
Abstract
The study objective was to determine effects of fluoroquinolone metaphylaxis on fecal prevalence of Salmonella and Campylobacter and fecal prevalence of quinolone-resistant Salmonella and Campylobacter in feedlot cattle. On Day 0, cattle (n = 288) at risk for bovine respiratory disease (BRD) were randomly assigned to either a nontreated control pen (12 pens) or a fluoroquinolone-treated (enrofloxacin; Baytril® 100) pen (12 pens). Rectal fecal samples were collected from cattle on days 0, 7, 14, 21, and 28. Feces were cultured for Salmonella enterica and Campylobacter spp. using enrichment and selective isolation methods, and confirmed by serology and PCR. Susceptibilities to nalidixic acid and ciprofloxacin were determined using microbroth dilution methods. Data analyses were performed using linear mixed models. Overall, Salmonella sp. and Campylobacter spp. were recovered from 10.2% (139/1,364) and 12.4% (170/1,364) of the fecal samples, respectively. Campylobacter species included hyointestinalis, jejuni, and coli. Neither Salmonella sp. nor Campylobacter spp. prevalence was significantly impacted by fluoroquinolone treatment (p = 0.80, p = 0.61, respectively). However, Salmonella prevalence differed between study weeks (p < 0.01) with prevalence decreasing over time. Before treatment, 98.9% (91/92) of Salmonella isolates were susceptible to nalidixic acid and ciprofloxacin. All Salmonella recovered posttreatment (n = 43) were susceptible to both antimicrobials. The majority of Campylobacter spp. recovered before treatment were resistant to nalidixic acid (23/35; 65.7%) and ciprofloxacin (21/35; 60.0%). There was no significant treatment by week interaction (p = 0.85) or treatment effects (p = 0.61) on the posttreatment prevalence of Campylobacter resistance. There was, however, a significant week effect (p = 0.05), with Campylobacter resistance prevalence decreasing over time. In this 28-day study, we found no evidence that a fluoroquinolone used for metaphylaxis significantly impacts fecal prevalence of Salmonella sp. or Campylobacter spp. or the fecal prevalence of nalidixic acid or ciprofloxacin resistance.
Introduction
S
Human pathogenic organisms, such as Salmonella and Campylobacter, often carry antimicrobial resistance genes (Luo et al., 2003; Chen et al., 2007). As antimicrobial resistance continues to become one of the biggest human health threats around the world, the use of antimicrobials in livestock has become highly scrutinized (Jan et al., 2012; Page and Gautier, 2012). This is particularly true for medically important antimicrobials such as fluoroquinolones, which are used to treat both human and animal diseases (Jan et al., 2012; Page and Gautier, 2012). However, there are minimal data that could be used to estimate the risk to humans associated with the use of fluoroquinolones or other antimicrobials in feedlot cattle.
In the beef industry, the metaphylactic use of antimicrobials is an important tool for reducing bovine respiratory disease (BRD), the most common cause of morbidity and mortality (Bateman et al., 1990; Edwards, 2010). Metaphylaxis involves treating a group of cattle with a U.S. Food and Drug Administration (FDA)-approved antimicrobial with the intent of controlling the incidence of acute-onset BRD in high-risk cattle (Ives and Richeson, 2015). To date, there is no evidence that the metaphylactic use of fluoroquinolone has a negative impact on human health. Prevalence of quinolone resistance in retail ground beef has been very low over the past decade for both Salmonella and Campylobacter (FDA, 2015). However, the continued judicious use of fluoroquinolones in cattle and monitoring of resistant organisms that may be pathogenic to humans seem warranted. There are no published data assessing whether metaphylaxis with a fluoroquinolone, such as enrofloxacin, has any impact on fecal shedding of Salmonella or Campylobacter in feedlot cattle. Therefore, the objective of this study was to assess the effect of using the fluoroquinolone enrofloxacin for metaphylaxis on the fecal prevalence of Salmonella and Campylobacter and the fecal prevalence of quinolone-resistant Salmonella and Campylobacter in feedlot cattle.
Materials and Methods
Study design
Two hundred ninety-eight, 8-month-old, crossbred beef calves were purchased from a livestock auction in Iowa and transported to a Kansas cattle feeder two days before study initiation. To be eligible for this study, calves were required to have had (1) no known history of fluoroquinolone use on sale, (2) no overt clinical signs of illness at arrival, and (3) been considered at high risk for BRD. High-risk cattle are those with one or more risk factors for BRD, which may include auction market sourced, unknown vaccination or preconditioning status, recent weaning, transported from multiple farm origins, extended transport time, or exposure to dramatic temperature changes, or wet or cold weather conditions (Bayer Healthcare, LLC, 2012; Ives and Richeson, 2015).
Cattle were assigned a unique identification number and administered a modified live vaccine (Bovishield Gold®5; Zoetis, Kalamazoo, MI) and a parasiticide (Dectomax®; Zoetis) during processing. Cattle were commingled across all truck loads in a large open feedlot pen and managed according to the feedlot's standard management practices, until study initiation.
Before study initiation, 24 pens, arranged in a circular manner, were randomly assigned to a treatment group. A coin toss determined that the first study pen was assigned to treatment A and the second was assigned to treatment B. Remaining pens were systematically assigned to treatments, so that every other pen was assigned to the opposite treatment. Using the random number function in Excel® 2010 (Microsoft, Redmond, WA), animal ID numbers were then randomly assigned to a pen. The total number of pens, animals per pen, and samples per animal over time for each treatment group were determined via simulation methods to detect a relative difference of 50% between treatment groups over time in the pen-level mean resistance prevalence, with type 1 and type 2 errors ≤0.05 and 0.2, respectively.
On study day 0, cattle were administered either a single subcutaneous 7.5 mg/kg dose of Baytril®100 (enrofloxacin, Bayer Animal Health, Shawnee, KS; treatment A) (n = 144; 12 pens; 12 calves per pen) or no drug (treatment B) (n = 144; 12 pens; 12 calves per pen), according to their randomized assignment; hereafter, groups referred to as treated and control pens, respectively. Body weights were measured before treatment to ensure cattle were properly dosed. Once cattle were assigned to pens and treatment was administered, cattle were placed in their respective pens where they were housed for the duration of the study (30 days).
Cattle health
Routine health observations were conducted daily by the feedlot veterinarian. Cattle that required therapy for noninfectious conditions (e.g., bloat, injury) and did not require an antimicrobial were treated as per the standard site protocol and placed back into their study pen. Cattle that required antimicrobial therapy for any reason, based on the veterinarian's assessment (blinded to treatment group), were removed from the study pen. Once calves were removed from study pens, they were no longer considered part of the study.
Calves were fed a starter ration comprising wet distiller's grain and grass hay. Monensin (Rumensin®; Elanco Animal Health, Indianapolis, IN) was mixed in the ration, per label, to prevent coccidiosis. No tetracyclines, macrolides, or other antimicrobials were included in the feed.
Sample collection
Individual fecal samples (approximately 20 g) were collected from the rectum of all study cattle on study days: 0, 7, 14, 21, and 28 (+/−2 days). Samples were collected using separate latex gloves and placed in WhirlPak bags (Nasco, Inc., Fort Atkinson, WI) prelabeled with the animal ID, study day, and date of sampling. Samples were immediately transported on ice to the Kansas State University preharvest food safety laboratory for same-day processing and analysis.
Salmonella enterica isolation
Ten grams of feces was suspended in 90 mL tetrathionate (TT) broth (Becton, Dickinson Co., Sparks, MD) and incubated at 37°C for 24 h. After incubation, 100 μL of suspension was transferred to 10 mL Rappaport-Vassiliadis (RV) broth (Becton, Dickinson Co.) and incubated at 42°C for 24 h. Enriched samples were then streaked on Hektoen enteric (HE) agar plates (Becton, Dickinson Co.) and incubated at 37°C for 24 h. Three colonies with Salmonella enterica morphology (blue-green colony with black centers) from each sample were streaked onto blood agar (Remel, Lenexa, KS) and incubated at 37°C for 24 h. A single colony was tested by slide agglutination with polyvalent O antisera for Salmonella enterica serogroups O:4 (B), O:6,7 (C1 and C2), O:9 (D1), O:9,46 (D2), and O:3,10 (E). Isolates that agglutinated with the polyvalent O antisera were considered Salmonella sp. Confirmed isolates were stored in cryoprotection beads (Hardy Diagnostics, Santa Maria, CA) at −80°C.
Campylobacter isolation
Campylobacter spp. was isolated from feces using a method described by Burrough et al. (2013). Briefly, fecal samples were suspended at a ratio of 1:10 (1 g feces to 9 mL broth) in Mueller–Hinton (MH) broth (Becton Dickinson Co., Sparks, MD) containing selective (polymyxin B, rifampicin, trimethoprim, and cycloheximide; Oxoid, Cambridge, United Kingdom) and growth supplements (“SS”; sodium pyruvate, sodium metabisulfite, and ferrous sulfate; Oxoid) and incubated under microaerophilic conditions (5% oxygen, 10% carbon dioxide, and 85% nitrogen) at 42°C for 48 h. The enrichment culture was streaked for isolation onto MH+SS agar (Becton Dickinson) and incubated under microaerophilic conditions for 48 h at 42°C. Plates were observed for Campylobacter growth and three, well-isolated colonies per sample were streaked onto MH agar. Plates were incubated at 42°C for 24–30 h under microaerophilic conditions. Pure cultures were then transferred to separate cryovials containing MH broth with 30% glycerol (Fisher, Fair Lawn, NJ). Vials were stored at −80°C until genus confirmation and species identification were performed.
Campylobacter spp. confirmation and speciation
Presumptive Campylobacter colonies were removed from the freezer and streaked for isolation onto MH+SS agar plates. Plates were incubated under microaerophilic conditions at 42°C for 48 h. Approximately two to three colonies were transferred to single-cell lysis buffer (1 mL Tris-EDTA [1 × TE] + 10 μL proteinase K [5 mg/mL]) for DNA extraction (Olah et al., 2006). A multiplex PCR assay (Yamazaki-Matsune et al., 2007) that identifies six species of Campylobacter (jejuni, coli, lari, fetus, upsaliensis, and hyointestinalis subsp. hyointestinalis) was used. Campylobacter jejuni ATCC 33560, Campylobacter coli NCTC 36572, Campylobacter lari ATCC 35222, Campylobacter fetus subsp. fetus ATCC 27374, Campylobacter upsaliensis ATCC 49815, and Campylobacter hyointestinalis subsp. hyointestinalis ATCC 35217 were used as positive controls.
Susceptibility testing of Salmonella enterica
Clinical and Laboratory Standards Institute's (CLSI) microbroth dilution methods were used to determine minimum inhibitory concentrations (MIC) of nalidixic acid and ciprofloxacin for all Salmonella isolates tested (CLSI, 2008). Susceptibility testing was performed on one Salmonella isolate per fecal sample. Stock solutions of nalidixic acid and ciprofloxacin (Sigma-Aldrich, St. Louis, MO) were prepared in sterile, distilled water at a concentration of 1 mg/mL based on the potency of the antimicrobial. Isolates were tested at concentrations of 100, 50, 25, 12.5, 6.25, 3.125, 1.56, 0.78, 0.39, and 0.195 μg/mL for nalidixic acid and 6.25, 3.125, 1.56, 0.78, 0.39, 0.195, 0.098, 0.049, 0.024, and 0.012 μg/mL for ciprofloxacin.
The inoculum was prepared by selecting a single colony from a blood agar plate and mixing it with 10 mL Mueller–Hinton II broth (Becton–Dickinson, Sparks, MD). The inoculum was incubated for 6 h and the cell turbidity was adjusted to 0.5 McFarland turbidity standard. The culture was diluted (1:100) in Mueller–Hinton broth II for an inoculum of 5 × 105 cfu/mL. A 96-well plate was used to perform the antimicrobial susceptibility test for each antimicrobial, and plates were incubated for 24 h at 37°C. Escherichia coli (ATCC 25922) was used as a positive control. Isolates were tested in quadruplicate and results were recorded as either “growth” or “no growth” for each well.
The MIC was the lowest concentration of antimicrobial that inhibited growth of the organism. Salmonella isolates were considered susceptible to nalidixic acid if the MIC was ≤16 μg/mL and ≤0.06 μg/mL for ciprofloxacin. Isolates were considered resistant if the MIC was ≥32 μg/mL or ≥1 μg/mL, for nalidixic acid and ciprofloxacin, respectively (CLSI, 2013). Further testing was performed for Salmonella sp. isolates considered resistant using Sensititre® plates that contained 17 antimicrobials and a semiautomated testing system (Sensititre; TREK Diagnostics, Westlake, OH). Isolates determined to be resistant by microbroth dilution were submitted to the National Veterinary Services Laboratory (NVSL; Ames, IA) for serotyping.
Susceptibility testing of Campylobacter
One Campylobacter spp. isolate per fecal sample was tested using the microbroth dilution method, as described above, at concentrations of nalidixic acid at 100, 50, 25, 12.5, 6.25, 3.125, 1.56, 0.78, 0.39, and 0.195 μg/mL, and concentrations of ciprofloxacin at 25, 12.5, 6.25, 3.152, 1.56, 0.78, 0.39, 0.195, 0.098, and 0.049 μg/mL. Inoculum was prepared by streaking pure culture onto MH+SS agar and incubating under microaerophilic conditions for 48 h at 37°C. One isolate from each plate was transferred to MH agar (without supplements) and incubated for 24–30 h at 42°C. Using a sterile disposable swab, pure culture was transferred to 3 mL MH II broth. Inoculum was adjusted using a spectrophotometer (OD 600) and MH II broth, so that the inoculum was equivalent to 0.5 McFarland standard. The final inoculum was prepared at a 1:100 dilution using MH II broth with 2.5% lysed horse blood.
Antimicrobial susceptibilities were performed using 96-well microtiter plates (Becton and Dickinson, Franklin Lakes, NJ) and a quality control organism, C. jejuni ATCC 33560, which was used as a positive control. Plates were covered using perforated plate covers and incubated under microaerophilic conditions at 37°C for 48 h. Isolates were tested in quadruplicate and considered susceptible if the MIC was ≤16 for nalidixic acid or ≤1 for ciprofloxacin. Campylobacter isolates were considered resistant if the MIC was ≥64 μg/mL for nalidixic acid or ≥4 μg/mL for ciprofloxacin (FDA, 2013).
Statistical analyses
A sample was considered positive for Salmonella sp. or Campylobacter spp. if at least one confirmed isolate was obtained from an individual fecal sample. Descriptive statistics were used to evaluate the MIC data for both nalidixic acid and ciprofloxacin for Salmonella sp. and Campylobacter spp. isolates recovered during this study. The mode of MIC values from isolates tested in quadruplicate was used for each sample. Differences in the pen-level mean prevalence of Salmonella sp. and Campylobacter spp., as well as in the pen-level (nalidixic acid or ciprofloxacin) resistance, between treatment groups were evaluated using generalized linear mixed models, which were fitted using a binomial distribution, maximum likelihood estimation, complimentary log–log link, Kenward–Roger degrees of freedom, and Newton–Raphson and Ridging optimization procedures (Proc GLIMMIX SAS 9.3; SAS Institute, Inc., Cary, NC). The two types of outcomes, evaluated in separate models, included (1) number of Salmonella sp. or Campylobacter spp. test positive samples in a pen (events)/number of samples collected per pen (trials), for each sampling week, and (2) number of resistant isolates per pen/the number of samples collected per pen for each sampling week. Independent variables consisted of treatment (metaphylaxis yes/no), week (n = 4), treatment by week interaction, and a variable representing the baseline prevalence on study day 0 (before antimicrobial administration) for each pen. Random effects, using an autoregressive covariate structure, were used to account for the lack of independence of repeated measures in pens over time. Goodness-of-fit tests were used to examine model fit, and Pearson and deviance residuals were obtained to evaluate potential outliers and influential observations at the pen level.
Results
Baseline cattle weights and fecal shedding
Mean body weights of cattle at enrollment were not significantly different between treatment groups (Table 1). Baseline prevalence of Salmonella was numerically higher in cattle assigned to a treated pen compared with cattle assigned to a control pen, but this difference was not statistically significant (Table 1). The majority of Salmonella sp. isolates belonged to serogroup O:4 (B; 87.1%; 81/93), followed by serogroup O:3,10 (E; 4.3%; 4/93), serogroup O:2 (A; 2.2%; 2/93) and serogroup O:6,7 (C1; 2.2%; 2/93). Four isolates did not test positive for any of the six serogroups evaluated. Susceptibility results revealed that 98.9% (91/92) of the Salmonella isolates recovered before treatment were susceptible to both nalidixic acid and ciprofloxacin. The MIC of the isolate resistant to nalidixic acid was ≥32 μg/mL and was identified as serotype Agona. This isolate was tested against 17 human antimicrobials (amoxicillin/clavulanic acid, ampicillin, azithromycin, cefoxitin, ceftiofur, ceftriaxone, chloramphenicol, ciprofloxacin, gentamicin, kanamycin, nalidixic acid, streptomycin, sulfisoxazole, tetracycline, and trimethoprim/sulfamethoxazole) and was resistant to all but ciprofloxacin, gentamicin, and kanamycin.
For assessing whether means differ between groups.
Baseline prevalence of Campylobacter spp. (before antimicrobial administration) tended to be higher in cattle randomly assigned to a treated pen compared with cattle assigned to a control pen (Table 1). The majority of the isolates across both treatment groups were C. coli (20/35; 57.1%), followed by C. jejuni (13/35; 37.1%) and C. hyointestinalis (2/35; 5.7%). The recovery of C. coli from the feces of enrolled cattle was significantly higher (p = 0.04) in cattle assigned to a treated pen (10.4%; 15/144) compared with cattle assigned to a control pen (3.5%; 5/144) before treatment administration. No difference was found for C. jejuni or C. hyointestinalis (p > 0.05).
Most Campylobacter spp. isolates recovered before treatment were resistant to nalidixic acid (23/35; 65.7%) and ciprofloxacin (21/35; 60.0%). Of nalidixic acid-resistant isolates, 73.9% (17/23) were from cattle assigned to treated pens and 26.1% (6/23) were from cattle assigned to control pens. For ciprofloxacin-resistant isolates, 71.4% (15/21) were from cattle assigned to treated pens and 28.6% (6/21) were from cattle assigned to control pens. Mean pen-level (nalidixic acid) resistance prevalence, before treatment, was significantly higher for cattle assigned to treated pens (11.8%) when compared with cattle assigned to control pens (4.2%; p = 0.03) and also tended to be different for ciprofloxacin (p = 0.06) for treated pens (10.4%) versus control pens (4.2%), despite the fact that all of these samples were collected before treatment administration.
Postantimicrobial animal removals
Within the four study weeks, 10 clinically ill cattle were removed from treated pens and 17 cattle were removed from control pens. All cattle were removed due to clinical signs of BRD, with the exception of two cattle that were removed from control pens due to lameness. Three mortalities occurred during this study (1 from treated, 2 from control pens). Necropsies indicated that all deaths were attributable to BRD.
Postantimicrobial administration—Salmonella prevalence
Overall sample-level prevalence of Salmonella sp. across both treatment groups, posttreatment, was 4.3% (46/1076). Salmonella sp. prevalence did not differ significantly between the treated versus the control pens of cattle (p = 0.80; Fig. 1). The treatment by week interaction was not statistically significant (p = 0.41), however, prevalence did vary by study week (p < 0.01), with Salmonella prevalence decreasing over time for both treatment groups. Most Salmonella sp. isolates recovered, posttreatment, belonged to serogroup O:4 (B; 67.4%; 31/46), followed by serogroup O:3,10 (E; 8.7%: 4/46) and serogroup O:6,7 (C1; 4.3%; 2/46). Nine isolates did not test positive for any of the six serogroups tested.
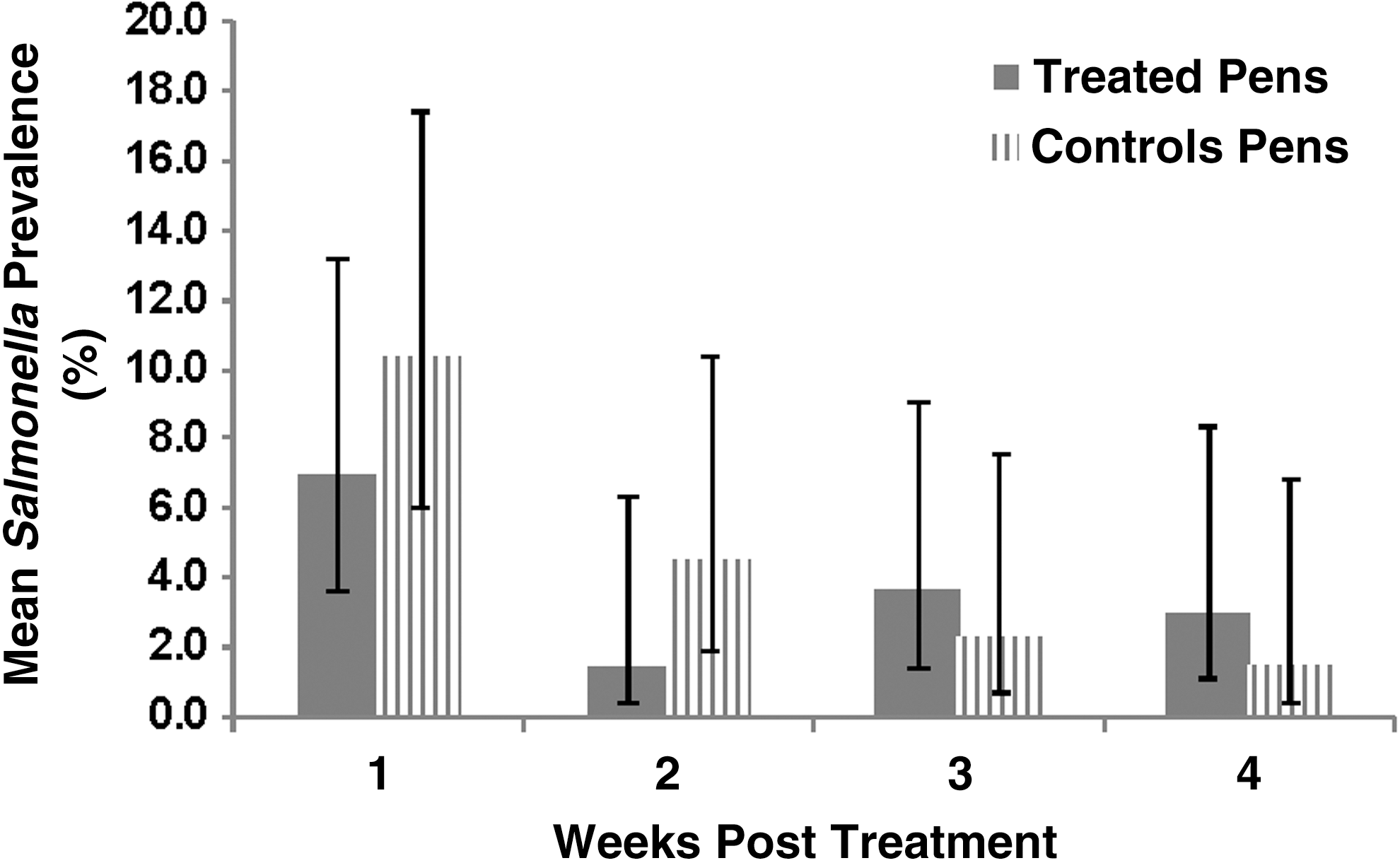
Model-adjusted mean pen-level Salmonella prevalence (and 95% confidence intervals) by week posttreatment, for cattle in fluoroquinolone-treated and nontreated control groups. There was no significant interaction between time and treatment group, or a significant difference between treatment groups, but prevalence across both groups differed over time.
Postantimicrobial administration—Salmonella susceptibility
Of 46 isolates confirmed to be positive for Salmonella sp., 43 (93.5%) were viable and subjected to susceptibility testing. All tested isolates were susceptible to both nalidixic acid (MIC ≤16 μg/mL) and ciprofloxacin (MIC ≤0.06 μg/mL). Most of the isolates had an MIC of 6.25 μg/mL (83.7%; 36/43) for nalidixic acid and an MIC of 0.024 μg/mL for ciprofloxacin (74.4%; 32/43). No isolate was above the breakpoint for either antimicrobial tested.
Postdrug administration—Campylobacter prevalence
Overall Campylobacter spp. sample-level prevalence across all study pens, posttreatment, was 12.5% (135/1076). There was no significant difference (p = 0.65) in Campylobacter spp. prevalence in pens treated with enrofloxacin compared with the nontreated control pens (Fig. 2). There was no significant treatment by week interaction (p = 0.62) nor was there a significant difference by study week (p = 0.10).
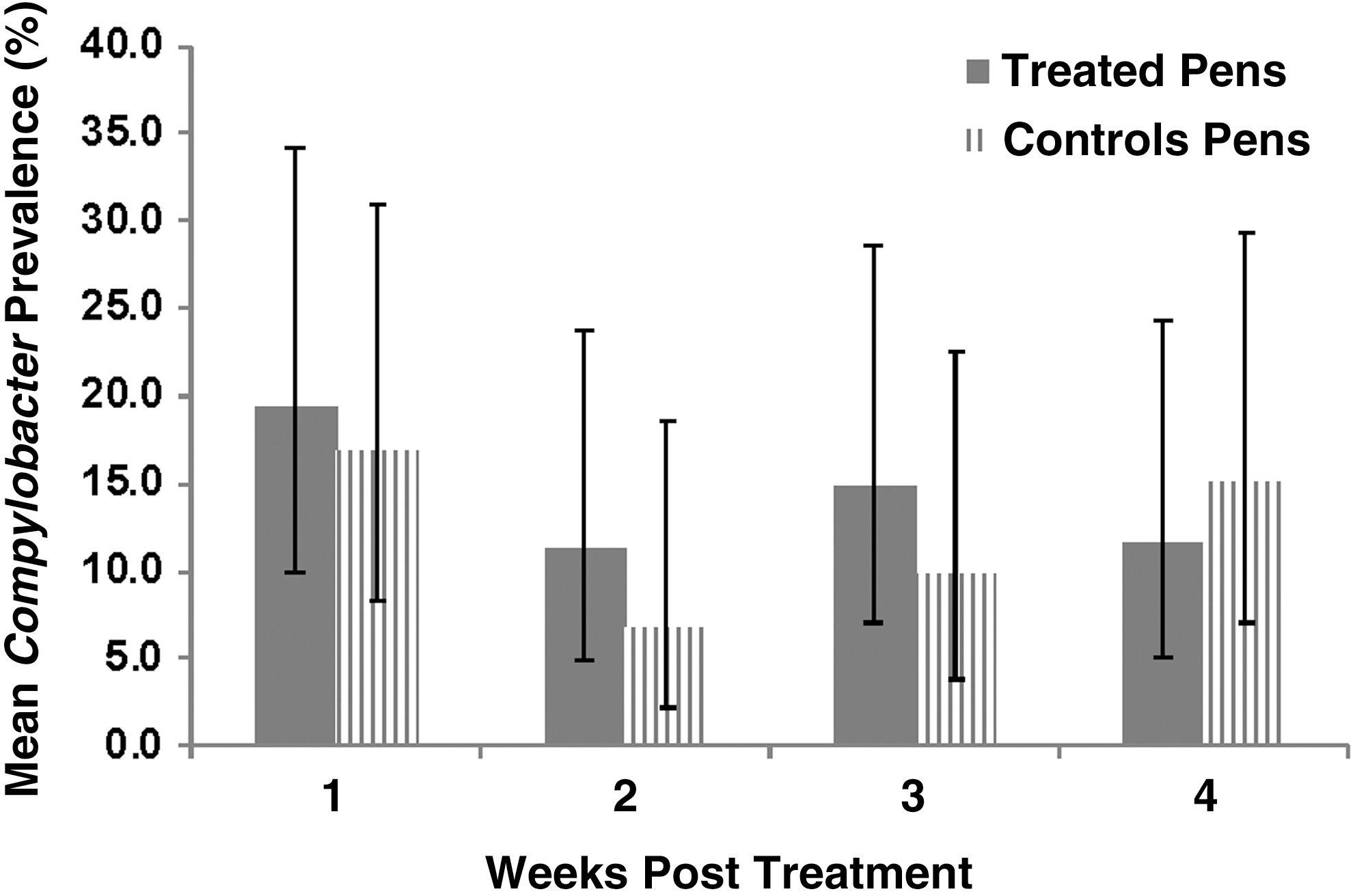
Model-adjusted mean pen-level Campylobacter prevalence (and 95% confidence intervals) by week posttreatment, for cattle in fluoroquinolone-treated and nontreated control groups. There was no significant interaction between time and treatment group, or significant differences between treatment groups or among weeks.
Of the Campylobacter spp. isolates recovered, C. coli accounted for 78.5% (106/135) and C. jejuni accounted for 21.5% (29/135) of isolates. Model-adjusted pen-level prevalence of C. coli was significantly higher in treated pens compared with the control pens (p = 0.04). There was no significant treatment by week interaction (p = 0.88), but there was a significant week effect (p < 0.01). Mean C. jejuni prevalence was not significantly different (p = 0.70) in the treated pens compared with the control pens nor was there a significant treatment by week interaction (p = 0.80) or a week effect (p = 0.07), posttreatment.
Postantimicrobial administration—Campylobacter susceptibility
All 135 Campylobacter spp. isolates obtained posttreatment were viable and subjected to susceptibility testing. Nalidixic acid and ciprofloxacin MICs for these isolates ranged from 1.56 μg/mL to ≥100 μg/mL for nalidixic acid and from 0.049 μg/mL to ≥6.25 μg/mL for ciprofloxacin. Of isolates tested, 81.5% (110/135) were resistant to both nalidixic acid (MIC ≥64 μg/mL) and ciprofloxacin (MIC ≥4 μg/mL). One hundred four of these isolates were C. coli and the remaining 31 isolates were C. jejuni. There were more C. coli, 98.1% (102/104), than C. jejuni, 28.1% (8/31), isolates resistant to both antimicrobials (p < 0.01). All other isolates (18.5%; 25/135) were susceptible to both antimicrobials (i.e., there were no isolates resistant to just one drug).
Pen-level prevalence of resistant Campylobacter spp. did not differ significantly (p = 0.61) between the treated and control pens for either drug tested (Fig. 3), and there was no evidence of a significant treatment by study week interaction (p = 0.85). There was, however, a significant week effect as the mean prevalence of resistant Campylobacter spp. isolates decreased over time for both treatment groups (p = 0.05).
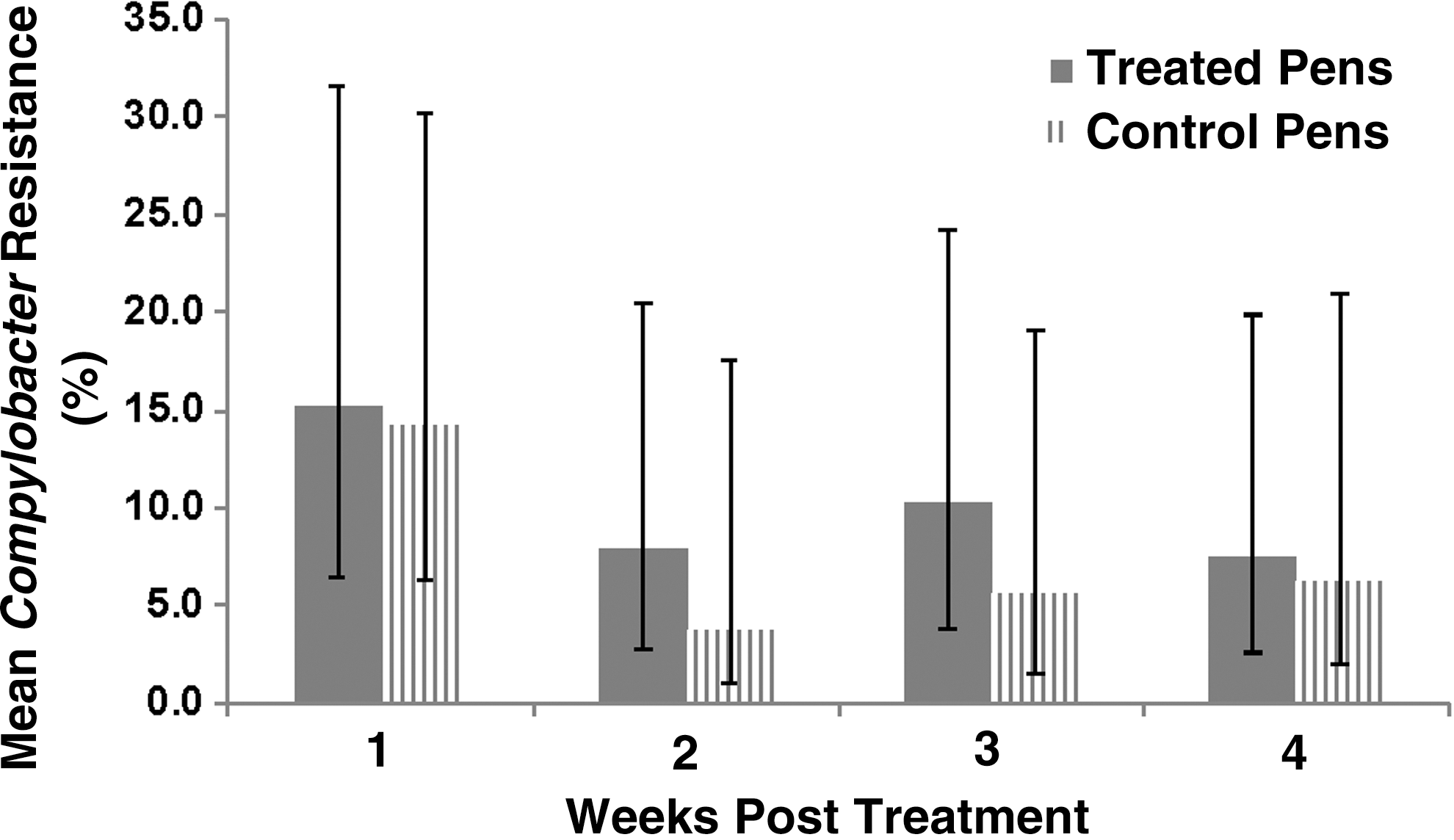
Model-adjusted mean pen-level percent of Campylobacter resistance to both nalidixic acid and ciprofloxacin for isolates recovered from feces of cattle in fluoroquinolone-treated and nontreated control groups. There was no significant interaction between time and treatment group, or a significant difference between treatment groups, but prevalence across both groups differed over time.
Discussion
This study demonstrated no evidence that enrofloxacin metaphylaxis impacted the prevalence of Salmonella sp. or Campylobacter spp. in the feces of feedlot cattle, nor did it impact the fecal prevalence of resistance to nalidixic acid or ciprofloxacin. Although not associated with treatment group, the overall prevalence of quinolone resistance in Campylobacter spp., particularly C. coli, was quite high (over 80% during the study period). In contrast, the overall prevalence of Salmonella sp. resistance to nalidixic acid or ciprofloxacin was very low (0% during the study period). Although the epidemiology and risk factors for antimicrobial resistance in Campylobacter spp. and Salmonella sp. in feedlot cattle must be further investigated, based on this study, enrofloxacin metaphylaxis was not found to be a significant driver of the prevalence and susceptibility of these organisms.
Considering the study season, location, and cattle type, the prevalence of Salmonella sp. and Campylobacter spp. in cattle during enrollment was consistent with other reports (Galland et al., 2000; Kunze et al., 2008; Sanad et al., 2011). Salmonella sp. prevalence did not differ significantly when comparing treated to nontreated pens of cattle. These findings were similar to results reported from a previous study where the number of fluoroquinolone treatments administered to cattle at five commercial feedlots was not associated with Salmonella prevalence (Smith et al., 2016). Salmonella prevalence did, however, differ across study weeks with prevalence decreasing over time. It is possible that being exposed to other cattle at the sale barn, the stress of transport, as well as acclimation to a new diet and environment had an impact on overall prevalence of Salmonella. Transportation of cattle can increase fecal shedding of Salmonella (Barham et al., 2002). Type of diet also has been associated with the Salmonella culture-positive status of cattle (Green et al., 2010). With prevalence in the current study being three to four times higher at study initiation than at the end of the study, it seems plausible that initial production or transportation stresses affected the prevalence of Salmonella sp. fecal shedding over time.
On the day of enrollment, prevalence of Salmonella sp. resistance was low, which was expected given that cattle had no previous exposure to a fluoroquinolone. The low prevalence of quinolone-resistant Salmonella sp. isolated from cattle is consistent with the literature (Beach et al., 2002; FDA, 2015). The single Salmonella enterica isolate that was resistant to nalidixic acid before treatment was also multidrug resistant (14 human antimicrobials). It is not unusual for Salmonella isolates from beef cattle to be either pan-susceptible or resistant to multiple antimicrobials (Dargatz et al., 2000; Khaitsa et al., 2007; Krueger et al., 2014).
All Salmonella isolates recovered posttreatment were susceptible to both nalidixic acid and ciprofloxacin, with MICs well below the breakpoint for both antimicrobials. These results are consistent with other research, where the isolation of quinolone-resistant Salmonella from beef cattle was either low or not detected (Dargatz et al., 2000; Beach et al., 2002; Kunze et al., 2008). Fluoroquinolone metaphylaxis did not significantly affect prevalence of resistant organisms isolated from cattle feces in this study, which is consistent with a previous observational study where fluoroquinolone use for BRD treatment within pens of cattle was not associated with the susceptibility of Salmonella sp. to human quinolones (Smith et al., 2016).
Data regarding the prevalence of Campylobacter spp. isolated from U.S. beef cattle are scarce. The current study showed that the prevalence of Campylobacter spp. across study weeks was highly variable for cattle in both the treated and nontreated control pens, and that metaphylaxis treatment with a fluoroquinolone was not associated with prevalence. As discussed for Salmonella sp., it is possible that acclimation to a new diet and environment may have contributed to the variability in Campylobacter spp. prevalence (Sproston et al., 2011).
C. jejuni is reported as the most prevalent Campylobacter species in cattle and the most common cause of human campylobacteriosis in the United States (Altekruse et al. 1999; Englen et al. 2005; FDA, 2014; Gaudreau et al. 2014). However, C. jejuni prevalence in this study was fairly low, accounting for less than 25% of the Campylobacter isolates recovered. C. coli was the most prevalent Campylobacter species isolated in this study. C. coli is typically isolated from swine and poultry and tends to have higher antimicrobial resistance prevalence when compared to C. jejuni (Moore et al. 2006). A similar trend was noted in the current study with significantly more C. coli isolates showing resistance to both nalidixic acid and ciprofloxacin when compared to C. jejuni. Campylobacter mutants found to be highly resistant to quinolone antimicrobials typically have a single-point mutation in the gyrA gene, which is part of the quinolone drug binding site (Luo et al., 2003). This single-step mutation keeps quinolone antimicrobials from binding, causing clinically relevant levels of resistance to quinolone antimicrobials (Wang et al., 1993).
Studies of the effects of fluoroquinolone treatment on the prevalence of fluoroquinolone-resistant Campylobacter spp. in swine and poultry have shown that resistance prevalence increases rapidly after treatment and persists for several days in both the animal and the environment (McDermott et al., 2002; Usui et al., 2014). However, results from the current study showed that the mean Campylobacter spp. resistance prevalence was not significantly impacted by fluoroquinolone use, and declined over time for both treatment groups to less than 10% by the end of the 4-week study period. Assuming this decline reflects what is commonly found in commercial feedlots after the Baytril® 28-day withdrawal time has been met, cattle should have minimal resistance prevalence at harvest. With the National Antimicrobial Resistance Monitoring System (NARMS) no longer reporting Campylobacter spp. due to low prevalence of this organism in ground beef, this assumption is plausible (FDA, 2015).
Prevalence of Campylobacter spp. resistance at baseline (before drug administration) was rather high with >60% of the isolates resistant to at least one of the antimicrobials. These results were unexpected, since cattle had no documented history of being administered a fluoroquinolone when cattle were purchased for this study. However, a similar finding was noted in a longitudinal study of antimicrobial-resistant Campylobacter prevalence in conventional and antibiotic-free swine (Quintana-Hayashi and Thakur, 2012a). Authors reported a high prevalence of multidrug-resistant (MDR) Campylobacter obtained from antibiotic-free swine and their environment, with the majority of isolates identified as C. coli. In a follow-up study, multilocus sequence typing on the MDR C. coli isolates demonstrated that pigs did not carry MDR isolates at the farm but acquired the resistant strains at the slaughter plant (Quintana-Hayashi and Thakur, 2012b).
It is possible that cattle in the current study acquired resistant isolates either at the sale barn or during transit to the study site. However, the time of acquisition of the resistant organisms could not be determined in the present study as the cattle were not sampled before arrival to the study site. As there were no data on resistance prevalence for cattle before study initiation, it was not accounted for in the study design, and the resulting randomization process unfortunately resulted in a difference in the prevalence of animals shedding resistant organisms between treated and control pens. While the statistical analysis of treatment effects accounted for the baseline prevalence, the large number of cattle shedding resistant Campylobacter spp. before enrollment may have influenced the overall Campylobacter prevalence or resistance prevalence throughout the study. As noted by Zhang et al. (2006), resistant Campylobacter isolates can have a fitness advantage compared with susceptible isolates, even in the absence of antimicrobial selection pressure. This may have influenced the overall resistance prevalence during the 4-week study period, even though resistance did not differ significantly between treatment groups (Fig. 3).
No significant differences were observed in the pen-level fecal prevalence of quinolone-resistant Campylobacter between cattle administered fluoroquinolone metaphylaxis compared with nontreated cattle. Although MIC values were generally high throughout the study, they were consistently high for isolates recovered from both treatment groups. Adding monensin to the feed of both treatment groups may have influenced the overall resistance prevalence of Campylobacter spp. However, ionophores have a different mode of action than fluoroquinolones, and bacteria typically adapt to ionophores rather than mutate or acquire resistance genes (Russell and Houlihan, 2003). With C. coli being the most prevalent species isolated, an overall high prevalence of resistance to quinolones could be expected (Moore et al., 2006; Gaudreau et al., 2014).
In conclusion, our results indicated that metaphylaxis administration of the fluoroquinolone, enrofloxacin, to beef cattle for the control of BRD did not significantly affect fecal prevalence of Salmonella sp. and Campylobacter spp. or fecal prevalence of resistance. A relatively large proportion of the cattle enrolled in this study were shedding Salmonella sp. and Campylobacter spp. in their feces before study initiation, with a high percentage of the Campylobacter spp. resistant to quinolones even though to our knowledge cattle were not previously exposed to a fluoroquinolone. Despite cattle harboring quinolone-resistant Campylobacter spp. before enrollment, metaphylaxis use of a fluoroquinolone did not increase the fecal prevalence of resistance after administration and, in fact, the prevalence of resistance declined (in both treatment groups) throughout the study period.
Footnotes
Acknowledgments
This research was supported by Bayer Animal Health and the Department of Diagnostic Medicine/Pathobiology, College of Veterinary Medicine, Kansas State University. Investigators thank Dr. Keith DeDonder, Dr. Kelly Lechtenberg, and the staff at Veterinary Biomedical Research Center, Inc., as well as the staff and students at the Kansas State University, College of Veterinary Medicine, Preharvest Food Safety Laboratory for their time and contribution to this study. This is contribution number 17-186-J from the Kansas Agricultural Experiment Station.
Disclosure Statement
The first author (A. Smith) is a graduate student of Kansas State University and an employee of Bayer Animal Health who produce a commercially licensed fluoroquinolone for the treatment and control of BRD.