
Abstract
Salmonella is a predominant foodborne pathogen in the United States and other countries. Mobile genetic elements such as plasmids allow Salmonella to adapt to external stress factors such as nutrient deprivation and host factors. Incompatibility group I1 (IncI1) plasmid-carrying Salmonella enterica strains were examined to determine the presence of plasmid-associated genes and their influence on phenotypic characteristics. The objective of this study was to understand the genetic determinants on IncI1 plasmids and their impact on antimicrobial susceptibility, competitive growth inhibition of Escherichia coli, and plasmid transfer. Primers were designed for genes that play a role in virulence, antimicrobial resistance, and plasmid transfer based on previously sequenced IncI1 plasmids. Polymerase chain reaction assays were conducted on 92 incompatibility group I1 (IncI1)-positive S. enterica strains. Phenotypic characterization included conjugation assays, antimicrobial susceptibility testing, and bacteriocin production based on the inhibition of growth of colicin-negative E. coli J53. The antimicrobial resistance genes aadA1, tetA, sul1, and bla CMY were detected in 88%, 87%, 80%, and 48% of the strains, respectively. Over half of the strains were resistant or intermediately resistant to streptomycin (85%), sulfonamides (76%), tetracycline (74%), and ampicillin (68%) and 57% of the strains inhibited growth of E. coli J53 strain. Among putative virulence genes, colicin-associated colI and cib were detected in 23% and 35% of strains and imm and ccdA were present in 58% and 54% of strains, respectively. Approximately 61% of strains contained plasmids that conjugally transferred antimicrobial resistance, including 83% where the recipient received IncI1 plasmids. Most of the strains carried an assortment of transfer associated (pil and tra) genes with between 63% and 99% of strains being positive for individual genes. Taken together the study affirms that IncI1 plasmids likely play roles in the dissemination of antimicrobial resistance and virulence-associated factors among enteric organisms.
Introduction
S
Plasmids can replicate independently of the bacterial chromosome (Carattoli, 2011) and have evolved to potentially acquire genes for antimicrobial resistance, adhesion, and virulence among other characteristics (Martinez and Bauero, 2002). Plasmids are often defined by their incompatibility (Inc) types and diverse plasmid types are known to harbor and spread antimicrobial resistance genes (Cambray et al., 2010; Lai et al., 2013; Sanad et al., 2016). Among them, IncI1 plasmids, which typically range from 90 to 110 kb, have been noted for being associated with dissemination of ceftriaxone resistance among Salmonella strains (Smith et al., 2015).
The ability of IncI1 plasmid to carry and spread extended spectrum cephalosporin (ESC) resistance genes offers a potential explanation for the plasmids' prevalence among multidrug-resistant Salmonella (Folster et al., 2016). IncI1 plasmids are known for their potential to disseminate among other enteric pathogens (Wong et al., 2016). IncI1 plasmids are known to encode two distinctive types of pili, thin flexible (pil locus) and thick rigid (tra locus), in their conjugative transfer region; along with multiple inversion regions called shufflons and sog, which suppresses mutations (Sampei et al., 2010). These characteristics are unique compared to other plasmid types, such as IncF, which possess a smaller transfer region than IncI1 (Komano et al., 2000).
Salmonella require some metabolic resources (i.e., fitness cost) to retain IncI1 plasmids (Martin et al., 2016). Smith et al. (2015) found that IncI1 plasmids typically carry genes responsible for partitioning and host-addiction systems. These addiction genes (e.g., ccdAB) cause the bacterium to retain the plasmid after cell division, facilitating successful inheritance and maintenance of the plasmid (Doumith et al., 2012).
Successful plasmid-gene combination such as IncI1 and bla CMY-2, are prevalent among certain enteric bacteria (Mnif et al., 2010); however, there remains a need to understand the diversity and function of IncI1 plasmids. The major objectives of this study were to detect IncI1-associated antimicrobial resistance, putative virulence, and transfer-related genes present in IncI1-positive Salmonella strains and determine their impact on Salmonella phenotypes. Experiments were conducted on isolates from diverse sources, including poultry, cattle, swine, and human patients, to determine their phenotypic and genotypic characteristics and the capability of these plasmids to undergo horizontal transfer.
Materials and Methods
Bacterial strains
Ninety-two Salmonella enterica isolates previously identified as carrying IncI1 plasmids were selected for the study (Kaldhone et al., 2008; Lynne et al., 2008, 2009; Melendez et al., 2010; Han et al., 2011; Marrero-Ortiz et al., 2012). Strains originated in the United States and were collected from chicken (29%, 27/92), bovine (29%, 26/92), turkey (17%, 15/92), porcine (14%, 13/92), and human (12%, 11/92) related sources between the years 1992–2009. The serovar distribution included Heidelberg (40%, 37/92), Typhimurium (23%, 22/92), Kentucky (18%, 16/92), Newport (9%, 8/92), and others (10%, 9/92; Fig. 1). Strains were frozen at −80°C in Brain Heart Infusion broth with 20% glycerol for archival storage. For use in experiments, strains were streaked on tryptic soy agar containing 5% sheep's blood (blood agar) and incubated at 37°C overnight.

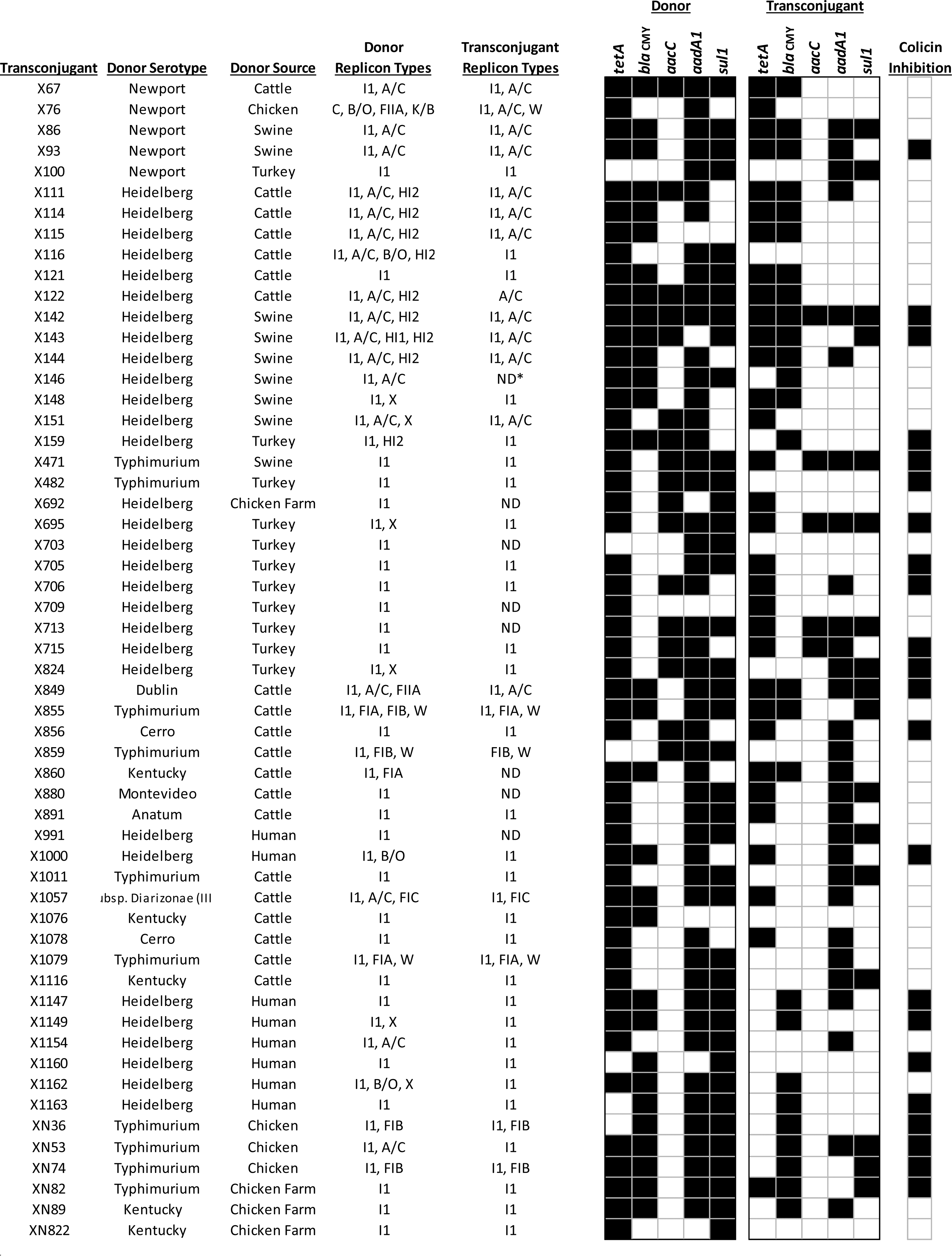
Isolate information and results of antimicrobial resistance, virulence, and transfer phenotypic and polymerase chain reaction-based testing. For antimicrobial susceptibility testing results, a white box is susceptide, a grey box is intermediate and a black box is resistant. A black box indicates a PCR positive result in PCR screening experiments. *MW: midwestern state, not further defined. **ND: not determined.

Conjugation experiments
Salmonella strains served as potential donors and the sodium azide-resistant Escherichia coli J53 strain was the recipient for conjugation (Jacoby and Han, 1996). Initial experiments were conducted using a cross-streaking method. Donors and the recipient were streaked across one another on Luria-Bertani (LB) broth agar plate and incubated at 37°C for 18 h. At the point of intersection, the equivalent of three to four colonies were picked and streaked onto LB agar plates containing ampicillin (32 μg/mL), gentamicin (10 μg/mL), or streptomycin (32 μg/mL) and sodium azide (350 μg/mL). These selection plates were incubated for up to 48 h to identify likely transconjugants. Those combinations that did not yield transconjugants were subjected to a broth-mating method (Zheng et al., 2016) at either the ratio of 1:9 (donor: recipient) or 1:1, if the 1:9 ratio was unsuccessful. In each case, 100 μL of the suspension was spread on LB agar selection plates as described above to identify transconjugants.
Polymerase chain reaction for replicon types, antimicrobial resistance, virulence and transfer-associated genes
DNA template was prepared using a boiling method in which three to four bacterial colonies were picked from the blood agar plate, suspended in 200 μL of sterile water, and boiled to lyse the cells (Wang et al., 2015). Polymerase chain reactions (PCRs) were conducted to confirm the presence of the IncI1 replicon (Carattoli et al., 2005) and for potential IncI1-associated antimicrobial resistance (sul1, aadA1, aacC, tetA, and bla
CMY), putative virulence (ccdA, ccdB, col1, cib, imm, copD, and cusA), and plasmid transfer-associated genes (pilJ, pilM, pilP, pilS, traG, traL, traQ, and traT) (Supplementary Table S1; Supplementary Data are available online at
Antimicrobial susceptibility testing
Strains were previously tested for antimicrobial susceptibility using either broth microdilution or disc diffusion following the Clinical and Laboratory Standards Institute (CLSI) guidelines (Foley et al., 2006; Kaldhone et al., 2008; Lynne et al., 2008, 2009; Melendez et al., 2010; Han et al., 2011; Marrero-Ortiz et al., 2012). Antimicrobial susceptibility testing (AST) data from each study were compiled and across the different studies and all the strains were tested for susceptibility to ampicillin (Amp), ceftriaxone (Axo), ciprofloxacin (Cip), chloramphenicol (Chl), gentamicin (Gen), kanamycin (Kan), nalidixic acid (Nal), streptomycin (Str), sulfisoxazole (Sul), trimethoprium-sulfamethoxazole (Sxt), and tetracycline (Tet). To verify the earlier results, a subset of the strains (n = 35) were retested using disc diffusion testing for Amp, Axo, Chl, Gen, Str, Sul, Sxt, and Tet, using E. coli ATCC 25922 as the quality control strain (CLSI, 2008).
Colicin inhibition assay
The ability of Salmonella and transconjugant strains to produce colicin was evaluated by assessing growth inhibition of E. coli J53. J53 was suspended in sterile water and its bacterial cell concentration adjusted to a 0.5 McFarland standard. The suspension was swabbed for confluence on blood agar plates and 10 μL of a Salmonella suspension, prepared by suspending two bacterial colonies in 100 μL of sterile water, was spotted on the J53 lawn. The plates were incubated at 37°C for 16 to 18 h and then examined for growth inhibition of J53 adjacent to the Salmonella growth.
Results
PCR assays were conducted to detect the presence of genes previously identified as being associated with IncI1 plasmids and potentially responsible for antimicrobial resistance, conjugal transfer, and virulence properties. Positive PCR results were identified for aadA1 in 88% (81/92) of strains, followed by tetA (87%, 80/92), sul1 (80%, 74/92), bla CMY (48%, 44/92), and aacC (30%, 28/92) (Fig. 1). For the corresponding resistance phenotypes, 78% (78/92) of the Salmonella strains were resistant or intermediately susceptible to Str (85%, 78/92), followed by Tet (74%, 68/92), Sul (76%, 69/92), Amp (68%, 62/92), Axo 43% (39/92), and Gen (35%, 33/92; Fig. 1). Only three (3%) strains were susceptible to all antimicrobials examined in comparison.
Over half of the strains carried the putative virulence genes imm (58%, 53/92) and ccdA (53%, 49/92), while copD (37%, 34/92), cib (35%, 32/92), cusA (33%, 30/92), and col1 (23%, 21/92) were identified in fewer strains (Fig. 1). When present, the copper resistance genes, copD and cusA, were often detected (n = 25) together. col1 and cib are two of several known bacteriocin (colicin) encoding genes and imm is the host immunity gene for colicin's effects. Over half of the strains (57%, 53/92) were positive for colicin production and inhibited J53 growth (Fig. 1). A positive test was defined as having a clear zone of growth inhibition of the bacterial lawn adjacent to the Salmonella spotted on the plate (Supplementary Fig. S1).
Among the conjugation-related genes, pilM (98%, 91/92), pilP (91%, 84/92), and pilJ (82%, 76/92) occurred in most of the strains as did traL (96%, 90/92), traQ (95%, 89/92), and traG (84%, 77/92) (Fig. 1). Over half of the strains (60%, 56/92) were able to generate transconjugants. Five isolates were susceptible to each of the selection antimicrobials and as such the plasmid transfer was not determined (marked as “ND” in the conjugation results in Fig. 1). Some transconjugants (74%, 41/56) were obtained by the plate mating method, while the remaining 15 were obtained by the broth mating method at a 1:9 proportion (n = 8) or 1:1 proportion (n = 7). When the presence of the IncI1 plasmid in the transconjugants was evaluated, the majority (82%, 46/56) carried IncI1 plasmids, while the remaining strains contained other resistance plasmids (Fig. 2). PCR for the resistance genes in the transconjugants indicated that 59% (33/56) of the strains were positive for tetA, followed by aadA1 (52%, 29/56), bla CMY-2 (46%, 27/56), sul1 (30%, 17/56), and aacC (9%, 5/56) (Fig. 2). In addition, 29% (22/56) of the transconjugants could produce bacteriocins that inhibited the growth of J53 (Fig. 2).
Evaluation of transconjugants generated in the study. A black box indicates a positive result in the test. *ND indicates no plasmid type was detected by PCR screening.
Discussion
IncI1 plasmids have often been associated with the carriage of antimicrobial resistance genes. Less is known about the potential contribution of IncI1 plasmids to virulence in Salmonella. This study was undertaken to assess their potential contribution to virulence through putative virulence gene detection and their production of bacteriocins, which may inhibit bacterial competition for colonization; evaluate their role in antimicrobial resistance; and determine whether the plasmids can be transferred conjugally. Many earlier studies of IncI1 plasmids focused on ESC resistance (Smith et al., 2015; Folster et al., 2016; Liakopoulos et al., 2016). This article examined several additional resistance genes that have been reported to be present in IncI1 plasmids in Salmonella including, aadA1, aacC, bla CMY-2, sul1, and tetA (Fricke et al., 2009; Han et al., 2012).
When a resistance gene was detected, the corresponding resistance phenotype was typically observed, with a few noted exceptions. A relatively high percentage of susceptible strains were positive for tetA (n = 18) and sul1 (n = 17) even after repeated PCR testing. When the AST for several of the isolates was repeated they were still identified as susceptible to Tet and Sul. The disparity between presence of sul1 and lack of Sul resistance was reported in previous studies (Awad et al., 2016). tetA encodes an efflux pump that is regulated by the tetR gene product (Chopra and Roberts, 2001).
The disparity between genotype and phenotype could be due to mutations that reduce or abolish the function of corresponding proteins or expression of resistance genes. Both sul1 and tetA were detected in isolates that originated from multiple sources and time periods in the current study, thus highlighting their widespread distribution in Salmonella from different food animal populations. Tet resistance genes have been identified on a wide range of transmissible resistance plasmids, indicating their importance due to their distribution among bacteria and potential for horizontal gene transfer.
Another resistance gene with relatively high prevalence was aadA1. aadA1 was detected in 78 strains, 71 (88%) of which displayed resistance or intermediate susceptibility to Str. bla CMY-2 is associated with resistance to multiple β-lactam antimicrobials, including Axo, a drug used for the treatment of severe Salmonella infections. In this study, 87% of strains resistant to Axo were positive for the bla CMY-2 gene. bla CMY-2 is known to be distributed among several IncI1 and IncA/C plasmids (Welch et al., 2007; Fricke et al., 2009). Liakopoulos et al. (2016) concluded that emergence of ESC-resistant Salmonella in The Netherlands was due to the presence of bla CMY gene on IncI1 plasmids, potentially originating from imported poultry from South America. This example highlights the global concern of ESC-resistant Salmonella and the potential for their widespread dissemination.
The ability to transfer antimicrobial resistance provides the capability of pathogens to maintain their presence in animal production areas (Anacarso et al., 2016; Aviv et al., 2016). A major concern is that food animals could become a reservoir of these resistant bacteria and subsequent infections would lead to illnesses that would be difficult to treat with conventional antibiotics (Aviv et al., 2016). Most of the strains in the current study were positive for tra and pil genes associated with the IncI1 conjugation machinery (Fig. 1) and more than half produced transconjugants when mated with sodium azide-resistant E. coli J53. The majority (82%) of the transconjugants generated carried the IncI1 plasmid transferred from the donor. When the transconjugants were examined for the resistance genes, it was observed that not all of the resistance genes were able to be transferred (Fig. 2), which may be due to genes being on a plasmid that was not transferred or located on the chromosome. In several cases it was difficult to determine whether the resistance genes were on the IncI1 plasmids or another plasmid, such IncA/C plasmids that often carry multiple resistance genes. Twelve of the 46 IncI1-positive transconjugants contained IncA/C plasmids, thus the plasmids often co-transfer. In 24 cases, the transconjugants appeared to only receive an IncI1 plasmid and in these cases there was a diversity of resistance genes transferred to the recipient (Fig. 2), including bla CMY-2, aadA1, aacC, sul1, and tetA.
To further evaluate the genetics of IncI1-positive strains, nine of the isolates (142, 143, 144, 146, 397, 991, 1148, N36, and N89) underwent DNA sequencing analyses as parts of parallel studies (Kaldhone et al., 2017; Khajanchi et al., 2017). The results of the sequencing indicated a diversity of resistance genes on the IncI1 and other plasmids within the strains. For example, both isolate 142 from swine, and 1148 from a human patient, had large contigs containing the IncI1 replicon-associated sequence and aadA1, aacC, and sul1. Isolate 142 contained additional resistance genes that are likely associated with different plasmid types, while 1148 did not (Kaldhone et al., 2017). With isolate N36, the IncI1 replicon contained the bla CMY-2 gene. Many other strains had multiple plasmid DNA sequence contigs, which were difficult to assign the specific resistance genes to a particular plasmid type.
Initial attempts to evaluate the conjugative ability of plasmids were conducted on solid plate media using a cross streaking method. A phenotype detected for several donor–recipient interactions was the apparent inhibition of the E. coli recipient strain by the donor. Many of the available sequences for IncI1 plasmids indicate the presence of the colicin genes, such as cib or col1 and host immunity gene imm (Smith et al., 2015). Thirty-two strains were positive for cib, of these, 25 could inhibit E. coli (Fig. 1). In addition, 21 strains were positive for col1, and 12 displayed growth inhibition of E. coli.
Nedialkova et al. (2014) used a mouse model to examine production of colicin by Salmonella Typhimurium and demonstrated that colicin could inhibit growth of other enteric bacteria under gut inflammatory conditions (Nedialkova et al., 2014). Avirulent strains that did not produce colicin did not exhibit a competitive advantage. In the current study, the Salmonella serovars associated with growth inhibition included Newport, Heidelberg, Typhimurium, and Kentucky. Of the strains from different sources, 10 of the 11 (91%) S. Heidelberg isolated from human patients demonstrated an ability to inhibit E. coli growth. Interestingly, a relatively low percentage (8%, 2/25) of the isolates from cattle, regardless of serotype, demonstrated the inhibitory phenotype, which may indicate that the production of colicins may have a selective advantage in other hosts, such as poultry and swine.
Just under half (26/57) of the strains that demonstrated growth inhibition had plasmids that could be transferred using the methods employed in this study. The broth-culture mating was used to attempt to dilute the potential colicin effect; however, even in the broth culture, the presence of colicin may limit conjugation. This phenomenon warrants further exploration to evaluate the effect of colicin on conjugation. Future studies may need to be done with recipient strains that are more resistant to colicin inhibition. Interestingly, of the colicin-positive strains that could transfer plasmids, 22 transconjugants also demonstrated the inhibitory phenotype (Fig. 2), this includes 13 strains that appeared to only an IncI1 plasmid.
There were also several instances where a colicin inhibitory phenotype was observed, but the strains were negative for cib and/or col1. This finding was not overly surprising as there are several different classes of bacteriocin genes that have been identified among enteric bacteria (Cascales et al., 2007) and will warrant further study to elucidate a comprehensive understanding of colicin production and their mechanisms of action. Conversely, some strains examined in the current studies possessed the cib and/or col1 but failed to show inhibition. Spriewald et al. (2015) reported that iron limitation could influence cib gene expression and our studies were not conducted under iron-limited conditions, which could have negatively impacted the toxin production.
The ability of IncI1 plasmids to encode factors that contribute to antimicrobial resistance and virulence is important to public health. Monitoring these plasmids could serve as guide for detecting and potentially limiting spread of resistant and virulent strains and allow for targeted interventions, such as effective antimicrobial therapy, to effectively limit loss arising from outbreaks. Approaches should be undertaken along the farm-to-fork continuum to prevent cross contamination, reduce specific risks, and minimize the spread of clinically important Salmonella and lead to improved public health.
Footnotes
Acknowledgments
The authors would like to thank Drs. Carl Cerniglia, Ashraf Khan, and Huizhong Chen for their insightful review and critique of the article and acknowledge Drs. Alessandra Carattoli and George Jacoby for strains for replicon typing and conjugation studies. Pravin Kaldhone's graduate assistantship was provided by the Center for Advanced Surface Engineering, under the National Science Foundation Grant No. OIA-1457888 and the Arkansas EPSCoR Program, ASSET III.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.