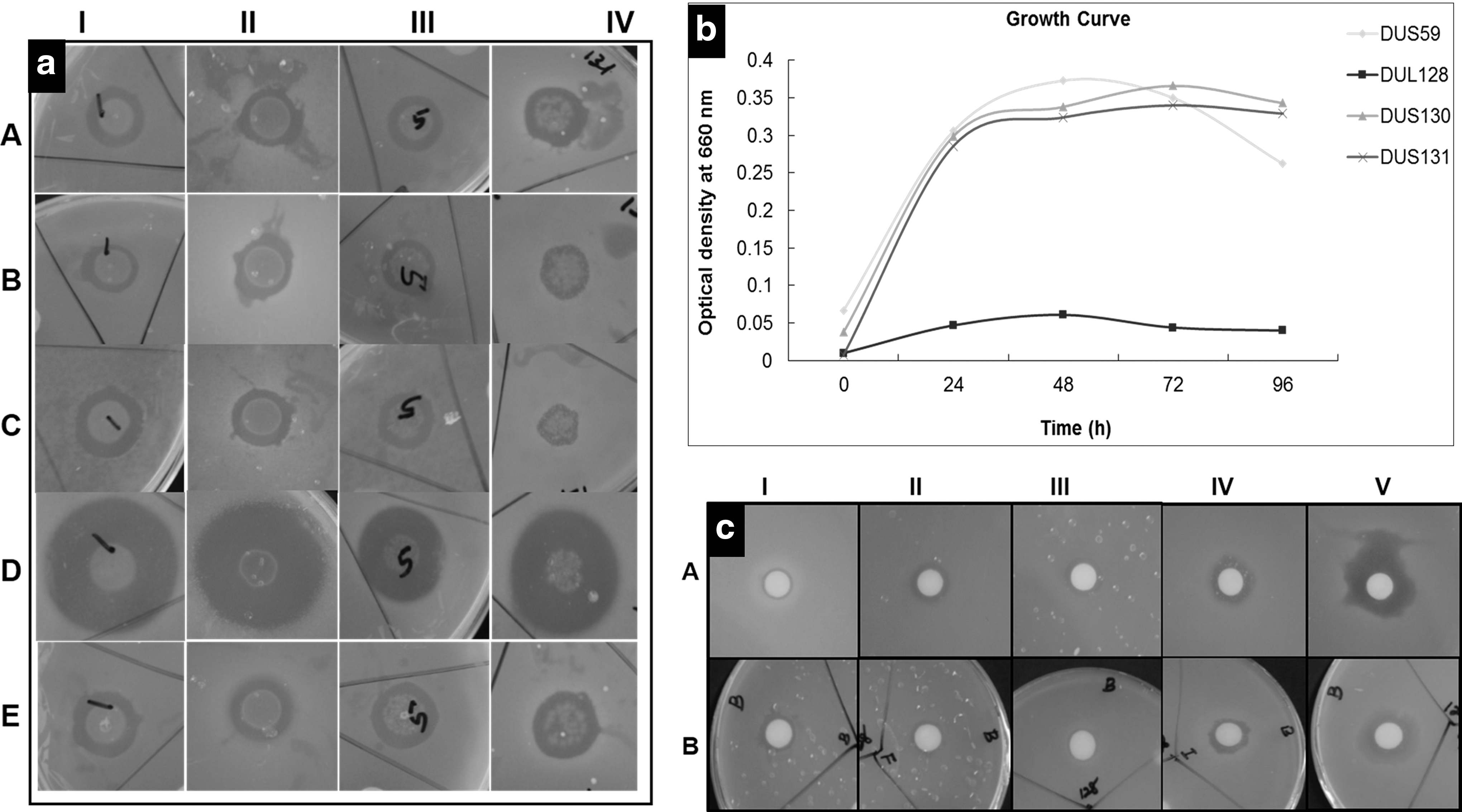
The EB isolated from D. uniformis were evaluated for their antibacterial effects against five FPB. Of all EB tested, four showed broad-spectrum antibacterial activity against the five investigated FPB. Specifically, B. sp. cryopeg (DUS59), P. sp. rif200865 (DUL128), Staphylococcus warneri (DUS130), and Bacillus psychrodurans (DUS131) showed inhibition zones of 13.85 ± 0.30, 11.06 ± 0.36, 11.86 ± 0.77, and 11.36 ± 0.53 mm, respectively, against L. monocytogenes ATCC 7644. However, they showed inhibition zones of 12.56 ± 0.46, 11.60 ± 0.79, 12.59 ± 1.09, and 9.58 ± 0.66 mm against S. Typhimurium, EB B. sp. cryopeg, P. sp. rif200865, S. warneri, and B. psychrodurans, respectively in NA plates. The zones of inhibition of B. sp. cryopeg, P. sp. rif200865, S. warneri, and B. psychrodurans were 15.88 ± 0.09, 14.23 ± 0.10, 12.70 ± 0.33, and 10.75 ± 0.09 mm, respectively, against B. cereus, while they were 23.07 ± 0.11, 19.81 ± 0.03, 18.64 ± 0.07, and 21.47 ± 0.27 mm, respectively, against S. aureus, and 15.47 ± 0.27, 10.11 ± 0.18, 14.33 ± 0.12, and 12.60 ± 0.04 mm, respectively, against E. coli O157:H7. Among the EB, B. sp. cryopeg (DUS59) exhibited the highest inhibitory activity against S. aureus (23.07 ± 0.11 mm inhibition zone), while B. psychrodurans (DUS131) exhibited the lowest activity against S. Typhimurium (9.58 ± 0.66 mm inhibition zone; Table 2 and Fig. 2a). The four EB were assessed for their growth, and the optical density of EB DUL128 was highest at 48 h, while that of the remaining three EB (DUS59, DUS130, and DUS131) was highest at 98 h. Once reaching maximum growth, the growth started declining for all species (Fig. 2b).
Diameter of zone of inhibition expressed as mean ± SD.
Different superscript letters represent significant differences at p < 0.05.
SD, standard deviation.
Evaluation of antibacterial activity of solvent extracts
B. sp. cryopeg (DUS59) isolated from leaf tissue and Paenibacillus sp. (DUL128) isolated from stem/root tissue of D. uniformis were selected for further solvent extraction. The antibacterial activities of these two selected EB solvent extracts were tested against the five FPB. The solvent extracts of concentrated n-hexane, chloroform, and ethyl acetate showed no activity against any of the five pathogenic bacteria, whereas butanol extract exerted different degrees of antibacterial activity against the test organisms. Specifically, the butanol fraction of DUS59 produced zones of inhibition of 10.38–19.09 mm against the five FPB, with the highest inhibition activity being against E. coli 0157:H7 and the lowest against L. monocytogenes (10.38 mm). The DUL128 butanol fraction exhibited zones of inhibition of 9.39–14.35 mm against five tested FPB (Table 3 and Fig. 2c), with the highest being observed against S. aureus and the lowest against B. cereus. The 5% DMSO, which was taken as the solvent control, did not show any inhibition activity against any of the tested pathogens. Phylogenetic tree analysis of the two selected EB was conducted using their 16S rRNA sequences. The neighbor-joining tree showed these two EB to be different genera, with DUS59 and DUL128 being closely related to B. sp. cryopeg and P. sp. rif200865, respectively (Fig. 3).
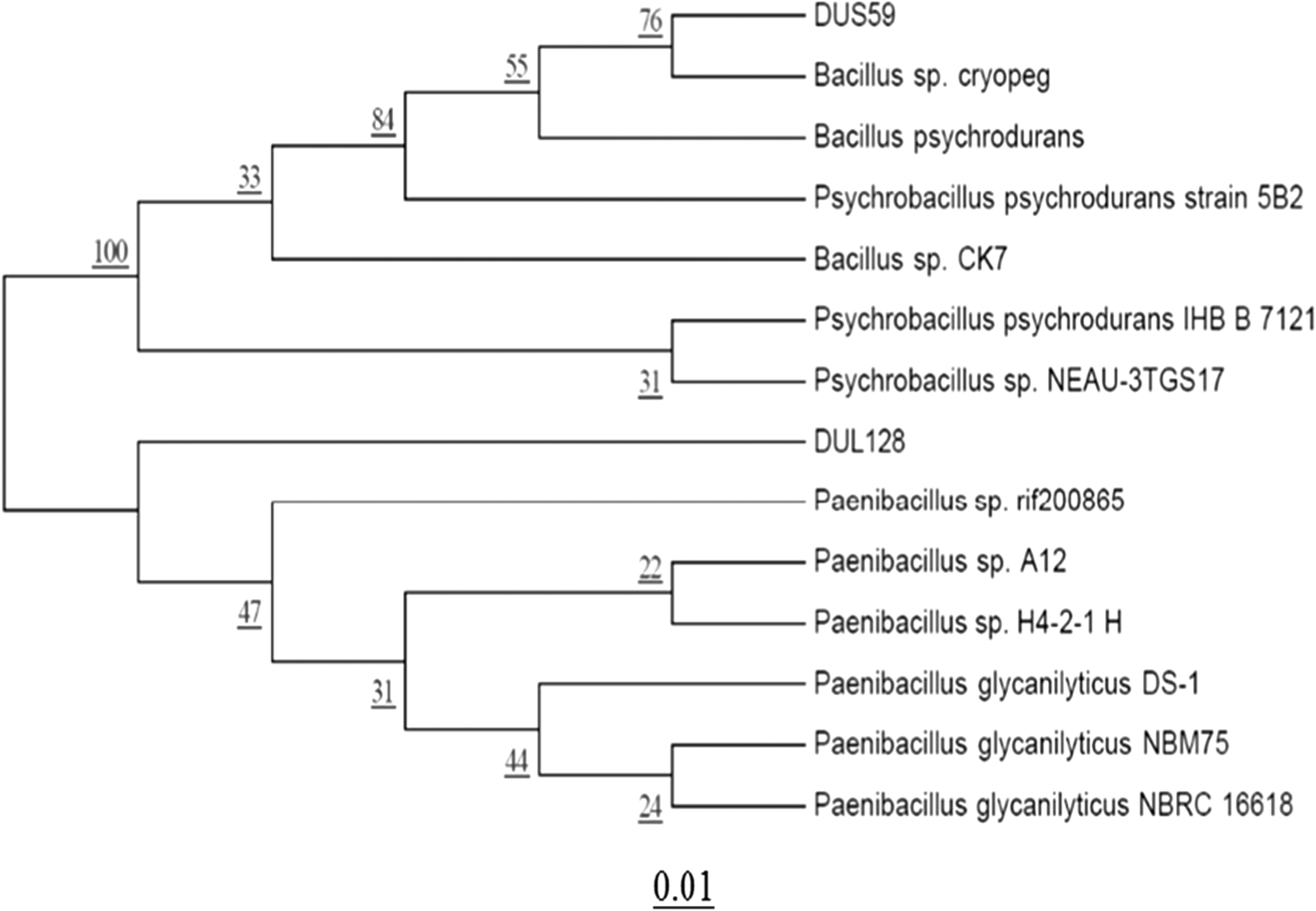
Phylogenetic tree exhibiting similarity among 16S gene sequences of Bacillus sp. cryopeg (DUS59) and Paenibacillus sp. rif200865 with the most closely related species. Evolutionary distances were computed using the maximum composite likelihood method. Bootstrap values (1000 tree interactions) are indicated at the nodes.
Diameter of zone of inhibition in mm expressed as mean ± SD.
Each experiment is carried out in triplicate.
Different superscript letters represent significant differences at p < 0.05.
MBC, minimum bactericidal concentration; MIC, minimum inhibitory concentration; SD, standard deviation.
MIC and MBC of solvent extract
The MIC values of the butanol extract of B. sp. cryopeg were 250 μg/mL against S. aureus and E. coli O157:H7, while they were 500 μg/mL against L. monocytogenes, S. Typhimurium, and B. cereus. The MBC values were 500 μg/mL against B. cereus, S. aureus, and E. coli O157:H7, while they were 1000 μg/mL against L. monocytogenes and S. Typhimurium. In the case of P. sp. rif200865, the MIC values of the butanol extract were 250 μg/mL against S. aureus ATCC 12600, and 500 μg/mL against L. monocytogenes, S. Typhimurium, B. cereus, and E. coli O157:H7. The MBC values were 500 μg/mL against S. aureus and 1000 μg/mL against L. monocytogenes, S. Typhimurium, B. cereus, and E. coli O157:H7. The MIC values of both B. sp. cryopeg and P. sp. rif200865 solvent extracts were 250 μg/mL, while their MBC values were 500 μg/mL against S. aureus ATCC 12600 (Table 3). In the case of B. sp. cryopeg, the MIC value was 250 μg/mL and the MBC value was 500 μg/mL against E. coli O157:H7.
SEM analysis
The effects of butanol extract of B. sp. cryopeg (DUS59) and Paenibacillus sp. (DUL128) on the morphology of five tested pathogenic bacteria were monitored by SEM analysis. The five tested FPB treated with the butanol extract of B. sp. cryopeg and Paenibacillus sp. varied in morphology, having irregular and disrupted cell structures compared with the control bacteria treated with 5% DMSO, which had smooth and regular structures (Fig. 4). The bacteria treated with the butanol extracts of B. sp. cryopeg and Paenibacillus sp. had less in density compared with the control bacteria (Fig. 4).
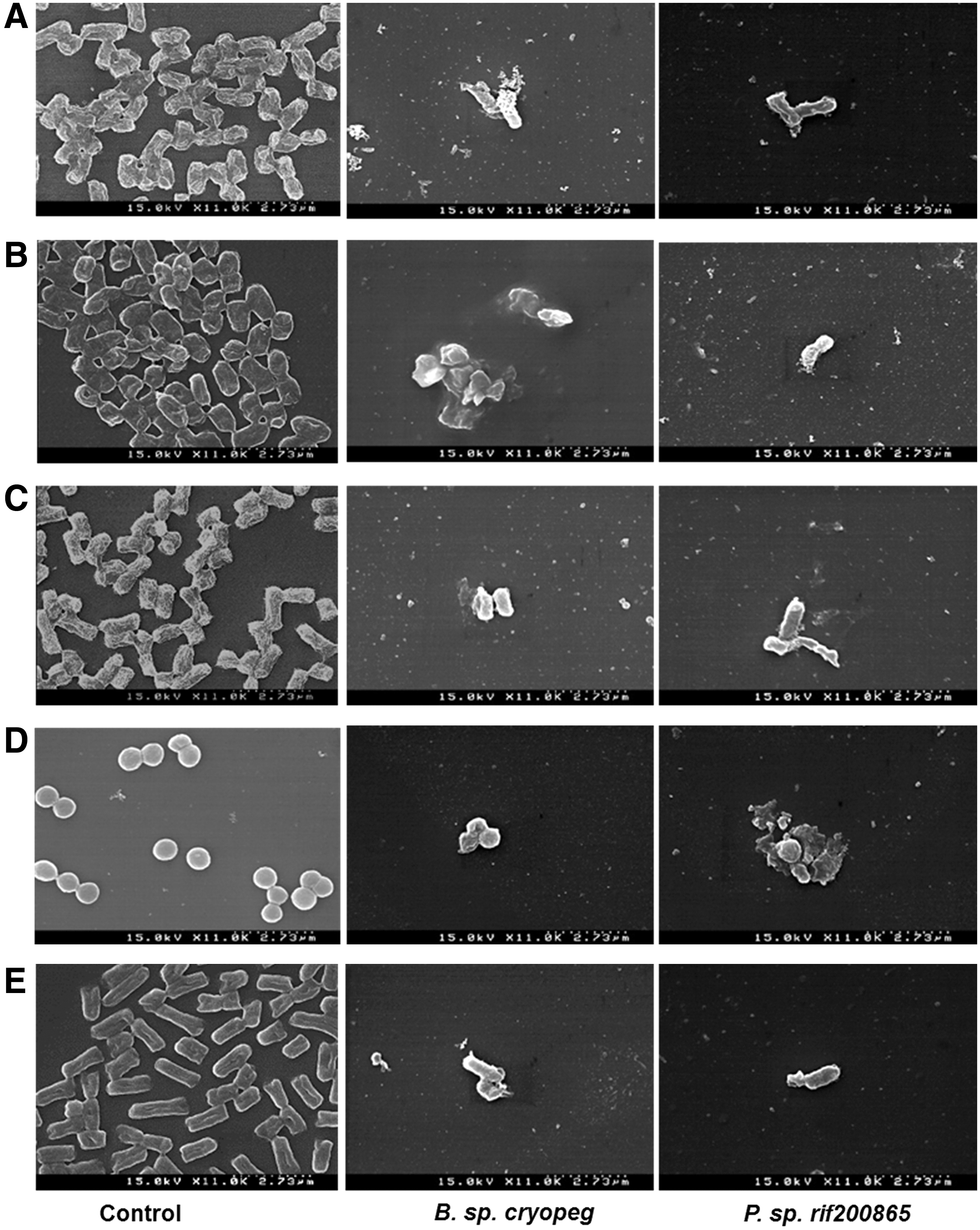
Scanning electron microscopy of the butanol solvent extract of Bacillus sp. cryopeg (DUS59) and Paenibacillus sp. rif200865 (DUL128) against five FPB.
Discussion
To explore the potential applications of EB from D. uniformis, it was necessary to determine the distribution of different EB associated with the host plants. We identified 51 culturable EB associated with leaf (5–10 leaves) and stem/root (5–10 of 1 cm cut) tissues of a number of fern plants, D. uniformis samples collected randomly from different locations of the Bohyun Mountain, Yeongchun, Republic of Korea. There have been many reports of diversity of EB from various other traditional medicinal plants (El-Deeb et al., 2012; Li et al., 2012; Bhore et al., 2013; Sivasankari et al., 2013; Min et al., 2014; Zhao et al., 2015); however, to the best of our knowledge, this is the first study to show diverse types and the applications of EB in D. uniformis. EB can produce valuable metabolites that can be utilized by the pharmaceutical industry because they produce the same metabolites as their hosts (Strobel and Daisy, 2003; Ebrahimi et al., 2010; Mehanni and Safwat, 2010; Preveena and Bhore, 2013).
The presence of microbes in a host plant can be affected by the compounds contained in the host plants (Strobel and Daisy, 2003; Sulistiyani et al., 2014). For these reasons, bacterial endophytes have attracted a great deal of attention owing to their potential for use in isolation of bioactive antimicrobial metabolites (Menpara and Chanda, 2013). The extracts from D. uniformis are well known to have antimicrobial activity (Preveena and Bhore, 2013); therefore, the isolated culturable EB of D. uniformis might produce novel antibiotics. The population diversity of EB varies based on the habitat, stage, and tissues of the host plants. Bacillus, Enterobacter, Microbacterium, Pseudomonas, Stenotrophomonas, Klebsiella, Mycobacterium, Pantoea are the most abundant EB found in plants (Seghers et al., 2004; Sulistiyani et al., 2014). Thirty different bacterial genera from 63 isolates have been identified from the interior of ginseng root (Cho et al., 2007), and 20 different endophytic bacterial genera have been isolated from the meristematic tissues of three varieties of strawberry (Dias et al., 2009). In addition, 49 actinobacteria were isolated from surface-sterilized wheat roots (Justin et al., 2003). These reports confirm that EB populations are noticeably divergent in different plant species and that the diversity of the populations may differ considerably for different locations.
The present study was conducted to isolate EB from the fern plant D. uniformis (Fig. 1A and Supplementary Table S1). The results revealed that D. uniformis is enriched with 51 EB, and the density of EB was higher in leaf tissues than stems/roots, after which 26 EB were identified based on their 16S rRNA sequences (Table 1). When compared with the bacterial distribution in various plant species that were not Pteridophytes found by Seghers et al. (2004) and Sulistiyani et al. (2014), the EB population of D. uniformis is very different, possibly due to it being a Pteridophyte.
FPB are a cause for concern worldwide (Scott, 2003). The major FPB, E. coli, L. monocytogenes, B. cereus, S. aureus, and S. Typhimurium (Hall et al., 2005; Jimenez et al., 2005; Voravuthikunchai et al., 2006; Zarei et al., 2014), have been found to be extremely important to human health and disease. S. aureus and B. cereus are also responsible for many health-related problems (Sofia et al., 2007). The majority of EB are producers of different types of antibiotics (Christina et al., 2013), and the results of the present study indicate that B. sp. cryopeg (DUS59), P. sp. rif200865 (DUL128), S. warneri (DUS130), and B. psychrodurans (DUS131) exerted a broad spectrum of antibacterial activity against L. monocytogenes, B. cereus, S. aureus, E. coli, and S. Typhimurium (Fig. 2 and Table 2). The above four EB were active against Gram-positive and Gram-negative FPB, which is similar to the results reported by Roy and Banerjee (2010).
Among the four EB, two EB, P. sp. rif200865 and B. sp. cryopeg, were further characterized for their antimicrobial capacity by the disk diffusion method using solvent extraction fractions. Basing on literatures (Wise et al., 2012; Kumar et al., 2013; Dutta et al., 2014), butanol was used for extraction of metabolites from the EB. The butanol extract of the two EB showed antibacterial activity against the five FPB (Table 3 and Fig. 2c). The neighbor-joining tree showed that DUS59 and DUL128 belong to two different genera (Fig. 3). SEM analysis revealed that the foodborne pathogens treated with the MIC of butanol extract go through various changes, a loss of cellular integrity and deformations on the surface of the bacterial cell (Fig. 4). When tied with the earlier study, these conclusions proposed that the active compounds of butanol extract might have bound to the surface of bacterial cells, entered the cytoplasmic membrane, and resulted in cell disruption (Fig. 4) (Zhu et al., 2005).
Conclusions
The results of this study demonstrate that D. uniformis contain diverse culturable EB and that some EB produce antibiotic substances that were capable of preventing the growth of FPB. Specifically, the butanol extract of P. sp. rif200865 and B. sp. cryopeg effectively inhibited L. monocytogenes, B. cereus, S. aureus, E. coli, and S. Typhimurium. Accordingly, the isolated EB can be used by the food industry as a natural method to protect food or as sources of novel antimicrobial substances by the pharmaceutical industry. With the increased attention that natural foods and remedies are receiving, the EB isolated from D. uniformis could be a promising source of natural antibacterial substitutes.
Footnotes
Acknowledgment
This work was supported by grants from the Systems and Synthetic Agro-biotech Center through the Next-Generation Bio-Green 21 Program (PJ011117), Rural Development Administration, Republic of Korea.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.