
Abstract
In the beef industry, product contamination by Salmonella enterica is a serious public health concern, which may result in human infection and cause significant financial loss due to product recalls. Currently, the precise mechanism and pathogen source responsible for Salmonella contamination in commercial establishments are not well understood. We characterized 89 S. enterica strains isolated from beef trim with respect to their biofilm-forming ability, antimicrobial resistance, and biofilm cell survival/recovery growth after sanitizer exposure. A total of 28 Salmonella serovars was identified within these strains. The most common serovars identified were Anatum, Dublin, Montevideo, and Typhimurium, with these accounting for nearly half of the total strains. The vast majority (86%) of the strains was able to develop strong biofilms, and the biofilm-forming ability was highly strain dependent and related to cell surface expression of extracellular polymeric structures. These strains also demonstrated strong tolerance to quaternary ammonium chloride (QAC) and chlorine dioxide (ClO2), but were more sensitive to chlorine treatment. Sanitizer tolerance and bacterial postsanitization recovery growth were closely associated with strains' biofilm-forming ability. Thirty percent of the examined strains were found resistant to multiple antimicrobial agents and the resistance phenotypes were serovar associated, but not related to strains' biofilm-forming ability. Pulsed-field gel electrophoresis analysis tended to group strains by serovar rather than by biofilm-forming ability. Collectively, these data indicate that the strong biofilm formers of certain S. enterica strains/serovars possess significant potential for causing meat product contamination in meat processing environment.
Introduction
I
Bosilevac et al. (2007) compared imported and domestic (U.S.) beef trim and also found that Salmonella prevalence was less than 1% in all samples, and serovars Anatum, Montevideo, and Enteritidis were identified from the U.S. trim samples. In addition, their surveys of retail and commercial ground beef samples showed that Montevideo and Anatum were more frequently isolated from contaminated meat products compared with other Salmonella serovars (Bosilevac et al., 2009). These observations suggest that certain Salmonella serovars might be better adapted to the meat processing environment, thus, causing product contamination in commercial plants.
While hide to carcass transfer during processing is an established contamination route in meat plants, other pathogen sources involving bacterial biofilm formation have been noted to exist in commercial establishments (Marouani-Gadri et al., 2009; Yang et al., 2015). Biofilm formation is an important strategy for bacteria survival because biofilm cells are more tolerant to physical/chemical treatments, including sanitization. When viable bacteria in biofilms become detached as the meat products pass over the area, occasions of meat product crosscontamination could occur. The potential contribution of biofilm formation by foodborne pathogens, such as E. coli O157:H7, to meat contamination has been reported (Wang et al., 2014a, 2016; Visvalingam et al., 2016). Meanwhile, the impact of Salmonella biofilms and its environmental persistence on food safety has been investigated as well (Ben Abdallah et al., 2014; Chousalkar and Gole, 2016; Gong and Jiang, 2017).
Thus, one potential contamination route of S. enterica at meat plants could be that certain Salmonella serovars or strains are better adapted to the meat processing environment through biofilm formation, survive the sanitization and daily cleaning procedures, therefore cause product crosscontamination. To test this hypothesis, we characterized a group of S. enterica strains isolated from contaminated beef trim at commercial processing plants in the United States for their biofilm-forming ability, antimicrobial resistance (AMR) profile, and biofilm cell survival/recovery growth after treatments by common sanitizers. Furthermore, pulsed-field gel electrophoresis (PFGE) genotyping analysis was performed in an attempt to identify any genomic patterns that might be associated with high biofilm-forming ability and sanitizer tolerance.
Materials and Methods
Bacterial strains, growth conditions, and curli/cellulose expression
Eighty-nine S. enterica strains were obtained from FSIS for this study. This diverse strain collection was isolated from multiple incidences at commercial plants located in geographically distant states/regions in the United States and harvested at various time points, in an effort to represent the breadth of Salmonella strains/serovars causing meat contamination under the current commercial settings. Among these, 60 strains were isolated from beef trim in 2014 from various beef processing plants located in 21 different states in the United States. Another 29 strains were isolated from 422 trim enrichment broth samples that were collected from various meat plants located in all 50 U.S. states and the country's territories/insular areas from 2012 to 2014 (Table 1).
Antimicrobial abbreviations: Am, amoxicillin–clavulanic acid; Ap, ampicillin; C, chloramphenicol; S, streptomycin; Su, sulfisoxazole; T, tetracycline; F, cefoxitin; Cf, ceftiofur; Ax, ceftriaxone; G, gentamicin; K, kanamycin; SxT, sulfamethoxazole–trimethoprim; Cp, ciprofloxacin; N, nalidixic acid; and Ai, azithromycin.
All strains were stored and cultured, as described previously (Wang et al., 2013, 2014b). The expressions of two bacterial extracellular polymeric structures (EPS) associated with biofilm-forming abilities and sanitizer tolerance, curli fimbriae and cellulose, were tested as previously described (Wang et al., 2013) using Congo Red Indicator plates and Luria broth agar plates containing Calcofluor dye, respectively.
Sanitizers
Three types of commercial sanitizers commonly used in the meat industry were applied in this study for the sanitizer tolerance experiments. Quaternary ammonium chloride (QAC) (Vanquish™; Dawn Chemical Corp., Milwaukee, WI), chlorine (Clorox, Oakland, CA), and chlorine dioxide (ClO2) (Pro-Oxine™; DanMar, Arlington, TX) were prepared as previously described (Wang et al., 2014a) and applied at 300, 200, and 50 parts per million (ppm), respectively, following the manufacturer's instructions.
Biofilm formation and biofilm cell postsanitization recovery growth
Biofilm formation by each individual strain was measured on 96-well polystyrene plates at 22°C to 25°C for 3 days as previously described with Crystal Violet staining and absorbance measurement (A 570nm) (Wang et al., 2012).
Biofilm sanitizer tolerance was tested by measuring bacterial recovery growth after each treatment using a 96-well plate absorbance assay as described previously (Wang et al., 2014) with one slight modification that the wavelength for measuring the absorbance of medium (Dey/Engley broth; Beckton Dickinson) color shift was set at 405/590 nm.
Biofilm formation on materials commonly used in the meat industry
Biofilms were developed on stainless steel (SS, 18 × 18 × 2 mm) or polyvinyl chloride (PVC, 14 × 12 × 3 mm) plastic chips by incubating the chips for 5 days at 7°C to simulate the fabrication room condition at meat plants, then biofilm cells were harvested and quantified (log10 CFU/cm2) as previously described (Wang et al., 2013).
Antimicrobial susceptibility test
AMR phenotype was determined based on the minimal inhibitory concentration (MIC) of the examined strains for each of the 15 common antimicrobial agents, measured with the Sensititre broth microdilution system and CMV2AGNF plates (TREK Diagnostic Systems, Cleveland, OH) as previously described (Schmidt et al., 2015; Wang et al., 2016).
Pulsed-field gel electrophoresis
All strains were analyzed using the PFGE separation of XbaI-digested genomic DNA method and banding patterns were analyzed and comparisons made as previously described (Arthur et al., 2008).
Statistical analysis
Data comparison of biofilm formation on 96-well plates was performed with a one-way analysis of variance (ANOVA) followed by Dunnett's multiple comparison to a single control group. Bacterial counts recovered from SS or PVC chips were log transformed and analyzed with a one-way ANOVA and Dunnett's multiple comparison posttest. To analyze the potential contribution of biofilm-forming ability to bacterial postsanitization recovery growth, strains were categorized, based on their A 570nm values, into “Weak” (A 570nm 0.5–1.0), “Moderate” (A 570nm 1.0–2.0), “Strong” (A 570nm 2.0–3.0), or “Superior” (A 570nm 3.0–4.0) biofilm former groups. Data of bacterial postsanitization survivability were analyzed using the PROC GLIMMIX procedure of SAS with a model that included biofilm-forming ability, sanitizer, and their interaction. Strain nested within biofilm type was included as a random effect. Least squares mean was generated for the biofilm-forming ability × sanitizer interaction and separated using the diff option. p Values less than 0.05 were considered statistically significant.
Results
Serovars classification and EPS expression
Twenty-eight different Salmonella serovars were identified within the examined strains using commercial antisera (Remel, Lenexa, KS). Two strains were deemed nontypeable. Of the 89 strains, 80 (89.9%) and 57 (64%) strains tested positive at various levels for curli fimbriae and cellulose expression, respectively. All 57 cellulose-positive strains also exhibited positive curli expression (Table 1).
Biofilm-forming ability
All strains developed biofilms on 96-well polystyrene plates at various levels higher than the background control (A 570nm 0.25–0.45). Notably, the majority of these strains exhibited high potency of biofilm formation as 64 (72%) and 13 (14%) strains developed Superior and Strong biofilms, respectively, whereas only 7 (8%) and 5 (6%) strains belonged to the Weak and Moderate biofilm groups, respectively. The biofilm-forming ability was highly strain specific and closely related to the positive expressions of curli and/or cellulose fimbriae (Table 1).
On SS and PVC surfaces under fabrication conditions (7°C), significant amount of bacterial colonization ranging from 2.7 to 5.9 log10 CFU/cm2 was detected by the examined strains, except S. Dublin strain FSIS1500564, whose numerable bacterial counts were lower than the limit of detection of the assay (1.5 log10 CFU/cm2), likely due to the insufficient cell replication under the low temperature. On both materials, approximately half of the strains exhibited biofilm cell density higher than 5.0 log10 CFU/cm2 (Table 2), and there was no significant difference in the amount of biofilm cells recovered from PVC and SS surfaces. The potency of biofilm formation on these materials also was highly strain dependent. Overall, strains that developed high biofilm masses on 96-well plates at room temperature also exhibited strong biofilm-forming potency on the chip surfaces at 7°C, which corresponded to the positive EPS expression status.
PVC, polyvinyl chloride; SS, stainless steel.
Postsanitization recovery growth of biofilm cells
All sterile PBS-treated positive control samples exhibited fully recovered growth that yielded average A 405/590nm values between 1.25 and 1.32 regardless of strains' biofilm-forming ability. Negative control samples that contained sterile Dey/Engley broth only yielded average A 405/590nm values lower than 0.50. Based on the absorbance measurements, the examined strains were classified into four groups: “No growth” (A 405/590nm <0.50), “Weak growth” (A 405/590nm 0.50–1.10), “Moderate growth” (A 405/590nm 1.10–1.25), and “Full Growth” (A 405/590nm >1.25).
The Salmonella strains overall demonstrated high tolerance to QAC and ClO2 treatments as most of the strains exhibited full or moderate recovery growth (Table 3); however, they were more sensitive to chlorine treatment as most of the examined strains exhibited no apparent recovery growth (43 strains, 48% of total strains) or weak recovery growth (31 strains, 35% of total strains) after the treatment. Notably, when the mean of A 405/590nm values were calculated for each biofilm potency group, statistical analysis indicated that sanitizer tolerance and recovery growth differed across the biofilm potency groups. The Superior and Strong biofilm former groups exhibited significantly (p < 0.05) higher postsanitization recovery growth compared with the Moderate or Weak groups (Fig. 1).
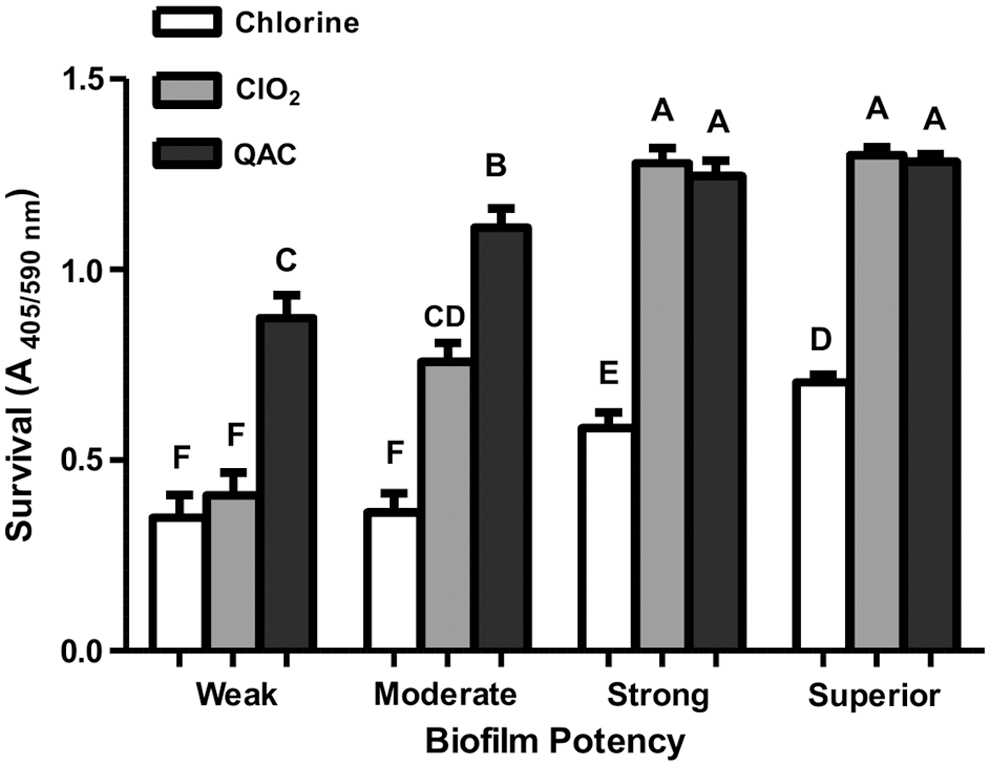
Postsanitization recovery growth of Salmonella strains isolated from beef trim, classified by their potency of biofilm formation on 96-well polystyrene plates. Error bars represent mean ± standard deviation. Columns labeled by the same letters are not statistically different (p > 0.05).
QAC, quaternary ammonium chloride.
PFGE genotyping of the strains
XbaI PFGE genotyping of the Salmonella strains produced eight distinct clusters (Fig. 2). The clusters tended to group strains by their serovars, rather than biofilm-forming ability or isolate source (Fig. 2). Notably, the biofilm-forming ability did vary within serovar even for strains with indistinguishable PFGE patterns. Strains of serovar Dublin were the most homogeneous by PFGE analysis and these strains were mostly weak or moderate biofilm formers.

XbaI restriction digest patterns and biofilm-forming ability of the Salmonella strains isolated from beef trim.
AMR profile
Table 1 shows the resistance profiles of the examined strains against 15 common antimicrobial agents. AMR was observed in 27 strains (30%) and most of these AMR strains were resistant to multiple antimicrobial agents, except four strains that were resistant to tetracycline only. The predominant AMR Salmonella serovars observed herein were Dublin (12 AMR strains) and Typhimurium (5 AMR strains), whereas the only two Newport strains in this study were both resistant to multiple antibiotics. Serovars of Anatum, Derby, Johannesburg, Kentucky, Meleagridis, Montevideo, Muenchen, and Muenster each contained one AMR strain.
The antimicrobial agents with the highest incidence of resistance included tetracycline (n = 26), sulfisoxazole (n = 22), streptomycin (n = 21), chloramphenicol (n = 18), and ampicillin (n = 16). Resistance to gentamicin and nalidixic acid was observed much less frequently, with only one and two strains showing resistance to these two antimicrobial agents, respectively. Amikacin and ciprofloxacin resistance was not detected in this study. The most common resistant pattern was resistance to nine antimicrobials (AmApFCfAxCSSuT). Eight AMR strains of serovars, Dublin, Newport, and Typhimurium, demonstrated this resistance pattern. Other resistance patterns observed in multiple strains included CSSuT (three strains) and SSuT (two strains).
Discussion
To investigate whether certain Salmonella serovars/strains might be better adapted to meat processing environment through biofilm formation/sanitizer tolerance and play roles in product contamination, we characterized 89 Salmonella strains isolated from beef trim for their biofilm-forming ability, sanitizer tolerance, and AMR profiles. A wide variety of serovars (n = 28) were identified within the examined strains, and eight of them (Agona, Berta, Infantis, Montevideo, Muenchen, Newport, Oranienburg, and Typhimurium) were among the top 20 most frequently isolated Salmonella serovars that have caused human infections as reported by the Centers for Disease Control and Prevention (CDC, 2016). The four major serovars found in the present study were Anatum (n = 7), Dublin (n = 13), Montevideo (n = 11), and Typhimurium (n = 9), which was in part consistent with a number of previous studies that found Anatum and Montevideo were among the Salmonella serovars frequently isolated from commercial ground beef samples/retail meat products (Bosilevac et al., 2009) and beef trim samples (Bosilevac et al., 2007; USDA FSIS, 2011).
Anatum and Montevideo also were two out of the three dominant Salmonella serovars that were found entering beef processing environment, which might also contribute to their higher likelihood of causing product contamination (Brichta-Harhay et al., 2011). The same study also found that the postintervention carcass contaminations by S. enterica on two separate occasions were caused by S. Dublin strains in one event and by S. Typhimurium strains in the other. Moreover, Doerscher et al. (2015) analyzed beef products provided by USDA for the National School Lunch Program from 2011 to 2014, and their serovar data showed that the 114 Dublin strains (48.93%) and the 23 Typhimurium strains (9.87%) were the two most commonly observed Salmonella serovars among the 23 serovars identified in boneless beef, and Dublin (15 strains, 37.5%) was also the most commonly identified serovar within the 15 Salmonella serovars found in ground beef.
These above findings were in line with our current observation that Dublin and Typhimurium were among the major Salmonella serovars isolated from beef trim, suggesting that certain Salmonella serovars might be more capable of surviving and causing product contamination in meat plants.
The majority of the examined strains (77/89, 87%) exhibited Superior or Strong biofilm-forming ability on 96-well polystyrene plates. Notably, the 11 Montevideo strains, the Salmonella serovar frequently isolated from contaminated beef products, all exhibited Superior (9 strains) or Strong (2 strains) biofilm-forming ability. Moreover, all the examined Newport and Typhimurium strains also exhibited Superior or Strong biofilm-forming ability.
Serovars Newport and Typhimurium have been reported by numerous studies (Clegg et al., 1983; Baloda et al., 2001; Berge et al., 2004; You et al., 2006) to have the capability of spreading from one environment to the other by contaminated feed sources or by manure or agricultural waste runoff, and also have the strong survival ability in manure and terrestrial environment for extended periods of time (up to 9 months). Our current observations that all the Montevideo, Newport, and Typhimurium strains isolated from beef trim had strong biofilm-forming ability suggest that biofilm formation might be one of the critical strategies that these serovars employ to survive under extreme conditions, contaminate the environment and, in this case, possibly disseminate within beef processing plants and result in meat product contamination.
The absorbance assay for testing Salmonella postsanitization survival/recovery growth was based on a positive correlation between bacterial cell densities and medium absorbance values, since the medium color shift of the Dey/Engley broth from purple to yellow as a result of carbohydrate fermentation from bacterial growth could be quantified by absorbance measurement set at 405/590 nm. Using this assay we evaluated the effectiveness of the common sanitizers against Salmonella biofilms (Table 3). QAC and ClO2 at recommended concentrations were not effective enough in eliminating Superior and Strong biofilms, which was in agreement with previous findings that common sanitizers, such as sodium hypochlorite, sodium hydroxide, and benzalkonium chloride, were not able to eradicate mature Salmonella biofilms on food contact surfaces (Joseph et al., 2001; Corcoran et al., 2014).
We also measured bacterial susceptibility (MIC) to the sanitizers; however, in most cases, sanitizer susceptibility was not correlated to bacterial postsanitization survival/recovery growth (data not shown), thus, the strong sanitization tolerance and recovery growth capability were most likely associated with each strain's biofilm-forming ability.
A relatively high prevalence (30%) of AMR was detected within the examined strains. Similar results were obtained from a previous study (Brichta-Harhay et al., 2011) that screened Salmonella strains isolated from 3040 postintervention carcasses for AMR and, among the 24 isolated strains, 10 (42%) were identified as AMR strains. Logue et al. (2003) also reported that multiple AMR, including tetracycline, sulfamethoxazole, and streptomycin, were commonly observed in Salmonella strains isolated from retail meats.
Importantly, the observed AMR phenotypes appeared to be serovar associated. The dominant AMR serovar was Dublin (12 out of 13 Dublin strains) followed by Typhimurium (5 out of 9 Typhimurium strains). These two serovars collectively made up 63% of all the AMR strains identified. Meanwhile, the two Newport strains in this study were both AMR strains. Similar observations of serovar-associated Salmonella AMR phenotypes have been reported previously. For example, the abovementioned 10 AMR Salmonella strains from previous study (Brichta-Harhay et al., 2011) that isolated from postintervention carcasses also included serovars of Typhimurium, Dublin, Newport, and Reading, with Typhimurium (6 out of 10 strains) being the dominant AMR serovar. Moreover, the same study also showed that within the 16,218 Salmonella cattle hide and carcass isolates, the most commonly observed AMR serovars were Typhimurium, Newport, and Uganda.
Notably, we showed that the biofilm-forming ability was positively correlated to strains' sanitizer tolerance; however, the association between biofilm formation and strains' AMR phenotype was not detected in this study (data not shown) even though such association has been reported previously (Kim and Wei, 2007; Gong et al., 2013; Emami et al., 2015). Conversely, previous studies have shown that bacterial AMR strains were not more tolerant to sanitizers than their antibiotic-sensitive counterparts (Anderson et al., 1997; Karatzas et al., 2008).
However, prolonged exposure of Salmonella strains to sanitizers at sublethal concentrations could lead to selection of strains with reduced antibiotic susceptibility or even resistance to multiple antibiotics (Karatzas et al., 2007; Condell et al., 2012; Futoma-Kołoch et al., 2015), in part, due to the fact that such exposure may potentially select for strains with overexpressed efflux pumps. Interestingly, serovar-related variation of crossresistance between sanitizers and antimicrobial agents was reported previously (Braoudaki and Hilton, 2004). Thus, the effect of sanitizer sublethal exposure on these Salmonella strains with respect to antibiotic resistance or antibiotic/sanitizer crossresistance requires further investigation.
PFGE analysis has been utilized for strain tracking in foodborne outbreaks and to identify genotypic clustering for phenotypic markers. In the present study, it was utilized in an attempt to identify strain-specific factors associated with biofilm formation. However, the clustering by serovars was observed much more than clustering by biofilm-forming ability. PFGE clustering by Salmonella serovars has been reported previously (Bosilevac et al., 2009; Brichta-Harhay et al., 2011). It should be noted that the biofilm-forming ability also varied somewhat by serovars in the present study, as most of the Dublin strains were Moderate or Weak biofilm formers, whereas all the Montevideo, Typhimurium, and Newport strains exhibited Superior or Strong biofilm-forming ability. However, to obtain a more detailed analysis of within-serovar variation in genotype and biofilm-forming ability, more strains of each serovar will need to be analyzed.
Conclusions
The majority of the S. enterica strains isolated from beef trim were able to form strong biofilms on contact surfaces under meat processing conditions. The biofilm-forming ability was strain dependent and at certain level, serovar associated. The strong biofilm formers exhibited high tolerance to common sanitizers. A relatively high prevalence (30%) of AMR was identified within these strains, which was not associated with their biofilm-forming ability or sanitizer tolerance. Collectively these data suggest that in-house biofilm formation by certain S. enterica strains or serovars, and the resultant high sanitizer tolerance might play an important role in beef product contamination at commercial plants.
Footnotes
Acknowledgments
The authors would like to thank Dr. John Johnston and his colleagues at FSIS for providing the Salmonella strains for this study. The authors would also like to thank Alberto O. Alvarado, Bruce Jasch, and Sydney Brodrick for technical support, and Jody Gallagher for secretarial assistance. This study was supported by research funding from the Beef Checkoff through the North American Meat Institute Grant No. 58-3040-5-003. Names are necessary to report factually on available data; however, the USDA neither guarantees nor warrants the standard of the product, and the use of the name by USDA implies no approval of the product to the exclusion of others that may also be suitable.
Disclosure Statement
No competing financial interests exist.