
Abstract
This study focused on advancing a rapid turbidimetric bioassay to screen antimicrobials using specific cocktails of targeted foodborne bacterial pathogens. Specifically, to show the relevance of this rapid screening tool, the antimicrobial potential of generally recognized as safe calcium diacetate (DAX) and blends with cranberry (NC) and oregano (OX) natural extracts was evaluated. Furthermore, the same extracts were evaluated against beneficial lactic acid bacteria. The targeted foodborne pathogens evaluated were Escherichia coli O157:H7, Salmonella spp., Listeria monocytogenes, and Staphylococcus aureus using optimized initial cocktails (∼108 colony-forming unit/mL) containing strains isolated from human food outbreaks. Of all extracts evaluated, 0.51% (w/v) DAX in ethanol was the most effective against all four pathogens. However, DAX when reduced to 0.26% and with added blends from ethanol extractions consisting of DAX:OX (3:1), slightly outperformed or was equal to same levels of DAX alone. Subculture of wells in which no growth occurred after 1 week indicated that all water and ethanol extracts were bacteriostatic against the pathogens tested. All the targeted antimicrobials had no effect on the probiotic organism Lactobacillus plantarum. The use of such rapid screening methods combined with the use of multistrain cocktails of targeted foodborne pathogens from outbreaks will allow rapid large-scale screening of antimicrobials and enable further detailed studies in targeted model food systems.
Introduction
T
Past research by others has focused on the antimicrobial properties of the sodium salt (sodium diacetate) in various food applications (Schlyter et al., 1993; Degnan et al., 1994; Mbandi and Shelef, 2001, 2002; Stekelenburg and Kant-Muermans, 2001; Stekelenburg 2003; Barmpalia et al., 2005), and blends of potassium lactate and sodium diacetate are widely used in Ready-to-Eat meats to control Listeria monocytogenes. Although Shelef and Addala (1994) demonstrated the antimicrobial properties of sodium diacetate toward L. monocytogenes, Escherichia coli, Pseudomonas fluorescens, Salmonella Enteritidis, and Shewanella putrefaciens, it showed no inhibitory effect toward Lactobacillus fermentans, Staphylococcus aureus, Pseudomonas fragi, Yersinia enterocolitica, or Enterococcus faecalis. However, extracts from botanical-based food materials are GRAS and have excellent potential as sources of natural antimicrobials in combating foodborne pathogens.
A key challenge, however, is the identification of effective combination of DAX/botanical blends with an antimicrobial potential capable of inhibiting foodborne pathogens without affecting beneficial probiotic organisms. Previous studies from our laboratory and others have demonstrated the inhibitory potential of a wide range of food-grade botanicals against foodborne pathogens (Puupponen-Pimia et al., 2001; Seaberg et al., 2003; Lin et al., 2004, 2005; Vattem et al., 2004a, b; McCue et al., 2005; Shetty and Lin, 2005; Kwon et al., 2007; Apostolidis, et al., 2008). The inhibitory activity of these botanicals was further enhanced with the inclusion of natural organic acids (Shetty and Lin, 2005; Apostolidis et al., 2008). Organic acids destabilize the membranes and increase permeability to botanical bioactives that could lead to further inhibition through membrane disruption. Therefore, the specific goal of this study was to explore the use of a high-throughput protocol to evaluate the antimicrobial properties of DAX/botanical blends as an initial rapid screening tool targeting a cocktail of foodborne pathogens from recent outbreaks and also the effect on beneficial lactic acid bacteria (LAB). Such rapid screening approach using turbidimetric methods provides the basis to target and subsequently screen narrow potential choices for further detailed study for wider mechanistic-based food safety-related applications.
Materials and Methods
Strains and media
Organisms used in this study were of various strains: Escherichia coli O157:H7, Salmonella spp., L. monocytogenes, S. aureus, and Lactobacillus plantarum (Table 1). All nutrients and media supplies were purchased from ThermoFisher Scientific (Waltham, MA) and Becton, Dickinson and Company (Franklin Lakes, NJ).
American Type Culture Collection, Manassas, VA.
Dr. C. Kaspar, University of Wisconsin–Madison, Madison, WI.
Microbial Food Safety Lab, Iowa State University, Ames, IA.
Salmonella Genetic Stock Centre, University of Calgary, Calgary, AB.
Wisconsin State Laboratory of Hygiene, Madison, WI.
ILSI NA Listeria monocytogenes Strain Collection, Cornell University, Ithaca, NY.
Escherichia coli O157:H7 was cultured at 35°C in tryptic soy broth plus 0.6% (w/v) yeast extract (TSBYE). L. monocytogenes was cultured at 30°C in brain-heart infusion (BHI). S. aureus and Salmonella spp. were cultured in TSB at 30°C and 35°C, respectively. L. plantarum was grown in de Man, Rogosa, Sharpe (MRS) broth at 35°C. Escherichia coli O157:H7, Salmonella, and S. aureus cell counts were performed using TSAYE plates; L. monocytogenes and L. plantarum cell counts were performed using BHI and MRS agars, respectively. Stock cultures of all organisms were maintained on refrigerated agar slants and at −80°C in 20% (v/v) glycerol.
With the exception of L. plantarum, a four- to six-strain mixture (“cocktail”) of each test organism was used for all experiments. Cocktails were prepared by aseptically combining 5 mL of overnight culture of each component strain (106–109 colony-forming unit [CFU]/mL) in a sterile 50-mL polypropylene centrifuge tube, and the multistrain pellet harvested by centrifugation (10,000 × g, 10 min, 4°C) in a Sorvall Super T-21 centrifuge with an SL-50T rotor. The supernatants were discarded and the pelleted cells were resuspended and washed in 25 mL sterile 0.85% (w/v) saline. The washed cells were harvested as previously described and the supernatants discarded. The cells were resuspended in 25 mL sterile 0.85% (w/v) saline and then diluted with saline to produce suspensions with ∼108 CFU/mL. Ten microliters of the final dilutions were aseptically pipetted into individual wells of sterile Bioscreen plates and evaluated for antimicrobial activity using a Bioscreen C Microbiology Reader (see details for Bioscreen operation hereunder).
Extract preparation
Natural plant extracts blended with 12 different organic acids (1:1) were provided by Amherst BioInnovations (Amherst, MA) and formulated as below after sourcing oregano (OX; Barringtion Nutritionals, Harrison, NY) and cranberry (NC; Decas Cranberry Products, Carver, MA). Powdered extracts were weighed directly into sterile screw-capped tubes using sterile spatulas. Two portions of each extract were prepared. One portion was added to sterile reverse osmosis (RO) water and the other to sterile RO water containing 20%, 30%, or 40% (v/v) ethanol. These preparations were vortexed thoroughly. Extracts that did not readily dissolve were heated for ∼30 min in a water bath at 55–70°C with further vortexing to improve dissolution. The cooled extracts were spun at 4°C on a Fisher Scientific Centrific centrifuge at 1380 × g for 30 min, after which the clarified supernatants were aseptically transferred to sterile screw-capped tubes. Initial concentrations of extracts were 1.5 or 3% (w/v). Dilutions and blends of the extracts were stored at 4–8°C until needed.
Phenolic acid analysis and antimicrobial formulations
Total phenolic content was assayed as described by Kwon et al., (2007) using gallic acid as a standard. The phenolic extracts from OX and NC were prepared as water-extracted and spray-dried products. The phenolic water or ethanol extracts were blended separately in 1:1 weight ratios with standard commercial DAX and used in these studies. The DAX blends with 50% oregano extracts (OXDA) and 50% cranberry extracts (NCDA) proved superior in preliminary trials and were therefore examined further in this study.
Bioscreen measurements
The antimicrobial activity of each extract was tested using a Microbiology Reader Bioscreen C (Growth Curves USA, Piscataway, NJ). Evaluations were performed as follows: 10 μL (∼106 CFU) of diluted washed cell suspensions was added to Bioscreen wells and the inocula dried at room temperature in a biological safety hood for 60–90 min. After this time, 60 μL of plant extract (1.8 mg/60 μL) was aseptically added to microtiter wells containing the air-dried microbial cocktails. After a 1-min exposure to the extract, 280 μL of sterile 1.25x culture broth was added to the wells (final volume of 350 μL and 0.51% [w/v] plant extract). Wells treated with 60 μL of sterile water or sterile 20% (v/v) ethanol served as positive controls; uninoculated wells with extracts plus sterile media were used to test the sterility of solutions and obtain background absorbance.
The plates were incubated in the Bioscreen at appropriate temperatures for 3–7 days with continuous low-amplitude shaking (0.6 mm left and then right). Absorbance readings at 600 nm (A600) were recorded hourly. Extracts were evaluated in replicates of four to five microtiter wells per run for each incubation. Replicate runs of at least two were performed for each extract and pathogen. At the completion of the experiments, Bioscreen data were downloaded into Excel for analysis. Strength of inhibition resulting from exposure to extracts was scored by visual inspection of bacterial culture turbidity as follows: not turbid = 4 (Abs600 average 0.11); slightly turbid = 3 (Abs600 average 0.64); moderately turbid = 2 (Abs600 average 1.30); turbid = 1 (Abs600 average 1.97); and highly turbid = 0 (Abs600 y-intercept of the line was 2.57). In this scoring process, 4 = highest inhibition strength and 0 = no inhibition strength.
Determination of the pH effect on growth
The effect of pH was determined by culturing E. coli in TSBYE adjusted to various pHs with H3PO4 (pH 4.0–7.0). The pHs of DAX-free media were adjusted with H3PO4 and then autoclaved. Wells were inoculated with a washed cell suspension of E. coli (9 × 105 CFU) followed by addition of specific pH-buffered media. Bioscreen incubation was with shaking for 48 h at 35°C and absorbance recorded hourly at 600 nm. Final concentration of DAX in each well was 0.51% (w/v). Each data point represents the average of four or five microtiter wells. Furthermore, the titration curve for 0.51% (w/v) DAX media was determined by halving of the solution for titration with 0.1 N HCl, and the other half for titration with 0.1 N NaOH. pH values were recorded on a precalibrated Accumet AB15 pH meter.
Viability testing
To characterize extracts as bacteriostatic or bactericidal, cell viability was determined for Bioscreen wells in which growth did not occur over the course of each experiment. Briefly, 10–50 μL aliquots were aseptically transferred from these wells to tubes containing 3–10 mL of an appropriate sterile culture broth, followed by incubation at a suitable temperature for 24–48 h. If growth was observed in the culture tube (highly turbid), the extract was considered to be bacteriostatic, while no growth was scored as bactericidal.
Results
Initially, 10 different extract combinations with organic acids were tested for biological activity against Escherichia coli O157:H7, which proved to be the most resistant. From this preliminary study, two extract blends with DAX (DAX as control, OXDA, and NCDA) were selected for further characterization. Figure 1 shows that growth of E. coli was substantially inhibited by aqueous and ethanolic 3% (w/v) DAX compared to growth in extract-free media. Inhibition was persistent with DAX prepared using 20% (v/v) ethanol as solvent, although the differences between water and 20% (v/v) ethanol were not apparent until after 61 h of incubation. E. coli was also inhibited by the extract blends NCDA and OXDA; however, the extent of inhibition was less than for DAX alone (Table 2). With the former extracts, inhibition was markedly greater in the presence of ethanol (3.43% [v/v] ethanol per well). These data suggest that ethanol may either enhance solubility of these extracts or the presence of ethanol acts to facilitate extract passage across the cell membrane. Postexperiment plating confirmed that wells with little to no visible growth contained viable cells, indicating the bacteriostatic nature of these treatments. In addition, the inhibitory actions of these extracts were not affected by inoculum size, as determined using E. coli inocula with cell densities of 1.5 × 104, 105, and 106 CFU/well (data not shown).
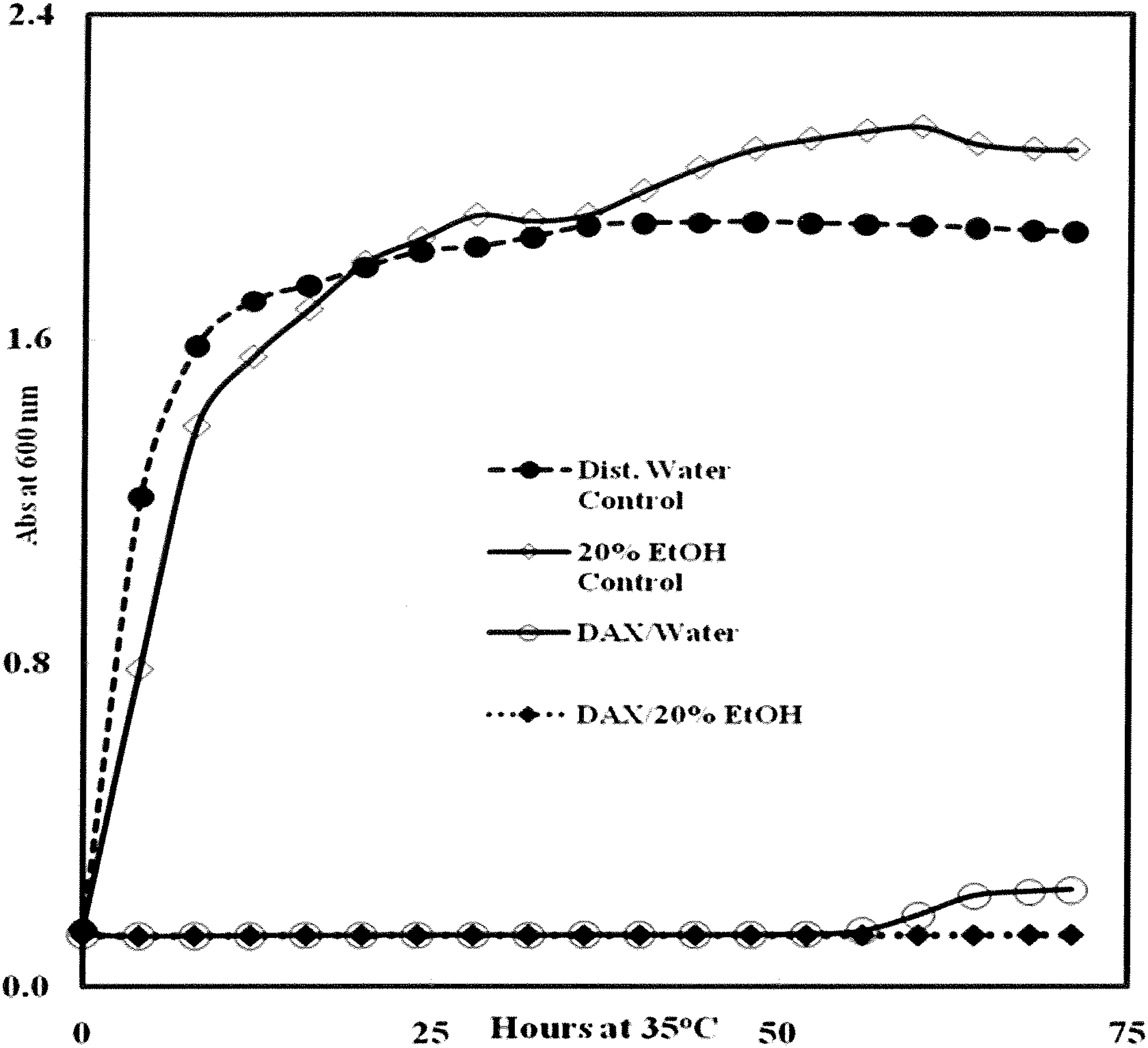
Effect of 3% (w/v) calcium diacetate (DAX) prepared in water and 20% (v/v) EtOH on growth of Escherichia coli O157:H7 cocktail (Table 1) in TSBYE. Wells were inoculated with 10 μL (2.8 × 106 CFU) of a washed cell suspension of E. coli in saline, followed by addition of 60 μL of 3% (w/v) DAX and 280 μL of sterile TSBYE. Final concentrations of DAX and EtOH in wells were 0.51% (w/v) and 3.4% (v/v), respectively. Each data point represents the average of four or five microtiter wells. No inhibition was observed in uninoculated control wells (data not shown). CFU, colony-forming unit; TSBYE, tryptic soy broth plus 0.6% (w/v) yeast extract.
DAX is 100% (w/v) calcium diacetate; NCDA blend is 50% (w/v) cranberry extract and 50% (w/v) calcium diacetate; OXDA blend is 50% (w/v) oregano extract and 50% (w/v) calcium diacetate.
RO water was used; final ethanol concentration in wells was 3.42% (v/v).
Starting pH was 7.1 for TSBYE broth and CFU per well was 2.8 × 106. Each extract and bacterial cocktail were evaluated in four to five microtiter wells per trial.
Inhibition strength: 0 = none; 1 = slight; 2 = moderate; 3 = strong; and 4 = very strong.
CFU, colony-forming unit; RO, reverse osmosis; TSBYE, tryptic soy broth plus 0.6% (w/v) yeast extract.
Table 2 shows that the extracts lowered the pH of TSBYE from ∼7.1 to <6.0, suggesting that reduced pH was a factor of treatment efficacy. Thus, E. coli in TSBYE was prepared in 125 mM phosphate buffer, pH 7.1. The experiment was also modified by preparing the extracts in 30% (v/v) ethanol (5.14% [v/v] ethanol per well) to ensure all plant extractable phenolics were present. Table 3 shows that all three extracts had a moderate-to-strong inhibitory activity against E. coli at pH 6.5–6.8. DAX was still inhibitory at these higher pH values, although it was less inhibitory than NCDA and OXDA (1:1) blends under the same conditions. Importantly, all ethanolic extracts were more inhibitory than controls with ethanol alone (5.14% [v/v] ethanol per well).
TSBYE prepared in 125 mM phosphate buffer (pH 7.1); CFU per well was 3.2 × 106. Each extract and bacterial cocktail were evaluated in four to five microtiter wells per trial.
DAX is 100% (w/v) calcium diacetate; NCDA blend is 50% (w/v) cranberry extract and 50% (w/v) calcium diacetate; OXDA blend is 50% (w/v) oregano extract and 50% (w/v) calcium diacetate.
RO water was used and final ethanol concentration in wells was 5.14% (v/v).
Inhibition strength: 0 = none; 1 = slight; 2 = moderate; 3 = strong; and 4 = very strong.
CFU, colony-forming unit; RO, reverse osmosis; TSBYE, tryptic soy broth plus 0.6% (w/v) yeast extract.
The effect of pH was examined further by culturing E. coli in DAX-free TSBYE adjusted to various pHs with H3PO4 (Fig. 2). E. coli failed to grow at pH 4; however, growth improved with increasing pH, to a maximum in the pH range of 5.6–7.0. Similar patterns (data not shown) were obtained with Salmonella spp., S. aureus and L. monocytogenes. In DAX (3% [w/v] DAX in water)-containing TSBYE adjusted to pH 4.8 and 5.1, complete inhibition of E. coli was observed (Fig. 3), whereas moderate inhibition was observed in DAX-containing media buffered with phosphate at pH 5.8. E. coli cells grew in the pH range 4.5–6.5; their failure to grow in this range in the presence of DAX shows that the inhibition was due to DAX (0.51% [w/v] DAX per well), rather than solely a pH effect.
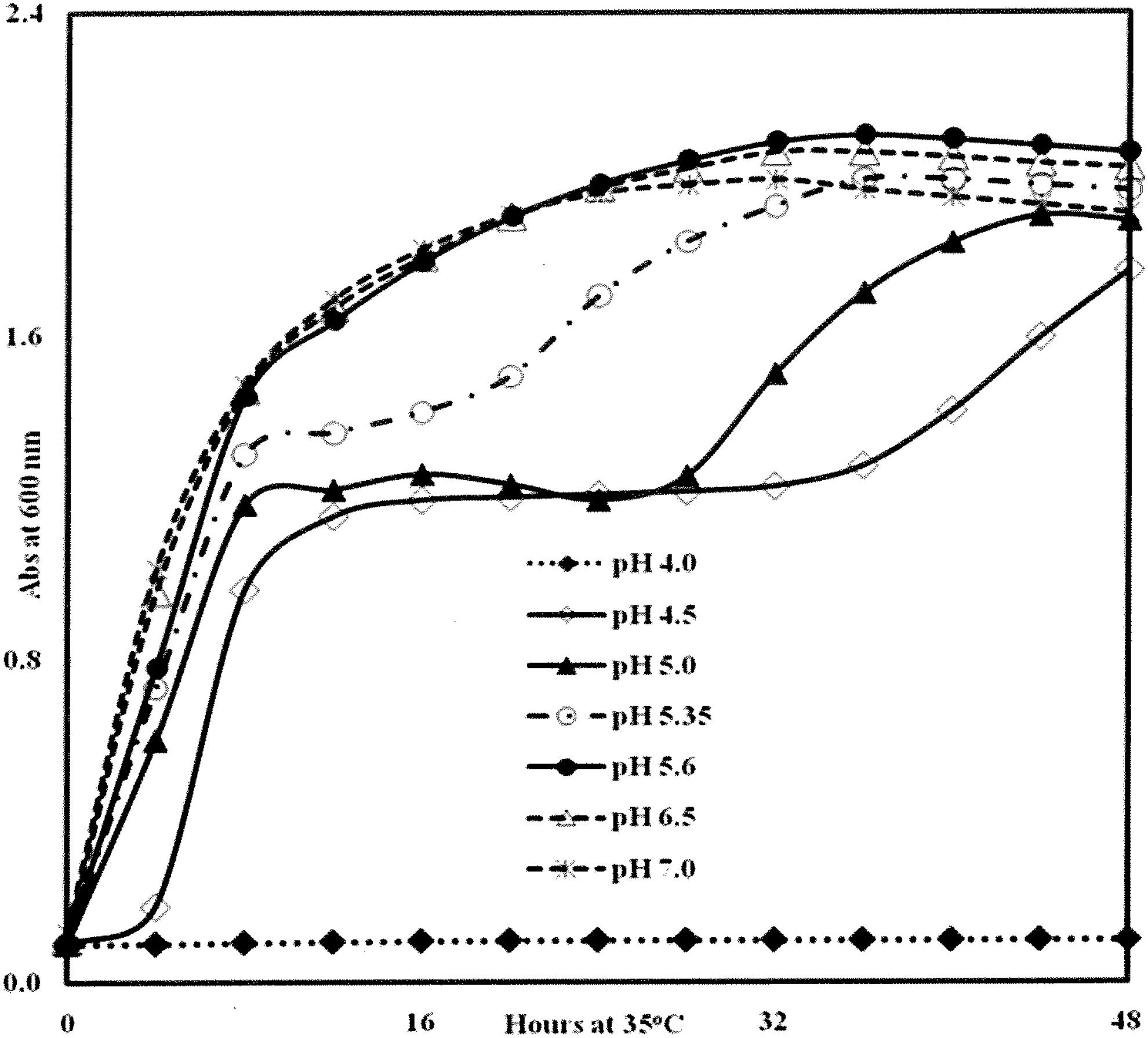
Effect of pH on growth of Escherichia coli O157:H7 cocktail in TSBYE medium at 35°C with shaking. Cells were prepared as described in Materials and Methods. Media pH's were adjusted with H3PO4 and then autoclaved. Wells were inoculated with a washed cell suspension of E. coli (9 × 105 CFU) followed by addition of media. Each data point represents the average of four or five microtiter wells. CFU, colony-forming unit; TSBYE, tryptic soy broth plus 0.6% (w/v) yeast extract.
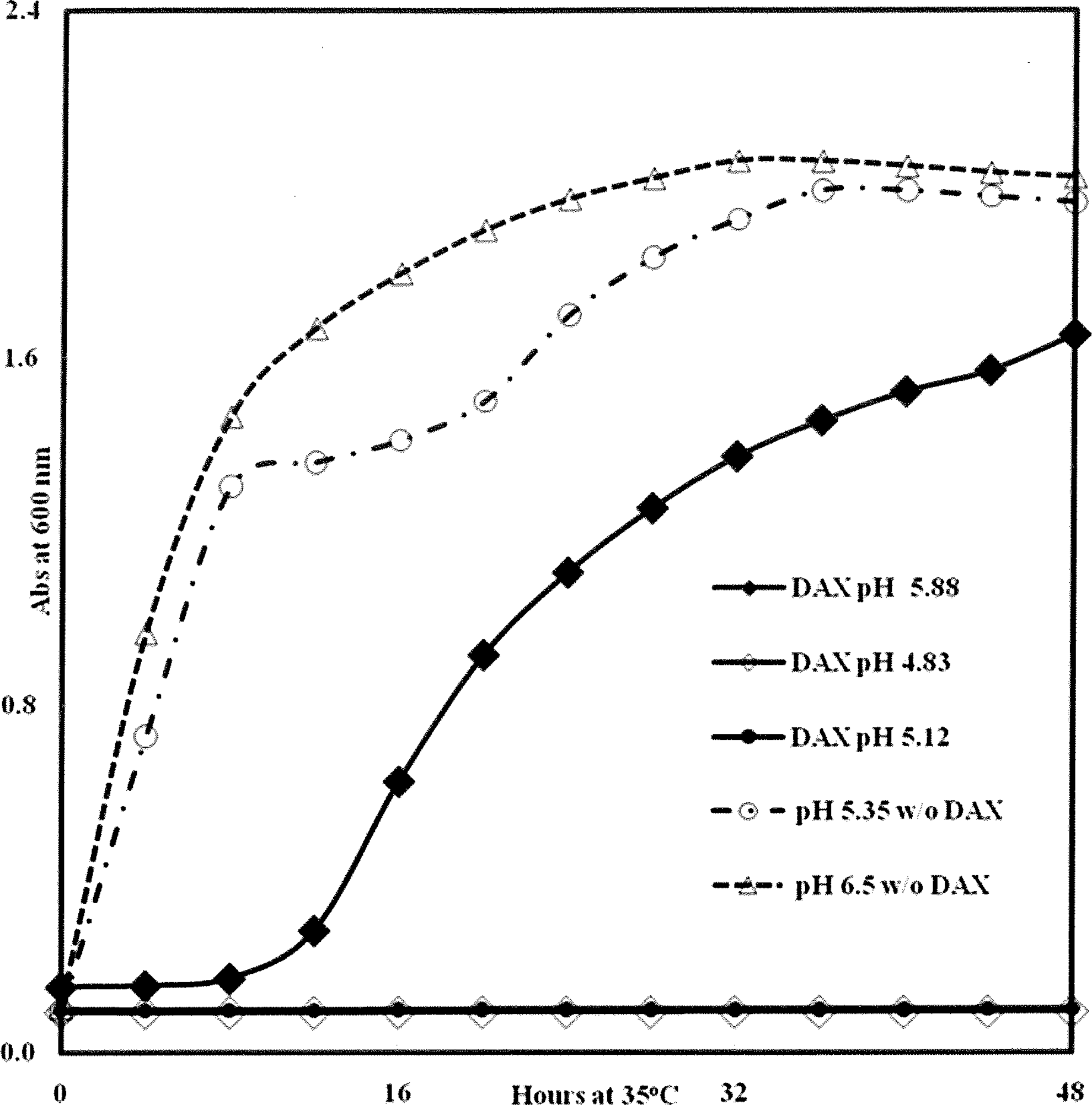
Effect of calcium diacetate (3% [w/v] DAX in water) on growth of Escherichia coli O157:H7 cocktail in TSBYE at various pHs. Cells were prepared as described in Materials and Methods. The pHs of DAX-free media were adjusted with H3PO4 and then autoclaved. Wells were inoculated with a washed cell suspension of E. coli (9 × 105 CFU) followed by addition of media. Incubation was with shaking for 48 h at 35°C. Final concentrations of DAX in each wells was 0.51% (w/v). Each data point represents the average of four or five microtiter wells. CFU, colony-forming unit; TSBYE, tryptic soy broth plus 0.6% (w/v) yeast extract.
Table 4 shows the effect of the extracts on L. plantarum and four common food pathogens, Escherichia coli O157:H7, Salmonella spp., L. monocytogenes, and S. aureus. In these trials, DAX was tested alone and in combination with OX and NC. DAX alone at 0.51% [w/v] DAX per well consistently produced the highest levels of inhibition for all pathogens tested. With all extracts, inhibition was higher when 20% (v/v) ethanol was used as the solvent versus water. Salmonella was the most sensitive organism, followed by L. monocytogenes, S. aureus, and E. coli. In contrast, none of the extracts had any effect on L. plantarum. At the completion of the experiment, wells with little to no growth were found to contain viable cells, demonstrating that the extracts were bacteriostatic, and not bactericidal, toward the pathogens.
Each trial represents an average of two completely replicated studies. Each extract and microbe were evaluated in four to five microtiter wells per trial.
Blends initially contained 1.5% or 3% (w/v) each extract. DAX is 100% (w/v) calcium diacetate; DAX/OX blend is 75% (w/v) calcium diacetate and 25% (w/v) oregano extracts; DAX/NC/OX blend is 50% (w/v) calcium diacetate, 25% (w/v) cranberry extract, and 25% (w/v) oregano extract.
Inhibition strength: 0 = none; 1 = slight; 2 = moderate; 3 = strong; and 4 = very strong.
Final ethanol concentration in wells was 3.42% (v/v).
Analyses showed that OXDA had the highest level of phenolic compounds and NCDA had moderate phenolic content (Table 5). Titration of a 1% (w/v) aqueous DAX solution yielded a pK at ∼4.7 (acetic acid has a pKa of 4.74) (Fig. 4). The pKs for phenolic compounds have a range of 4.2–4.6 (Wen et al., 2002). With all the extracts, the inclusion of ethanol in the solvent seems to have little effect on phenolic solubility. Thus, an improved antimicrobial activity was the result of ethanol presence.
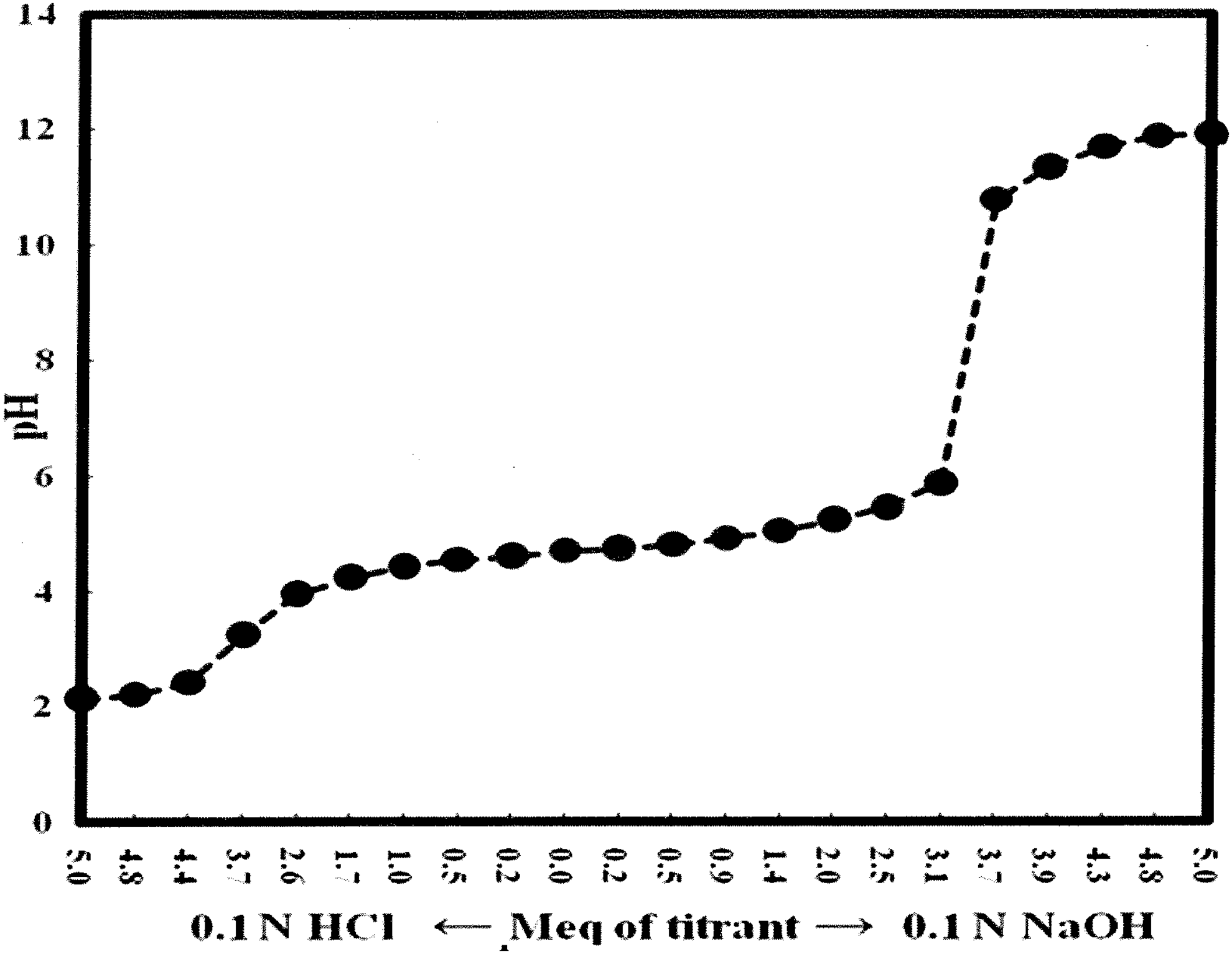
Titration curve of calcium diacetate (DAX). A 1% (w/v) solution of DAX was prepared in distilled water. Half of the solution was titrated with 0.1 N HCl, and the other half was titrated with 0.1 N NaOH. pH values were recorded on a precalibrated Accumet AB15 pH meter.
DAX is 100% (w/v) calcium diacetate; NCDA blend is 50% (w/v) cranberry extract and 50% (w/v) calcium diacetate; OXDA blend is 50% (w/v) oregano extract and 50% (w/v) calcium diacetate.
Total phenolics were assayed as described by Kwon et al. (2007), using gallic acid as the standard. Cranberry and oregano had minimal total phenolic contents of 10 and 40 mg/g, respectively.
DAX at 0.51% [w/v] per well exhibited the highest antimicrobial activity against the four pathogens tested (Table 4). OX demonstrated the highest measurable level of phenolics (Table 5). Extracts with 75% DAX plus 25% OX in 20% (w/v) ethanol (0.26% [w/v] per well) were similar or slightly improved against cocktails of E. coli and L. monocytogenes; for Salmonella spp., 100% DAX and 75% DAX with 25% OX gave similar results (Table 4).
Of all the antimicrobial substances tested, 3% (w/v) DAX (0.51% [w/v] per well) in ethanol or water was the most effective against all four pathogens (Table 4; Fig. 5). The effect was bacteriostatic. Although the extent of pathogen inhibition varied among trials with pathogen cocktails, the effect was reproducible. However, the use of cocktails also allows greater confidence in and applicability of results, as this approach compensates for strain-to-strain variation. For 3% (w/v) DAX trials lasting 7 days, pathogen growth was prevented for 48–60 h, with Escherichia coli O157:H7 recovering the quickest. (Fig. 5). At a minimum, regrowth would occur if only one of the 4–6 strains was resistant to a given treatment. Interestingly, ethanol extracts of 3% (w/v) DAX prevented growth of both Salmonella spp. and S. aureus during the full 7-day period, whereas water extracts did not (Fig. 5). The 20% (v/v) ethanol controls had no effect on any of the pathogens tested, whereas throughout the study, the ethanol extract blends outperformed those with water, which suggests that the presence of 3.4% (v/v) ethanol in each well improved the bacteriostatic potential for DAX to each pathogen. This difference deserves further study.
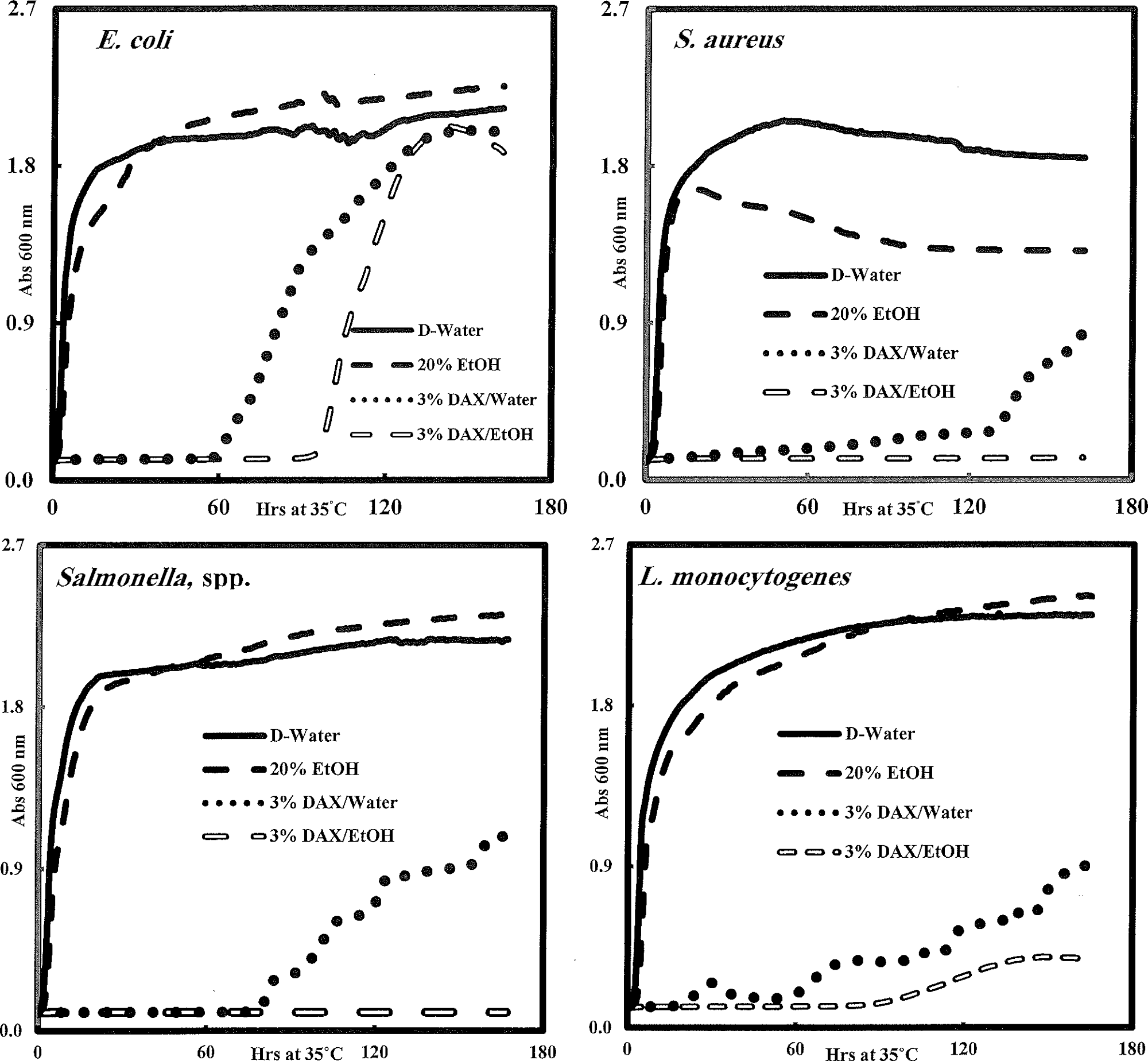
Effect of 3% (w/v) calcium diacetate (DAX) on long-term (>7 days) incubation of Escherichia coli O157:H7, S. aureus, Salmonella spp., and L. monocytogenes cocktails. The procedures for extract preparation and culturing cells were as described in Materials and Methods. Wells were inoculated with 10 μL of washed cell suspensions of the indicated organisms, followed by addition of 60 μL of extract and 280 μL of sterile TSBYE. Final concentrations of DAX and EtOH in wells were 0.51% (w/v) and 3.4% (v/v), respectively. Incubation was with shaking at 35°C. Each data point represents the average of four or five microtiter wells. No inhibition was observed in uninoculated control wells (data not shown). TSBYE, tryptic soy broth plus 0.6% (w/v) yeast extract.
Discussion
We observed some difference and similarities between the sodium and calcium diacetates. Shelef and Addala (1994) reported that sodium diacetate had no effect on S. aureus, but did inhibit Salmonella Enteritidis, while in this study, DAX did inhibit S. aureus and Salmonella spp. The lack of sensitivity of L. plantarum to DAX (this study) confirms the findings of Stekelenburg and Kant-Muermans (2001) with Lactobacillus curvatus and those of Shelef and Addala (1994) with Lactobacillus fermentis, demonstrating that sodium diacetate salts have no effect on lactobacilli.
Addition of extracts to TSBYE lowered the pH of this medium (Table 2) and bacterial inhibition was found to be greater at lower pH, as observed by others (Herald and Davidson, 1983; Kouassi and Shelef, 1998; Wen et al., 2002). Degnan et al. (1994) also demonstrated that sodium diacetate rinse (2 M; 28.4% [w/v]) of crab meat did reduce the pH to 5.3 (close to DAX pH, Table 2) and inhibit L. monocytogenes. All DAX blends (NCDA and OXDA) demonstrated a slight pH increase for water and 20% (v/v) ethanol extracts (pH 0.5–0.6 increase, Table 2) compared to DAX alone, while each 20% (v/v) ethanol blend (NCDA and OXDA) demonstrated a slight reduction in Escherichia coli O157:H7 inhibition (inhibition strength of four for DAX and three for blends).
Furthermore, when pH was buffered (Table 3), the antimicrobial activity for both blends (NDCA and OXDA) slightly outperformed DAX alone in ethanol. This observation is consistent with the presence of phenolic and/or other weak acids in the extracts, including benzoic acid, a major constituent of cranberries (Jay, 2005; Neto and Vison, 2011). At lower pH values, a higher percentage of these compounds are undissociated, and as uncharged molecules, can more easily cross the cell membrane to the cell interior (Wen et al., 2002). For example, the pKa of benzoate is 4.20. At pH 4.0, 60% is present in the antimicrobially effective undissociated form, whereas at pH 6.0, only 1.5% is undissociated (Jay, 2005). Kouassi and Shelef (1998) showed that the antilisterial activity of cinnamic acid was eliminated at pHs >6. In our case, the antimicrobial activity of the extracts was influenced by pH; however the inhibition was not prevented at neutral pH in the presence of 5.14% (v/v) ethanol per well (Table 3). Enhanced antimicrobial effects in the presence of ethanol suggest that ethanol facilitated antimicrobial uptake through permeabilization of the cell membrane. As a control, we adjusted TSBYE to pH levels corresponding to those measured after extract addition. All the pathogens tested grew in pH-adjusted extract-free TSBYE, demonstrating that inhibition by extracts did not stem from simple pH effects.
The antimicrobial activity of many plant extracts is generally attributed to the phenylpropanoid pathway-derived phenolic acids (e.g., p-coumaric acid, ferulic acid, caffeic acid, cinnamic acid, and potentially their derivatives) (Herald and Davidson, 1983; Kouassi and Shelef, 1998; Wen et al., 2002; Shetty and Wahlqvist, 2004; Shetty and Lin, 2005; Proestos et al., 2006). In this study, DAX extract combinations with OX and NC enriched in caffeic acid, and coumaric acid in case of OX (Kwon et al., 2007) and hydroxybenzoic acid in case of additional NC (Zheng and Shetty, 2000) were used (Table 5). For S. aureus, 75% DAX with 25% OX slightly outperformed 100% DAX at 0.26% extracts per well (Table 4), which correlates with Kwon et al. (2007) who showed that S. aureus was inhibited by gallic acid, caffeic acids, and phenol-containing plant extracts and inhibition by the latter was correlated with phenolic content.
The ability of some LAB to survive in the presence of phenolic compounds has been observed previously. For example, Tabasco et al. (2011) screened various LAB for suppression or growth in the presence of grape seed extract (GSE) and found that while some LAB were extremely sensitive to GSE, others were able to grow in the presence of this polyphenol-rich phytoextract. Interestingly, L. plantarum was not only able to survive exposure to GSE but also able to actively metabolize various flavan-3-ols in a separate screen by esterase, decarboxylase, and dehydrogenase activities (Tabasco et al., 2011).
In addition, this study showed the ability of various DAX/plant extract preparations to selectively inhibit key foodborne pathogens without affecting L. plantarum, a model probiotic organism. Extracts having antimicrobial effects resulting primarily from weak acids would be appropriate primary preservatives or hurdles in acidic foods, such as juices, soft drinks, condiments, and salad dressings (Jay, 2005). Alternatively, select extracts may be useful in the production of probiotic cheeses such as a recently described probiotic Edam-type cheese containing L. plantarum and having an initial curd pH ranging from 4.5 to 4.9 (Songisepp et al., 2012).
Conclusions
In summary, the rationale for this research was to advance a rapid protocol for screening the antimicrobial properties of natural extract synergies with DAX using a turbidimetric method, which employed a cocktail of single genera of known food pathogens. When required, once inhibition is observed for cocktails, evaluations can be rapidly performed with individual strains, including details for CFU/mL. Furthermore, the rapid determination for bacteriostatic or bactericidal activity using this turbidimetric method was also demonstrated. This rapid screening method will justify further studies in food systems with DAX alone (0.51%) and in combination with OX (0.26%), which showed initial positive results. Our study also provided initial insights on how ethanol can facilitate the selective inhibition by DAX with natural extract of important foodborne pathogens. Furthermore, it was imperative and important to show that such inhibition was absent in L. plantarum. Therefore, this research provides a valuable and rapid tool for screening large numbers of compounds against pathogen cocktails for the strategic and systematic formulation of novel hurdle technologies in support of food safety efforts.
Footnotes
Acknowledgments
This research was supported by the State of Iowa Bioscience Alliance, Iowa State University's Institute for Food Safety and Security, the Center for Crops Utilization Research, and the Iowa Agriculture Experiment Station. We would like to thank Bishnu Karki and Debjani Mitra for excellent laboratory assistance.
Disclosure Statement
No competing financial interests exist.