
Abstract
In this study, novel antimicrobial peptides had been derived by enzymatic hydrolysis of filefish (Thamnaconus modestus) byproduct (HFBP). Different proteases, (papain [P], trypsin [T], neutrase [N], pepsin [PE], and the mixture I [PN] and mixture II [PT]) and different hydrolysis time (60, 120, 180, and 240 min), have been used to generate peptides with different lengths and amino acid sequences. The antimicrobial properties of HFBPs were tested, against Gram-positive and Gram-negative strains, using disc diffusion method. HFBP prepared after 120 min of the enzymatic hydrolysis by trypsin (HFBP-T) exhibited greatest antibacterial activities. Bacillus cereus 10451 (BC) and Salmonella enteritidis 10982 (SE) strains were most sensitive to HFBP-T with an inhibitory zone of 24.68 and 29.67 mm diameter and minimum inhibitory concentration of 1.25 and 2.5 mg/mL, respectively. Moreover, the antimicrobial activities of tested HFBPs increased significantly at low pH and temperature. The amino acid analysis showed that HFBP-T protein hydrolysate is high in an amino acid of proline, which probably contributes to the antimicrobial activity. The results obtained from scanning electron microscopy suggested that HFBPs might kill bacteria by acting on the cell wall of bacteria. Conclusively, the HFBP derived from filefish byproduct with biological activates is an interesting alternative to the use of waste from the fishing industry as natural antimicrobials in food stuff.
Introduction
C
Among natural inhibitors of pathogenic bacteria, antimicrobial peptides (AMPs) derived from fish show promise. AMPs have been considered a part of innate immunity, playing major roles in protecting against a wide variety of pathogenic microorganisms including bacterial, fungal, and viral (Smith et al., 2010; Rajanbabu and Chen, 2011). Several studies have shown that fish AMPs demonstrate strong antibacterial activities against several Gram-negative and Gram-positive strains (Rajanbabu and Chen, 2011; Cruz et al., 2014; Falanga et al., 2016). Moreover, AMPs have anticancer activity, chemotaxis, antigen presentation, angiogenesis, inflammatory responses, and adaptive immune induction (Dong et al., 2008; Gaspar et al., 2013), which make them more desirable as potential ingredients for functional foods. It was also found that the enzymatically hydrolyzed fish skin gelatin has better biological activities compared to the peptides derived from fish muscle protein (Kim and Mendis, 2006). Most researchers have focused on the preparation of fish AMPs using the enzymatic hydrolysis of fish muscles (Song et al., 2012b; Jiang et al., 2014). However, there are few investigations have been done on AMPs from fish byproducts. The terms fish byproducts usually refer to viscera (liver, roe, stomachs, upon), heads, backbones, cuts, and rejected fish from processing. With increasing awareness in the fishing industry for optimal use of fishery byproducts to ensure the cost-effectiveness and minimize the environmental impact, AMPs derived from fish byproduct attracted the researcher's attention, recently. Filefish (Thamnaconus modestus) is one of the important fish in the Northwest Pacific Ocean (Fig. 1). In China, it is commercially fished, and it has been aquacultured successfully (Guan et al., 2011). In Chinese markets, these fish are sold without the skin and head, which are considered a waste and discarded, although it is a great source of minerals, proteins, and fat (Jayathilakan et al., 2012). This study is designed to extract peptides with antimicrobial properties from filefish byproducts (FB). To date, no studies have been done to isolate the bioactive peptides from FB. The skin provides fish with the first line of protection against water-borne microbes because it contains a collection of antibacterial substances, including AMPs (Subramanian et al., 2009; Su, 2011). Therefore, the objective of this study was to prepare bioactive peptides with antibacterial activity from FB by enzymatic hydrolysis. Several enzymes and their mixtures were used for the process of hydrolysis. The effect of different pH (4 and 7), temperatures (4°C, 25°C, and 37°C), as well as the incubation time on the antibacterial performance of peptide mixture was investigated.

Filefish (Thamnaconus modestus), the skin and head have been used in this study.
Materials and Methods
Materials
Fresh filefish (T. modestus) byproducts (FB), including the frame, head, and skin were collected from the local fish market in Zhoushan, China, and used as the raw material for protein isolate. The byproducts were ground into minces and sealed in plastic bags and stored at −20°C until used for peptide production. Papain (P), trypsin (T), neutrase (N), and pepsin (PE) were obtained from Sigma-Aldrich Chemical Co. (St. Louis, MO). All other chemicals and reagents were provided by Chemical Reagents Company (Zhoushan, China).
Microorganisms for antimicrobial testing
Staphylococcus aureus 10786 (SA), Bacillus cereus 10451 (BC), and Salmonella enteritidis 10982 (SE) were provided by China Center for Industrial Culture Collection (CICC; Beijing, China), while Escherichia coli GIM1.708 (E.coli) was obtained from Microbial Culture Collection Center of Guangdong (GIMCC; Guangdong, China). All strains were maintained as dry active strains and stored at −80°C. To reactivate the bacteria, SA, BC, SE, and E.coli were subcultured two to three times in Mueller-Hinton Broth (MHB) at 37°C for 18–24 h before use.
Isolation of filefish byproduct proteins
Fish proteins were isolated from FB using the pH-shift method, as described (Chomnawang and Yongsawatdigul, 2013). Briefly, the ground FB was mixed with cold distilled water (4°C) at a ratio of 1:9 (w/v) and homogenized for 1 min. The homogenate was transferred to a beaker placed in a container filled with crushed ice and 2 N NaOH has been used to adjust the pH to 11. The mixture was stirred for 25 min at 0–4°C using a magnetic stirrer. The suspension pH was kept constant during extraction by addition of 2 N NaOH or 2 N HCl. The solution was centrifuged at 10,000 rpm for 15 min at 4°C. The supernatant was poured through four layers of cheesecloth to remove solidified fat and insoluble parts, and then the supernatant's pH readjusted to pH 5.5 using 2 N HCl and allowed to stand for 10 min. The solution was centrifuged at 10,000 rpm for 15 min at 4°C. The sediment containing the protein isolates was collected and kept at −20°C until used in the enzymatic hydrolysis.
Enzymatic hydrolysis of filefish byproduct isolated protein
The filefish byproduct proteins (FBP) were mixed with distilled water at a ratio of 1:1 (w/v) as a substrate for the production of protein hydrolysate (hydrolysis of filefish byproduct [HFBP]) and then homogenized at a speed of 11,200 g for 1 min. The enzymatic hydrolysis was performed by using neutrase (N), papain (P), pepsin (PE), and trypsin (T) with a ratio of enzyme to substrate 1.5% (w/w) at their optimal conditions as shown in Table 1. The temperature and pH were kept constant during the hydrolysis. Twenty milliliter aliquots were withdrawn from the hydrolysate solution, at intervals, after 30, 60, 120, 180, and 240 min (the endpoint of hydrolysis process). The enzyme was deactivated directly after withdrawal of the aliquot by heating at 95°C for 10 min. The mixtures were centrifuged at 10,000 rpm at 4°C for 15 min. Then, the supernatants were used to determine the antimicrobial activities. The hydrolysate supernatant with the highest antimicrobial activity was freeze-dried and then preserved at −18°C for subsequent analysis. In the hydrolysis experiment using mixtures of two enzymes, hydrolysis conditions were adjusted to the optimal conditions of the first enzyme during the first half of hydrolysis and changed to optimal conditions of the second enzyme during the other half of hydrolysis process (Table 1). Two aliquots were withdrawn after 60, 120, 180, and 240 min to evaluate the antibacterial activity of each mixture. The mixture with the high antimicrobial activity was freeze-dried and then preserved at −18°C for subsequent analysis.
Antibacterial assay
The antimicrobial activities of the mixture of HFBP against the Gram-positive strains (SA, BC) and the Gram-negative strains (SE, E.coli) were evaluated through the determination of the inhibition zone (Jiang et al., 2014), with slight modifications. Sterilized Mueller-Hinton agar (MHA) medium (15–20 mL) was distributed into each sterile petri dish. After solidification, sterile swabs have been used to spread the test bacterial strain (106–108 colony-forming unit [CFU]/mL) evenly over the surface of the medium. The bacterial suspension has been adjusted to this concentration using a 0.5 McFarland Standard (Remel™; Thermo Fisher Scientific, Waltham, MA). Sterile blank paper discs (5 mm in diameter) were placed on each inoculated agar plate. Finally, 25 μL of sample from each hydrolysate was added to each filter paper. Then, the plates were incubated at 37°C for 24 h. The inhibition zone diameter was measured in millimeter, including the diameter of the filter paper. An equivalent volume of sterile water instead of the tested compounds was used as negative control.
Minimum inhibitory concentration assay
Sterile microtiter plates with 96 wells were used to determine the minimum inhibitory concentration (MIC) of FBP hydrolysate with highest antibacterial activities. FBP hydrolysates were prepared by the enzymatic hydrolysis using P, T, and PT and tested against different pathogenic microorganism strains according to the methods described by (Hatab et al., 2016) with slight modification. The sterile 96-well plates for each bacterial strain were set up as follows: in wells in row A, 200 μL portions of 80 mg/mL peptides were placed in sterile MHB. Wells from row B to H received 100 μL of sterile MHB. Serial dilution (with concentrations ranging from 80 to 0.625 mg/mL) was performed from row A to row H and then 100 μL was discarded from row H. Ten microliters of the adjusted bacterial suspension (106 CFU/mL by reference to McFarland stander) was added to each well before a 24-h incubation at 37°C. The absorbance (630 nm) of each well was read at 24 h with a microtiter plate spectrophotometer (iMark™ Microplate Absorbance Reader). A positive control (containing inoculums, but no peptide) and negative control (containing peptide, but no inoculums) were included on each microplate. The microbroth dilution assays were performed in triplicate. MIC was defined as the lowest concentration that causes complete inhibition of the growth of the tested microorganism.
Effect of pH and temperature on HFBP antimicrobial activities
The effects of different temperature (4°C and 25°C) and pH (4.0 and 7.0) on the antimicrobial activities HFBP and the growth of test strains were determined using the microbroth dilution method as described by Hatab et al. (2016).
Amino acid compositions of freeze-dried hydrolysates
Amino acids analyzer (Hitachi L-8900 Amino Acid Analyzer) has been used to determine the compositions of free amino acids of freeze-dried hydrolysates (Jin et al., 2012). All tests were performed in triplicate.
Scanning electron microscopy
The morphological changes of bacterial cells before and after treatment with HFBP were investigated using scanning electron microscopy (SEM; JEOL-model: JSM-6390 LV). The suspensions of different bacterial strains (500 μL) at the exponential phase were mixed with 500 μL of HFBP derived from different enzymatic hydrolysis and incubated at 37°C for 24 h. The mixtures were centrifuged at 10,000 rpm for 10 min. The supernatants were removed; the pellets were washed with phosphate-buffered saline (50 mM, pH 7.4) thrice and then fixed with 2.5% glutaraldehyde at 4°C for 4 h. To remove the glutaraldehyde, samples have been washed using buffered solution. A series of ethanol solutions (50%, 70%, 80%, 90%, and 100%) have been utilized to dehydrate the samples (10 min each rinse). The dehydrated samples were dried immediately in a critical-point dryer using CO2 (K850; EMITECH) and mounted on a stainless steel stab with double-stick tape, with a thin layer of gold in a high vacuum condition. The specimens were then examined with SEM.
Statistical analysis
All experiments were performed in triplicate, and the data were presented as mean ± standard deviation. Data were subjected to multiway analysis of variance, and the mean comparisons were performed by Tukey's multiple range test using SPSS 13.0 (SPSS, Inc., Chicago, IL). Statistical significance was considered to exist when p < 0.05.
Results and Discussion
In our study, the skin and head of filefish have been used to extract the protein hydrolysate (HFBP) with an antibacterial activity. Four different enzymes (including neutrase [N], papain [P], pepsin [PE], and trypsin [T]) and two mixtures (P + N [PN] and P + T [PT]) were used for the protein hydrolysis. The treatment of protein substrates with different proteases generates peptides with different lengths and amino acid sequences and thus exhibit varying degrees of biological activities (Nasri and Nasri, 2013; Taha et al., 2013).
Our results indicated that the hydrolysate from neutrase (N), pepsin (PE), and mixture I (PN) hydrolysis has no antibacterial activity against all tested strains at different hydrolysis time (data are not shown). However, HFBP derived after 120 min of hydrolysis by trypsin (HFBP-T), papain (HFBP-P), and the mixture II (PT [HFBP-PT]) were able to inhibit the growth of all tested bacteria, with various degrees of inhibition using the disc diffusion method as shown in Figure 2. It is a known fact that hydrolysis conditions, including time, temperature, and pH, as well as nature of the protein and the microorganism itself, greatly influence activities of resulted peptides (Imeldawy and Eunicecy, 2010; Taha et al., 2013; Nurdiani et al., 2015). HFBP-T has demonstrated the highest antibacterial activity against Gram-positive and Gram-negative bacteria. SE strain was most sensitive to HFBP-T with a significant zone of inhibition (29.67 mm), followed by BC strain (24.68 mm) (Fig. 3). A similar result was obtained with the peptide IK12, which is capable of disrupting the membrane of Salmonella enterica after 3 h of incubation (Kumaresan et al., 2015). On the contrast, HFBP-PT showed a weak antimicrobial activity against all tested strains, especially with E.coli (inhibition zone diameter 9.24 mm). Thus it is clear that using the mixture of two enzymes was not effective to produce an antibacterial hydrolysate.
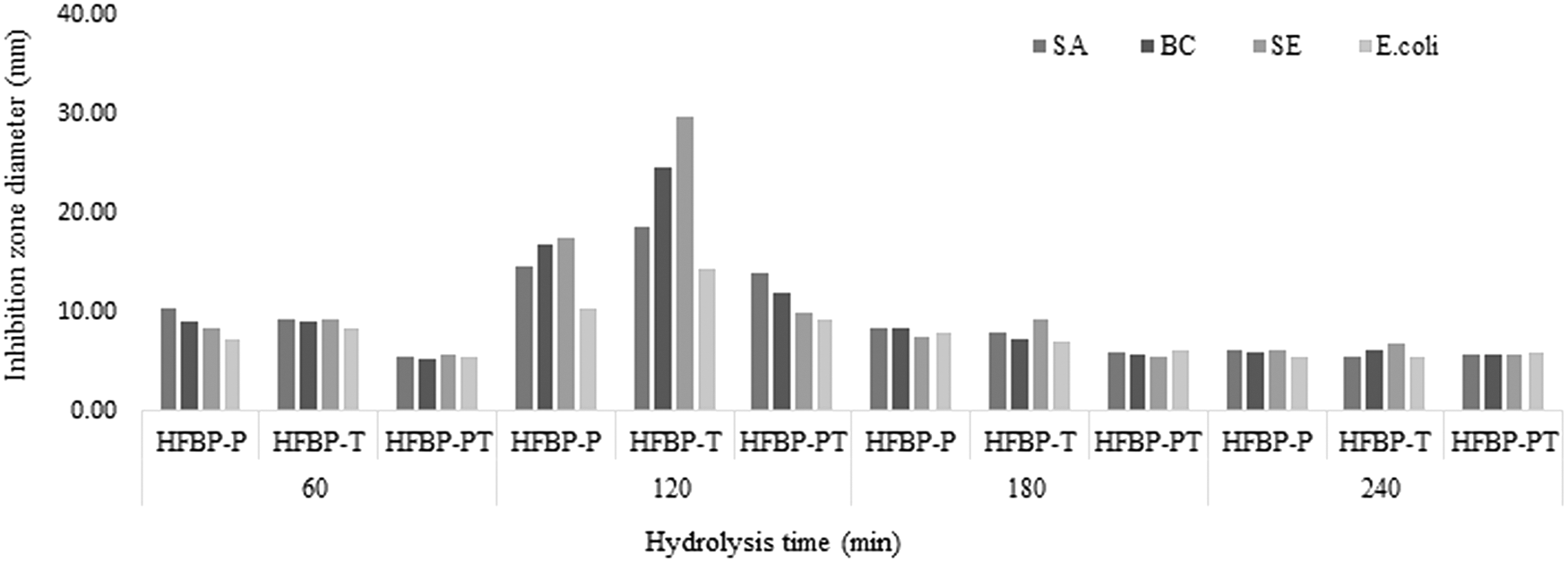
Antibacterial activities of peptides derived from the HFBP prepared by different protease treatments (papain [HFBP-P], trypsin [HFBP-T] and mixture II [HFBP-PT]), during different hydrolysis time, against some food pathogenic microorganisms. HFBP, hydrolysis of filefish byproduct; HFBP-P, papain-hydrolyzed filefish byproduct protein hydrolysate; HFBP-PT, papain+trypsin-hydrolyzed filefish byproduct protein hydrolysate; HFBP-T, trypsin-hydrolyzed filefish byproduct protein hydrolysate.
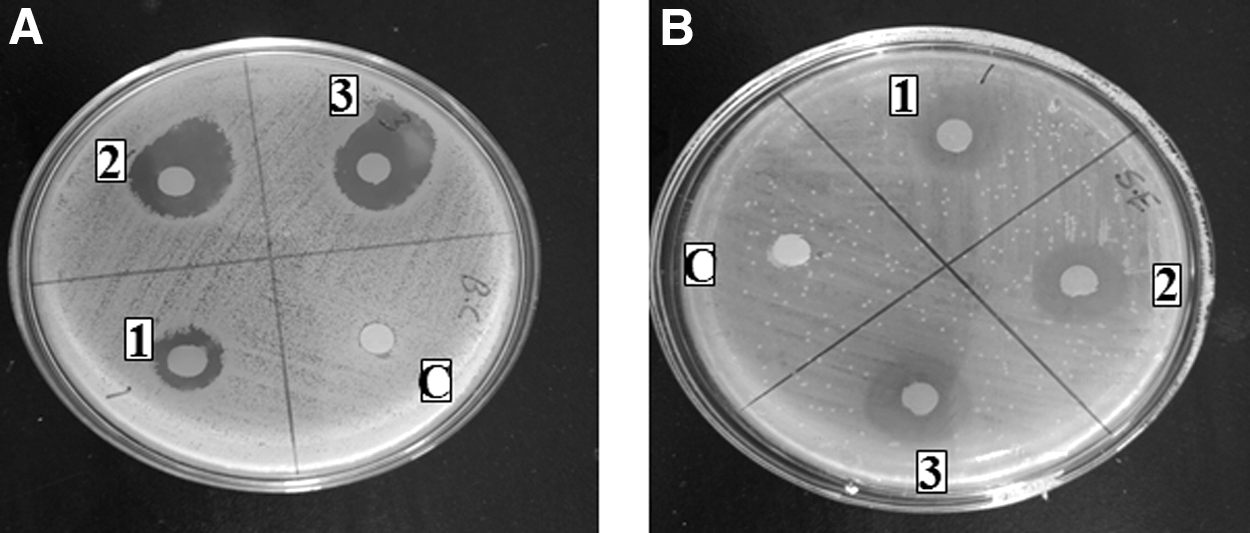
The zone of inhibition (mm) of HFBP-PT (1), HFBP-P (2), and HFBP-T (3) derived enzymatic hydrolysis of filefish for 120 min, at concentration 80 mg/mL on Bacillus cereus 10451
The MICs of HFBP-T, HFBP-P, and HFBP-PT were also determined and compared using the microbroth dilution method as summarized in Table 2. The results indicate that the lowest MIC was obtained with HFBP-T, which could inhibit the growth of SA, BC, SE, and E.coli strains at minimum concentrations of 5, 1.25, 2.25, and 5 mg/mL, respectively. BC strain was the most susceptible microorganism inhibited by the three hydrolysates, followed by SE. This result is in agreement with Mohd et al. (2009) who found that the palm kernel cake hydrolysates by trypsin have the lowest MIC on B. cereus compared with other tested strains. Song et al. (2012a) reported that half-fin Anchovy (Setipinna taty) hydrolysates derived from pepsin or trypsin probably contain antibacterial peptides with cationic and/or amphipathic characteristics. Demers-Mathieu et al. (2013) studied the antibacterial activities of peptide fraction obtained from hydrolysis of whey protein by trypsin; the results show that this fraction was able to inhibit the growth of Listeria monocytogenes and S. aureus at concentrations ranging from 10 to 20 mg/mL in the culture broth. Another possible reason for the highest antibacterial activity of HFBP-T could be due to a greater quantity of small molecular weight peptides, which contributed to a greater antimicrobial activity (Burris and Parks, 2004).
Hydrolysates were obtained after 120 min of enzymatic hydrolysis. HFBP-P, HFBP-T, and HFBP-PT referred to the hydrolysate obtained by papain, trypsin, or the mixture of papain and trypsin, respectively. The experiment was repeated thrice.
BC, Bacillus cereus 10451; E.coli, Escherichia coli GIM1.708; HFBP, hydrolysis of filefish byproduct; SA, Staphylococcus aureus 10786; SE, Salmonella enteritidis 10982.
For an accurate evaluation of the antimicrobial activity of the obtained peptides, the effect of temperature, pH, and incubation time on the growth of the selected strains was assessed without the addition of peptides by optical density (OD) measurement. Figure 4A–D show that there were significant (p < 0.05) effects of the pH, temperature, and incubation time on the growth curves of all tested strains. Our results indicate that the treatments used, pH 4 at 4°C and pH 7 at 4°C, could decrease the growth of all tested strains significantly (p < 0.05) over the incubation period. OD recorded significant changes (p < 0.05) with all strains after 48 h of incubation under similar conditions. Surprisingly, all strains were able to grow and revive exponentially at pH 4 at 25°C within the incubation time (48 h). Similar results have been reported in our previous work with Shiga-toxigenic Escherichia coli (STEC), where the tested strains showed faster growth at pH 4 and 25°C than 4°C at pH 4.0 or 7.0 (Hatab et al., 2016). A typical growth profile occurred at pH 7 and 25°C, whereas after 48 h of incubation, the OD reached 0.55, 0.567, 0.622, and 0.643 for SA, BC, SE, and E.coli, respectively. The significant differences between the bacterial growth at 4°C and 25°C, confirm the importance of low temperature for the safe storage of foods (Burt and Reinders, 2003).
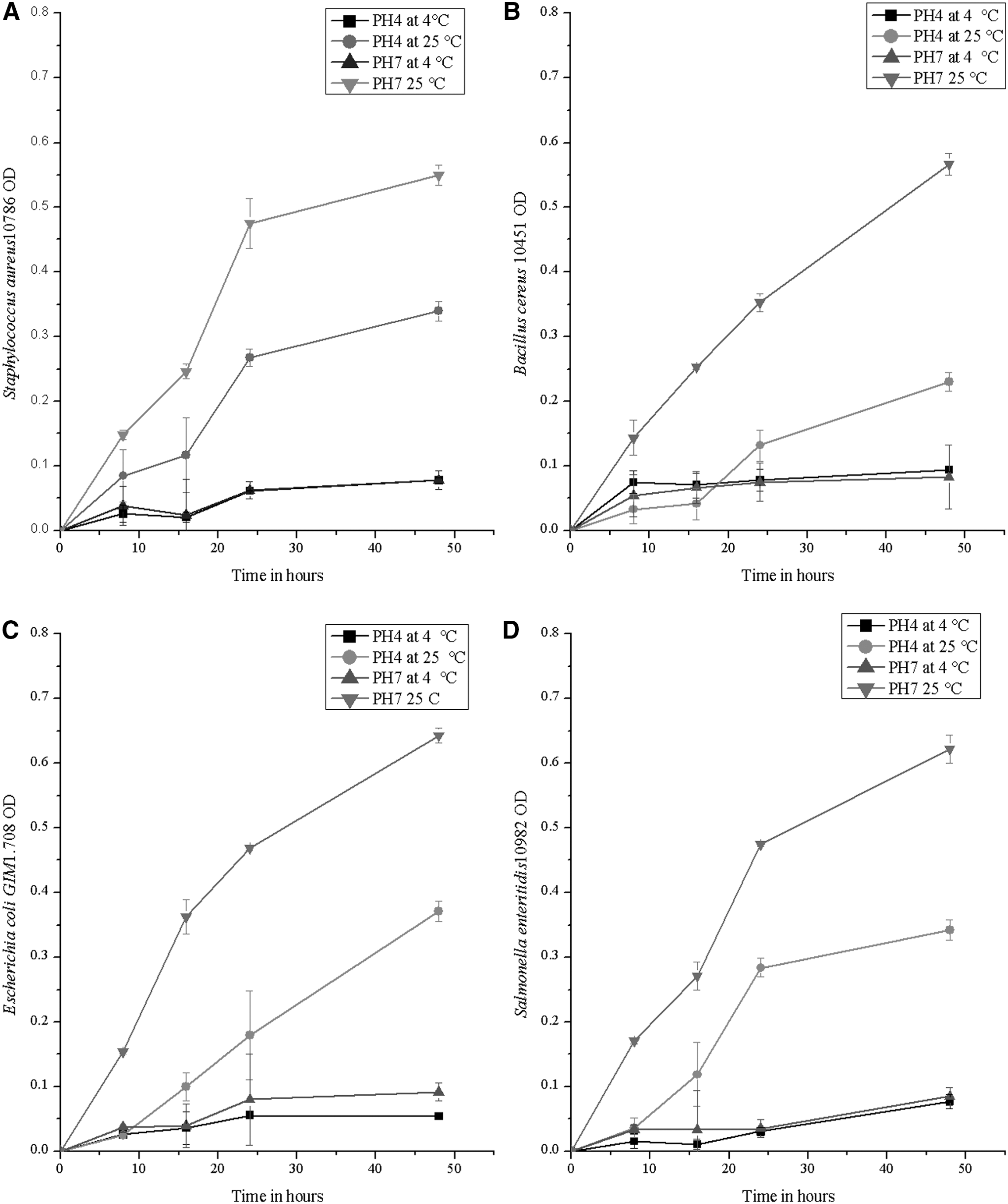
Survivor curves of food pathogenic microorganisms used in this study.
The effect of temperature and pH on the antimicrobial performance of generated peptides against Gram-negative and Gram-positive bacteria was investigated. The antimicrobial activities of HFBP-T, HFBP-P, and HFBP-PT were tested by determining the MICs of the peptides at a variety of pH (4.0 and 7.0) and temperature (4°C and 25°C) conditions. As shown in Table 3, the overall influence of pH and temperature on the antibacterial activity of HFBP-T, HFBP-P, and HFBP-PT against all strains was similar. The antibacterial activities of the three hydrolysates increased significantly (p < 0.05) with the decrease of pH and temperature. The lower amount of HFBP (0.625 mg/mL) was required to inhibit/kill all the tested strains at pH 4 and 4°C. The HFBP-T was found to be the most effective peptides, under the conditions of pH 4 at 4°C as well as pH 4 at 25°C, since 1.25 mg/mL was able to inhibit the growth of BC and SE. While under in the condition of pH 7 at 4°C the same amount (1.25 mg/mL of HFBP-T) was able to prevent the growth of all bacteria. It is well known that the low pH could inhibit the microbial growt by affecting nutrient transport, energy production, and enzymatic activity (Lambert et al., 2001; Burt, 2004; Hatab et al., 2016). On the contrary, Anderson and Yu (2005) reported that the activities of AMPs derived from ovine neutrophils decreased in acidic conditions than in basic conditions, and the optimum antimicrobial activities were obtained at pH 8.
HFBP-P, HFBP-T, and HFBP-PT were obtained after 120 min of the enzymatic HFBP using papain, trypsin, or the mixture of papain and trypsin, respectively. Three independent experiments were conducted for each treatment. The strains used are SA, BC, SE, and E.coli.
BC, Bacillus cereus 10451; E.coli, Escherichia coli GIM1.708; HFBP, hydrolysis of filefish byproduct; SA, Staphylococcus aureus 10786; SE, Salmonella enteritidis 10982.
To elucidate the possible reasons behind the different antimicrobial activities of generated peptides by different enzymes, the amino acid composition of each hydrolysate has been analyzed. The amino acid compositions of HFBP-T, HFBP-P, and HFBP-PT are given in Table 4. In general, the most abundant amino acids in all hydrolysates were the following: Thr, Cys, Met, Ile, Leu, Phe, Lys, and His, which are considered essential amino acids for a human. This fact is very important in food application of protein hydrolysates as a nutritious supplement or as a natural antimicrobial agent. Similar findings have also been obtained by the enzymatic hydrolysis for little hairtail (Trichiurus haumela) by Alcalase and Flavourzyme (Jin et al., 2012). On the other hand, the results showed that Asp, Thr, Ser, Met, and Gly contents were noticeably higher in HFBP-P hydrolysate as well as in HFBP-PT hydrolysate, in comparison with the HFBP-T samples. Moreover, Met was detected only in the in HFBP-P (0.525 nmol/20 μL) and HFBP-PT (0.267 nmol/20 μL). In contrast, Pro content was noticeably higher in HFBP-T samples. Previous researchers have reported that the most active antibacterial peptides against Gram-negative pathogens belong to the proline-rich peptide family (Burris and Parks, 2004; Mohd et al., 2009). Variance in the amino acid amount and composition between HFBP-T, HFBP-P, and HFBP-PT could be the reason for their different antimicrobial activities, which seem to be related to the composition of their amino acids (Gómez-Guillén et al., 2010). In this study, glycine and alanine constitute 17.82% of the total amino acids of HFBP-P and HFBP-PT. In the case of hydrolysate produced by trypsin (HFBP-T), glycine and alanine was constituted about 16.65% of the total amino acids. The high ratio of these amino acids resulted in increasing the hydrophobic properties of the produced hydrolysates, which increase the ability of the hydrolysates to bond with the bacterial cell wall (Kumaresan et al., 2015). The highest content of alanine, valine, leucine, lysine, arginine, and proline amino acids was observed with HFBP-T (42.96%) in total detected amino acids. It has been reported that these amino acids frequently appeared in the amino acid sequence of AMPs (Song et al., 2012a).
Determinations were performed in triplicate and data correspond to mean values.
AA, total amino acids; EAA, total essential amino acids, containing Thr, Cys, Val, Met, Ile, Leu, Phe, Lys, and Hys; HFBP-P, papain-hydrolyzed filefish byproduct protein hydrolysate; HFBP-PT, papain+trypsin-hydrolyzed filefish byproduct protein hydrolysate; HFBP-T, trypsin-hydrolyzed filefish byproduct protein hydrolysate; %HAA, percentage of total hydrophobic amino acids, containing Gly, Ala, Val, Met, Ile, Leu, Phe, Pro, and Trp.
Surface characterization of the tested bacterial cells was performed before and after treatment with AMPs using SEM. The changes of the cell membrane of BC (as Gram positive) and SE (as Gram negative) treated with HFBP-T were shown in Figure 5. Untreated groups showed smooth surface of BC and SE without any apparent cellular debris (Fig. 5A, C), while after treatment with HFBP-T, the cell wall appeared turgid and rough, as well as more pores were formed on the bacterial surface. The results given in Figure 5B and D indicate that the HFBP-T is causing membrane permeabilization. It is supposed that the HFBP-T interacts with the cell membrane, which causes membrane disruption and pore formation, allowing reflux of essential ions and nutrients (Taha et al., 2013; Song et al., 2016). In addition, it has been demonstrated that the anionic charges on the outer envelope of Gram-negative bacteria attracted the cationic antibacterial peptides; this action with hydrophobic properties of peptides enables them to perturb the cell membrane integrity or translocate into the cytoplasm (Friedrich et al., 2000; Song et al., 2012b). It is also possible that antibacterial peptides kill bacteria in a mechanism of nonmembrane disruption by intracellular binding molecules, and thus inhibit the cell wall biosynthesis and DNA, RNA, and protein synthesis (Song et al., 2012b; Kumaresan et al., 2015).

Scanning electron microscopy of BC and SE incubated without and with HFBP-T at 37°C for 24 h.
Conclusions
As a conclusion, this study indicated that the protein hydrolysates derived from enzymatic hydrolysis of FB using T, P, or the mixture II (PT) could potentially serve as natural antibacterial agents against pathogenic bacteria in the food industry. The antimicrobial activities of the generated peptides depend highly on several factors, including (1) used proteases; (2) hydrolysis time; (3) type of microorganism; and (4) inhibition condition (incubation temperature and pH). The HFBP-T prepared by trypsin has a significant inhibitory effect on both BC (Gram positive) and SE (Gram negative); these strains have a significant impact on food safety. The highest antibacterial activities of HFBP-T are probably due to the high content of proline amino acids. Further work is going to be performed to isolate and purify the bioactive peptide fractions from the hydrolysates, to elucidate the nature and structure of the fractions, and thus discern the mechanisms of antimicrobial action and the effects of different food matrix components on this activity.
Footnotes
Acknowledgments
This work was supported by a project granted by Chinese National Science and Technology (Project 2015BDA17B01), Chinese International Technology Cooperation Projects (Project 2012DFA30600), National Natural Science Foundation of China (NSFC31471609), as well as the program of Zhejiang Aquatic Products Processing Technology Research Joint Laboratory (Zhejiang Ocean University, ZhouShan, China), Key Science and Technology Project of Dinghai District (2015), and Zhejiang province science funding (LQ15C200010).
Disclosure Statement
No competing financial interests exist.