
Abstract
Transmission of antimicrobial resistance (AMR) from animal production systems to humans through the food supply is a public health concern. Currently, little is known about the prevalence of AMR among veal calves in the United States. Therefore, the objective of this prospective cohort study was to estimate the prevalence of AMR and multidrug resistance (MDR) among Escherichia coli within a vertically integrated production system. In addition, this study aimed to identify genes associated with phenotypic resistance to third- and fourth-generation cephalosporins (3GC and 4GC). Calves from four veal cohorts were randomly sampled resulting in a total of 166 farm fecal samples, 159 harvest fecal swabs, 164 preevisceration swabs, and 122 final carcass swabs. The prevalence of MDR among random-pick E. coli isolates recovered from the respective samples was 97% (161/166), 35% (55/159), 61% (51/84), and 24% (5/21). A selective isolation protocol found cefotaxime (a 3GC)-resistant isolates in 91% (127/140) of farm fecal samples, 34% (55/164) of preevisceration swabs, and 19% (23/122) of final carcass swabs tested. Isolates resistant to cefepime, a 4GC, were found among 24% (33/140), 6.7% (11/164), and 0.8% (1/122) of the same, respective samples. Isolates resistant to ciprofloxacin, a fluoroquinolone, were recovered from 75% (73/98) of farm fecal samples, 23% (38/164) of preevisceration swabs, and 6.6% (8/122) of final carcass swabs. The bla CMY-2 and bla CTX-M resistance genes were found in 89% (93/105) and 100% (42/42) of tested subsets of 3GC- and 4GC-resistant isolates, respectively. Pulsed-field gel electrophoresis (PFGE) analysis conducted on 3GC- and fluoroquinolone-resistant isolates showed three indistinguishable PFGE patterns from cefotaxime-resistant isolates recovered at farm and from two preevisceration carcass swabs. Although the prevalence of resistance declined between initial farm fecal samples and final carcass swabs, resistant bacteria recovered from carcasses illustrate the potential transmission of AMR to the human food supply.
Introduction
A
Antimicrobial use in food animals for prevention or treatment purposes (McEwen and Fedorka‐Cray, 2002) creates a selection pressure that contributes to the emergence, persistence, and transmission of antimicrobial resistance (AMR) through the food supply. Harvest-level sanitation and interventions are effective at reducing carcass contamination; nonetheless, complete elimination is likely impossible. Thus, preharvest interventions to reduce the prevalence of AMR may be necessary to mitigate transmission.
Previous research suggests that the prevalence of AMR in calves decreases with age (Hoyle et al., 2004). Since veal calves reach market maturation at a younger age than conventional beef cattle, the veal production system provides a unique opportunity to study the transmission of resistance in a high-risk population. Therefore, the objective of this study was to estimate the prevalence of AMR among Escherichia coli from a vertically integrated veal production system. In this system, the same company purchases all calves, contracts the growers, owns the harvest facility, and markets the veal products under one brand. In addition, the study aimed to illustrate the potential transmission of bacteria resistant to critically important antimicrobials from farm to carcass.
Materials and Methods
Study design and setting
A prospective cohort design was used to identify within-cohort changes in the prevalence of AMR. A convenience sample of four calf cohorts on separate grower farms, within 1 h of each other, was selected. In the production system, male dairy calves, age 1–3 d, were purchased at auctions in the Eastern United States and grouped into cohorts averaging 175 calves (range: 116–212) to fill contracted grower farms in the Midwest where they were intensively raised. Calves were housed in individual stalls for 9 weeks before transitioning to paired housing. Cohorts one and three were raised in enclosed barns with fan and tube ventilation; barns housing cohorts two and four were naturally ventilated with sidewall curtains. All calves were fed proprietary milk replacer produced without antimicrobials. Cohort two was provided supplemental grain containing 16% crude protein. Around 6 months of age, the calves were transported to a single harvest facility. Industry standard sanitary procedures were used for dehiding and evisceration. Sanitation vacuums were used to apply 180°F water to remove visual contamination following dehiding. A peroxyacetic acid wash was applied at 212–400 ppm to sanitize the carcass before entering the carcass chiller.
Samples were collected at four time points: fecal samples on farm, fecal samples from viscera, preevisceration carcass swabs, and final carcass swabs (Fig. 1). On a single farm visit, fecal samples were collected from 42 calves, ∼110 d (range: 103–115) old, using systematic random sampling (e.g., every fourth calf, depending on barn total). Sampled calves were given ear tags for identification at harvest. A total of 611 samples were collected from 166 calves distributed across four cohorts, including 166 farm fecal samples, 159 harvest fecal samples, 164 preevisceration swabs, and 122 final carcass swabs. Variations in sample numbers were due to missed sampling opportunities during carcass processing. Detailed herd or calf-level data on antimicrobial administration were unavailable; however, producers stated calves never received fluoroquinolones.
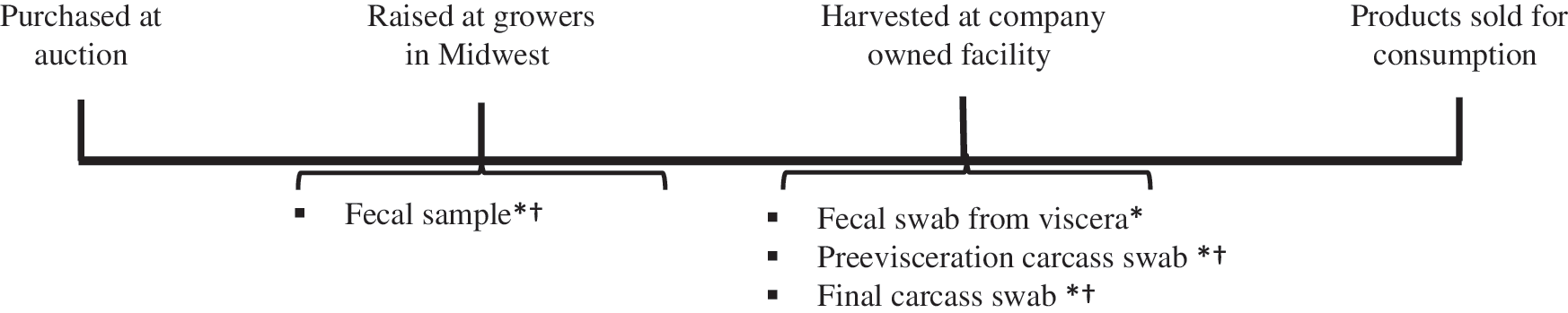
Veal calf production and sampling timeline. *Sample subjected to isolation protocol for random-pick Escherichia coli and testing with Kirby-Bauer disk diffusion assay. †Sample subjected to protocol for selective isolation of cephalosporin- and fluoroquinolone-resistant E. coli using MacConkey agars containing antimicrobials.
Sample collection
Farm fecal samples were collected using digital rectal palpation with clean nitrile gloves and placed into sterile bags (Nasco, Fort Atkinson, WI). Harvest fecal samples were collected from the rectum following evisceration using sterile swabs. Preevisceration and final carcass swabs were collected from a 10 by 10 inch area behind the shoulder using sterile sponges (Nasco) saturated with 20 mL tryptic soy broth (TSB) (Arthur et al., 2004). Preevisceration swabs were collected from the left side of the carcass following dehiding and before evisceration. Final carcass swabs were collected from the right side of the carcass within 2 h after placement in the chiller. Samples were transported on ice to the laboratory and kept at 4°C until processing.
Laboratory methods
A nonselective culture approach was applied within 24 h of sample collection to isolate a single, random E. coli isolate. In addition, a selective culture process was applied within 1 week of collection to isolate E. coli resistant to 3GC, 4GC, fluoroquinolones, and carbapenems. To accommodate laboratory capacity, the selective protocol was applied to farm fecal samples, preevisceration, and final carcass swabs. Only 15 of the 41 fecal samples from cohort four were subjected to the selective protocol. Cohort one fecal samples were only tested for cephalosporin resistance. Final carcass swabs from cohort four were unavailable.
For the isolation of random-pick E. coli, samples were plated onto MacConkey agar (Becton, Dickinson and Company, Sparks, MD). Before being plated, carcass swabs were processed using a previously reported method (Arthur et al., 2004); briefly, 80 mL of TSB was added and swabs were incubated at 25°C for 2 h and then 37°C for 16–18 h.
For the selective protocol, two 4 g fecal aliquots were homogenized and incubated for 18–24 h at 37°C in 36 mL of MacConkey broth containing 16 μg/mL nalidixic acid or 2 μg/mL cefotaxime. One milliliter aliquots from processed carcass swabs was incubated in 9 mL of nutrient broth containing the respective antibiotics. Broths containing nalidixic acid were used to inoculate MacConkey agars supplemented with 16 μg/mL nalidixic acid and 2 μg/mL ciprofloxacin. The broths containing cefotaxime were used to inoculate MacConkey agars supplemented with 8 μg/mL cefoxitin, 4 μg/mL cefepime, and 0.5 μg/mL meropenem (Mollenkopf et al., 2012).
Plates were incubated for 18–24 h at 37°C. One colony phenotypically consistent with E. coli was selected from each agar and indole tested. Lactose-positive, indole-positive colonies lacking a mucoid appearance were presumed to be E. coli (Duse et al., 2015).
Antimicrobial susceptibility testing
Kirby-Bauer disk diffusion assays (Clinical and Laboratory Standards Institute [CLSI], 2015) were performed on random-pick E. coli isolates to identify phenotypic resistance to 12 antimicrobials (Becton, Dickinson and Company, Sparks, MD) (Fig. 2). Isolates were categorized as susceptible, intermediate, or resistant using established breakpoints (Clinical and Laboratory Standards Institute, 2015); intermediate and resistant isolates were considered nonsusceptible. CLSI breakpoints for ceftiofur in E. coli are not established and the observed bimodal distribution of zone diameters for ceftriaxone was inconsistent with established values. Therefore, previously reported epidemiological cutoffs consistent with the observed zone diameter distributions (≤22 mm and ≤26 mm for ceftiofur and ceftriaxone, respectively) were used to categorize isolates as nonsusceptible (Pereira et al., 2011).

Proportion of random-pick E. coli, isolated from four sampling time points in a vertically integrated veal calf production system, phenotypically nonsusceptible to tested antimicrobials. AMP, ampicillin; CIP, ciprofloxacin; AXO, ceftriaxone; CHL, chloramphenicol; FOX, cefoxitin; GEN, gentamicin; NEO, neomycin; NAL, nalidixic acid; STR, streptomycin; SXT, sulfamethoxazole–trimethoprim; TET, tetracycline; XNL, ceftiofur. Letters over bars are used to indicate statistical significance. Within an antimicrobial, bars with shared letters are not statistically different.
Identification of bla CMY-2 and bla CTX-M genes
A subset of isolates (n = 105) from the selective protocol resistant to cefoxitin (2GC) and cefotaxime (3GC) was selected for polymerase chain reaction (PCR) to identify the bla CMY-2 gene. The forward primer from Alali et al. (2009) and the reverse primer from Winokur et al. (2001) were used to target a region of 559 bp. Isolates phenotypically resistant to cefotaxime (3GC) and cefepime (4GC) (n = 42) were subjected to PCR using a global primer (Lewis et al., 2007) for identification of the bla CTX-M gene.
Pulsed-field gel electrophoresis
Two subsets of 24 isolates, one resistant to fluoroquinolones and the other positive for the bla CMY-2 gene, were selected and analyzed separately with pulsed-field gel electrophoresis (PFGE) to describe the genotypic diversity of resistant isolates. For each subset, up to three isolates per cohort per time point were selected, preferentially selecting isolates from calves positive at two or more time points.
PFGE was performed using the established PulseNet protocol for non-O157 E. coli and the XbaI (Roche, Basel, Switzerland) restriction enzyme (Ribot et al., 2006). Gels were run using a Chef-Mapper DR-III (Biorad, Hercules, CA) system. Analysis of similarities was performed in GelCompar 6.5 (Applied Maths, Inc., Austin, TX) using Dice coefficient with 1% tolerance and 1% optimization.
Statistical analysis
The prevalence of resistance was calculated by dividing the number of nonsusceptible isolates by the total number of isolates recovered. To test the hypothesis that prevalence of resistance changed through production stages, the isolate-level categorization for each antimicrobial was included as the response variable in separate logistic regression models (PROC GLIMMIX; SAS v. 9.4, Cary, NC). Cohort and sampling time point were included as fixed effects.
Four different logistic regression models were used to test the hypothesis that the odds of a sample being positive for isolate resistant to cefoxitin, cefepime, nalidixic acid, or ciprofloxacin was different between cohorts. Cohort and time point were included as fixed effects. The interaction between cohort and sampling stage was included to identify differences in changes between farm and harvest for each of the cohorts. p-Values <0.05 were considered statistically significant.
Results
Prevalence of AMR in random-pick E. coli
E. coli was isolated from all (166/166) farm fecal samples. Farm prevalence of AMR among random-pick E. coli was >90% for ampicillin, neomycin, streptomycin, sulfamethoxazole–trimethoprim, and tetracycline (Fig. 2). Random-pick E. coli was isolated from all harvest fecal samples (159/159) and 51% (84/164) of preevisceration and 17% (21/122) of final carcass swabs. For ten antimicrobials, resistance was significantly lower (p < 0.05) in harvest fecal isolates relative to farm fecal isolates (Fig. 2); however, resistance among preevisceration isolates was significantly higher than harvest fecal isolates for five antimicrobials (p < 0.05) (Fig. 2). Detailed cohort prevalence estimates can be found in Supplementary Table S1 (Supplementary Data are available online at
Results formatted with the number of isolates exhibiting the resistance phenotype followed by (percentage of isolates).
AMP, ampicillin; AXO, ceftriaxone; CHL, chloramphenicol; FOX, cefoxitin; GEN, gentamicin; NEO, neomycin; STR, streptomycin; SXT, sulfamethoxazole–trimethoprim; TET, tetracycline; XNL, ceftiofur.
For this study, multidrug resistance (MDR) was defined as resistance to three or more classes of antimicrobials. MDR was found among 97% (161/166) of farm fecal sample isolates, 35% (55/159) of harvest fecal isolates, 61% (51/84) of preevisceration isolates, and 22% (5/21) of final carcass swab isolates (Fig. 3).
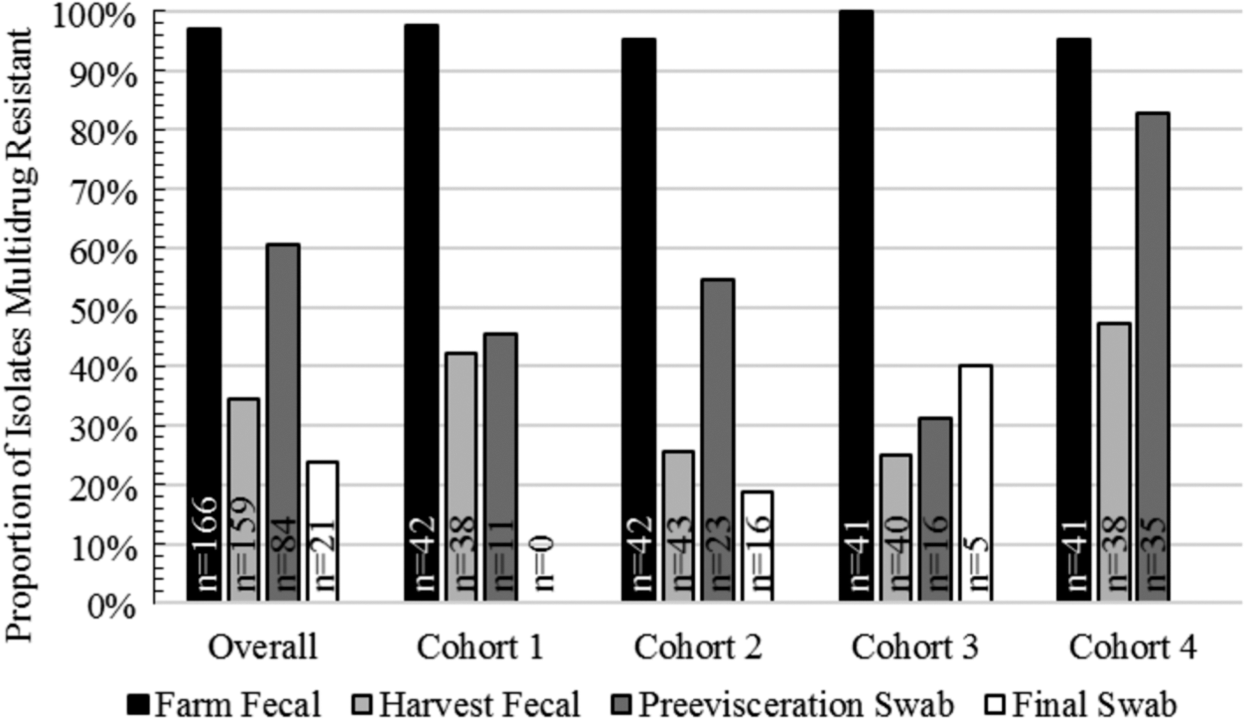
Proportion of random-pick E. coli, isolated from four separate calf cohorts at four sampling time points in a vertically integrated veal calf production system, phenotypically multidrug resistant. “n” represents the number of isolates recovered from each cohort at each time point. Multidrug resistant is defined as resistant to three or more classes of antimicrobials.
Selective isolation of cephalosporin- and fluoroquinolone-resistant E. coli
Using selective isolation, 88% (86/98), 75% (73/98), 91% (127/140), and 24% (33/140) of farm fecal samples possessed E. coli resistant to nalidixic acid, ciprofloxacin, cefoxitin, and cefepime, respectively (Table 2). Isolates resistant to cefoxitin and cefepime were also resistant to cefotaxime and had phenotypes consistent with the bla CMY-2 and bla CTX-M genes, respectively. There were substantial differences in the prevalence of resistance between cohorts; 94% (31/33) of isolates with the CTX-M phenotype were from cohort three. Samples from cohorts one (0/41) and two (0/41) were negative for cefepime-resistant E. coli. No meropenem-resistant isolates were found.
Results are formatted with number of resistant samples followed by (proportion of total samples).
Cohort 1 fecal samples were not analyzed for isolates possessing resistance to quinolones.
Final swabs from cohort 4 were unavailable for analysis.
At harvest, 37% (61/164), 23% (38/164), 34% (55/164), and 6.7% (11/164) of all preevisceration carcass swabs contained isolates resistant to nalidixic acid, ciprofloxacin, cefoxitin, and cefepime, respectively (Table 2). Final carcass swabs had a significantly lower prevalence with 13% (16/122), 6.6% (8/122), 19% (23/122), and 0.8% (1/122) containing a resistant isolate to the respective antimicrobials. Substantial variation in prevalence was seen between harvested cohorts (Table 2). For example, 82% (9/11) of cefepime-resistant preevisceration isolates were from cohort four. The prevalence of samples resistant to ciprofloxacin, nalidixic acid, and cefoxitin was significantly different between cohorts (p < 0.05). In addition, the interaction between cohort and time point was significant (p < 0.05) for nalidixic acid and ciprofloxacin, suggesting temporal changes in prevalence between farm sampling and carcass contamination are different between farms.
bla CMY-2 and bla CTX-M PCR
PCR confirmed the presence of the bla CMY-2 gene in 89% (93/105) of the selected isolate subset. From the 45 isolates resistant to cefepime, 42 were analyzed and were positive for the bla CTX-M gene.
Pulsed-field gel electrophoresis
PFGE demonstrated a diverse population of resistant E. coli. Among the 24 ciprofloxacin-resistant isolates, there were 20 unique patterns with four pairs of indistinguishable patterns (Fig. 4). Two indistinguishable pairs of isolates were recovered within farm samples and final carcass swabs; each pair was from the same calf cohort and time point. There were no pairs of indistinguishable patterns recovered between farm and harvest.
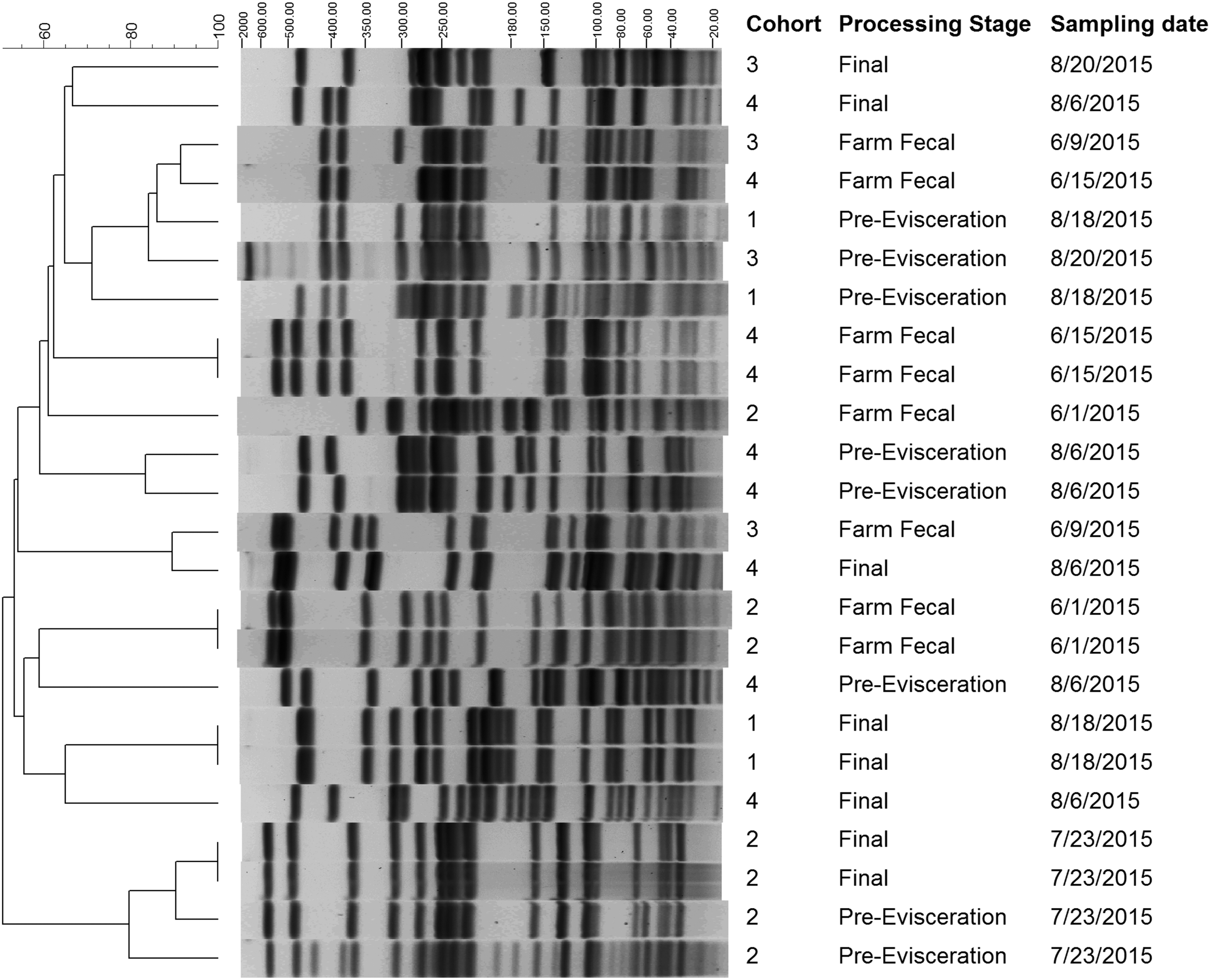
Pulsed-field gel electrophoresis of ciprofloxacin-resistant E. coli isolates recovered from farm fecal samples, preevisceration carcass swabs, and final carcass swabs, collected from four separate cohorts in a vertically integrated veal calf production system, and isolated using selective media. Isolates were analyzed using the Chef-Mapper DR-III system, XbaI enzyme, and PulseNet Protocol for non-O157 E. coli.
From the 24 isolates possessing the bla CMY-2 gene, there were 21 unique patterns and two indistinguishable sets of isolates. A pair of farm fecal samples from cohort three was indistinguishable, and a group of indistinguishable isolates was recovered from a cohort one fecal sample and two preevisceration swabs (Fig. 5).

Pulsed-field gel electrophoresis of E. coli isolates, possessing the bla CMY-2 gene, recovered from farm fecal samples, preevisceration carcass swabs, and final carcass swabs, collected from four separate cohorts in a vertically integrated veal calf production system, and isolated using selective media. Isolates were analyzed using the Chef-Mapper DR-III system, XbaI enzyme, and PulseNet Protocol for non-O157 E. coli.
Discussion
To our knowledge, this is the first study in the United States to estimate the prevalence of AMR in veal. Therefore, our results can be cautiously compared to conventional dairy calf studies and veal studies conducted in other countries. Differing estimates may be attributed to variations in diet, housing, antimicrobial use, or other management practices. When considering our results, random-pick E. coli isolates are intended to serve as indicators of resistance levels within commensal bacterial population. The selective isolates, which may only represent a small portion of the microbial population, are useful as a sensitive method to estimate the sample-level prevalence resistance to critically important antimicrobials. The pathogenic potential of the recovered bacteria was not assessed; nonetheless, foodborne transmission of bacteria harboring horizontally transferable genetic elements is likely to have an important public health impact.
In this study, MDR was found among 97% of farm fecal E. coli. Although antimicrobial use was unknown, the selection pressure from therapeutic use of antimicrobials may play a role in observed levels of resistance (Bosman et al., 2014). Although resistance may, in part, be attributed to the age of the calves (Khachatryan et al., 2004), the prevalence of MDR was higher than younger (2–8 d) conventional dairy calves in New York (Pereira et al., 2011) where 81% of fecal E. coli isolates were multidrug resistant.
In our results, ∼25% of random-pick isolates were nonsusceptible to 3GC, which is higher than veal studies in Germany (prevalence: 1.4–3.0%) (Kaesbohrer et al., 2012) and the Netherlands (prevalence: 0.7–2.0%) (van Geijlswijk, 2015), and the dairy calves in New York (prevalence: ∼10%). In total, 90.7% (range 73.3–97.6) and 23.6% (range 0–73.8) of samples plated onto selective media yielded isolates resistant to cefotaxime and cefepime, respectively. These results are higher than a Swedish study in which 11% of dairy calves had cefotaxime-resistant isolates (Duse et al., 2015).
The prevalence of random-pick isolates nonsusceptible to quinolones and fluoroquinolones (4.8%, 2.4%) are lower than reported in Germany (10.5%, 13.3%) and the Netherlands (10.7%, 12.1%), but slightly higher than the conventional dairy calves in New York (<1%, 0%). Selective protocol results are higher for nalidixic acid, 89% (range 66.7–97.6%), than the Swedish study (49%). No comparable studies using selective protocols for fluoroquinolone resistance were found. Ciprofloxacin resistance found in 75% (range 26.7–97.6) of farm fecal samples is concerning because the calves never received fluoroquinolones. Fluoroquinolones are labeled for treatment of bovine respiratory disease in dairy replacement heifers, but are prohibited for extralabel use (Extralabel Drug Use in Animals, 2017). However, the veal calves originated from conventional dairy farms and were processed through auction facilities. In these environments, it is possible that calves may have come in contact with other livestock (Duse et al., 2015) or surfaces possessing resistant bacteria. These bacteria may have colonized the calves and been subsequently maintained.
The decrease in resistant E. coli isolates between farm and harvest fecal samples may be attributed to an age-related decline (Hoyle et al., 2004); however, antimicrobials are routinely withdrawn before harvest, which is likely to play a role. The prevalence of resistance among harvest fecal samples was greater than other reports in veal. Only 36.5% (58/159) of harvest fecal isolates were pansusceptible, compared to 93% (164/176) reported in Australia (Barlow et al., 2015).
Our study additionally documented substantial variations in the proportion of carcasses where E. coli was recovered. This may be explained by variations in adherence to established harvest-level intervention protocols and/or the efficacy of the harvest-level interventions. Carcass contamination may occur through contact with the intestinal contents, the animal's hide, or the plant environment. Hides, likely carrying bacteria from the farm environment, have been shown to be a significant source of carcass contamination (Elder et al., 2000). The results showed levels of resistance among preevisceration isolates were higher relative to harvest fecal isolates, but similar to farm fecal isolates. This suggests that hides were an important source of carcass contamination with farm environment bacteria. In addition, preevisceration and farm fecal isolates had shared resistance phenotypes and PFGE patterns. The recovery of E. coli from final swabs, after the peroxyacetic acid, may suggest that the sanitizing wash was ineffective or carcasses were contaminated after entering the chiller. The interpretation of the prevalence estimates in this study is limited by the relatively small number of cohorts and the missing data from some cohorts. In addition, future studies should collect antimicrobial use and resistance data from a larger population to estimate the potential for antimicrobial use interventions to influence AMR.
Conclusion
The on-farm prevalence of E. coli with MDR and resistance to critically important antimicrobials were high. Harvest samples show the prevalence of resistance among random-pick isolates and selective isolates declined. PFGE demonstrated a genotypically diverse population of isolates resistant to fluoroquinolones or cephalosporins, with the potential for transmission between farm and harvest. Regardless of the observed decline through processing, resistant E. coli were recovered from preevisceration and final carcass swabs, demonstrating likely transmission through the food supply. Future studies should focus on preharvest and postharvest interventions to reduce the risk of transmission of resistant bacteria through the veal production system and examine the potential for reduced antimicrobial use to reduce levels of resistance.
Footnotes
Acknowledgments
We want to thank the veal producers, abattoir owners, and plant personnel for their collaboration. Authors gratefully acknowledge the assistance of Ann Ascot, Avery Strait, Rory O'Shaughnessy, Arati Sharma, Dixie Mollenkopf, and Dimitria Mathys for helping with sample collection and laboratory procedures.
Funding
This study was funded by internal funds from The Ohio State University College of Veterinary Medicine.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.