
Abstract
Endophytes are a potential source of novel bioactive compounds with medicinal properties. In this study, 41 endophytic bacteria (EB) were isolated from tissues of a medicinally important plant Taxus brevifolia (Pacific yew). The objective was to screen all the EB isolates for their antibacterial effects against five foodborne pathogenic bacteria: Bacillus cereus ATCC10876, Staphylococcus aureus ATCC12600, Listeria monocytogenes ATCC19115, Escherichia coli ATCC43890, and Salmonella Typhimurium ATCC19585. Among the EB isolates, T. brevifolia seed (TbS)-8, T. brevifolia fleshy part of fruit (TbFl)-10, T. brevifolia leaf (TbL)-22, TbS-29, and TbL-34 exerted significant antibacterial activity against the tested foodborne pathogens. Especially TbFl-10 showed the highest antibacterial activity against all the tested bacteria and was identified as Paenibacillus kribbensis (Pk). Furthermore, an ethyl acetate extract of Pk-TbFl-10 possessed antibacterial activities against the tested five foodborne pathogenic bacteria, with zones of inhibition from 15.71 ± 2.85 to 13.01 ± 2.12 mm. Scanning electron microscopy analysis revealed ruptured, lysed, shrunk, and swollen cells of all the tested foodborne pathogens treated with the ethyl acetate extract of Pk-TbFl-10, suggesting that a metabolite(s) of Pk-TbFl-10 penetrates the cell membrane and causes cell lysis leading to cell death. Our results indicate that Pk-TbFl-10 isolated from T. brevifolia can serve as a novel source of natural antibacterial agents against foodborne pathogenic bacteria, with potential applications in the pharmaceutical industry.
Introduction
E
Foodborne pathogens are a public health problem worldwide, and most of the resultant diseases are caused by bacteria and viruses (The Centers for Disease Control and Prevention [CDC], USA, 2010). In the United States, recently, CDC estimated that 9.4 million cases of illnesses were caused by 31 known foodborne pathogens, with 90% of all illness cases caused by Salmonella, Norovirus, Campylobacter, Toxoplasma, Escherichia coli O157:H7, Listeria monocytogenes, Staphylococcus aureus, and Clostridium perfringens (Addis and Sisay, 2015). These foodborne pathogens are tremendous threats to humans and therefore are diligently controlled especially during manufacture in the food industry (Mith et al., 2014; Afshordi et al., 2016; Khaled et al., 2016). The human population is becoming more concerned about the synthetic antibiotics and expressing increasing demand for the replacement from natural sources day by day. Due to the resistance of pathogenic microbes against current antibiotics, natural components isolated from microbes have been receiving attention as a source of antimicrobial food additives for control of pathogens (Strobel, 2003; Wiyakrutta et al., 2004).
Taxus brevifolia, commonly known as Pacific yew, is a member of Taxaceae and can be used to isolate paclitaxel (or Taxol®) from the bark as a chemotherapeutic drug for treatment of breast and lung cancer (Rao, 1993). Many endophytic fungi isolated from Taxus spp. have been reported to produce important bioactive metabolites (Wang et al., 2007). For example, several endophytic fungi, including Taxomyces andreanae isolated from T. brevifolia, produce paclitaxel and other biocontrol agents (Stierle et al., 1993; Pereira et al., 1999; Strobel et al., 2004).
Nonetheless, there have been no reports of the organ- or tissue-specific EB isolated from T. brevifolia. Therefore, this study was undertaken to isolate EB from different organs of T. brevifolia and to evaluate their antibacterial capacity in vitro against foodborne pathogenic bacteria, including three Gram-positive bacterial spp., Bacillus cereus, L. monocytogenes, and S. aureus and two Gram-negative bacterial spp., E. coli and Salmonella Typhimurium. To our knowledge, this is the first report of isolation of EB from T. brevifolia with a practical application.
Materials and Methods
Isolation and identification of EB
Healthy twigs of T. brevifolia were collected from the plant garden of Yeungnam University, Republic of Korea, in October 2014. The twigs collected in plastic bags were brought to the laboratory immediately, leaves and fruits separated and washed thoroughly under running tap water. The isolation of EB was carried out as per the standard procedure (Costa et al., 2012), with a slight modification. In brief, the washed samples were properly dried, weighed on a digital balance, and disinfected by 70% alcohol for 60 s, followed by 2% sodium hypochlorite for 180 s, and 100% ethanol for 30 s. Then, surface-disinfected tissues were rinsed with sterile distilled water at least three times and dried properly with autoclaved filter paper. The disinfected plant parts were ground with a sterilized aqueous NaCl solution (0.9%) at 3 mL/g, using a sterile mortar and pestle. The tissue extracts were incubated at 28°C for 3 h to enable a complete release of EB from the host tissues.
For the isolation of EB, the ground tissue extracts were diluted 10−1 and 10−2 with a sterilized aqueous NaCl solution (0.9%), and spread onto different types of 25% media plates, including yeast extract-nutrient agar (YNA), nutrient agar (NA), de Man, Rogosa, Sharpe (MRS), and Luria–Bertani (LB; Becton, Dickinson and Company, Franklin Lakes, NJ) to obtain single bacterial colonies. The plates were incubated for up to 15 days at 28°C. All the colonies were counted and expressed in colony-forming units (CFU) per gram of fresh tissue. Single-colony isolation was repeated at least three times for purification of each of the EB isolates. Finally, the selected EB isolates were cultured in different media and were stored in YNA media as a 50% glycerol stock at −80°C until use.
The EB isolates were identified based on16S rRNA gene sequencing. The sequencing data, alignment, and the phylogenetic tree were studied by means of the Molecular Evolutionary Genetics Analysis (MEGA) software (version 6.0). The phylogenetic tree analysis was accompanied by the neighbor-joining method (Tamura et al., 2013; Susilowati et al., 2015).
Antibacterial assay of EB against foodborne pathogenic bacteria
Total of five foodborne pathogenic bacteria, including three Gram-positive, B. cereus ATCC10876, L. monocytogenes ATCC19115, and S. aureus ATCC12600, and two Gram-negative E. coli ATCC43890 and Salmonella Typhimurium ATCC19585, were used in the present study to determine the antibacterial activities of the isolated EB. These foodborne pathogenic bacteria were obtained from the American Type Culture Collection (ATCC, Manassas, VA).
The EB isolates were screened for their antibacterial activity following the standard procedure (Roh et al., 2009), with small modifications. Screening was performed by culturing total isolated EB in 96-well plates with 10 μL of an individual bacterial strain grown in yeast extract-nutrient broth (YNB) and incubated for 36 h at 28°C. Next, the cultured bacteria were replicated and grown on 25% YNA (1.5% agar) medium for 36 h at 28°C. After that, completely grown EB were killed by addition of 1 mL of chloroform to the inversional addition on the lids of Petri plates, and incubation for 15 min. The lids of the plates were opened and allowed to stand for 20 min to remove traces of chloroform, and then, the plates were treated with UV light for 15 min to kill the bacteria completely. Simultaneously, 30 μL cultures of the five pathogenic bacteria grown overnight in the YNB at 37°C were resuspended in 10 mL of the fresh YNB at 55°C, mixed gently, and then poured slowly and uniformly on the patch plate without disturbing the plate. After solidification, all the Petri plates were sealed and incubated at 37°C for 24 h, and the diameter of inhibition zones was measured using an electronic digital caliper with precision up to 1/100 mm. Kanamycin and streptomycin that served as reference standard antibiotics were applied at 10 μg/disc.
Antibacterial activity of the solvent-extracted EB against foodborne pathogenic bacteria
The selected EB were subcultured in YNA plates for 24 h; next, a single colony was picked and inoculated into 200 mL of YNB in 1 L Erlenmeyer flasks followed by incubation with continuous shaking at 28°C for 36 h at 180 rpm. The culture broth was centrifuged at 3500 × g for 15 min at 4°C, and the supernatant was collected. The supernatant was mixed with an equal volume of ethyl acetate and then incubated overnight. Next day, the ethyl acetate layer was collected by means of a separating funnel and dried using a rotary evaporator (A-1000S; EYELA, Japan) at 50°C; we collected the dried extracts from the evaporated flask, redissolved them in 2 mL of methanol, and air-dried them in tubular glass vials. The ethyl acetate extract amount was measured, and the extract was dissolved in 5% dimethyl sulfoxide (DMSO) in methanol to 0.1 g/mL. At this concentration, 50 μL of the ethyl acetate extract per well (5 mg/well) was used to evaluate the antagonistic activity against the bacterial pathogens by an agar well diffusion assay, with measurement of the diameter of the inhibition zone in mm by means of an electronic digital caliper.
Determination of minimum inhibitory concentration and minimum bactericidal concentration
The minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of the ethyl acetate extract of TbFl-10 were determined against the foodborne pathogenic bacteria by the twofold dilution method (Patra et al., 2015). In brief, 5% DMSO was used as a control, and serial dilutions were prepared by mixing 5% DMSO with the ethyl acetate extract for the determination of MIC and MBC. Different dilutions (5, 2.5, 1.25, 0.625, 0.3125 mg/mL) of the TbFl-10 ethyl acetate extract in the YNB medium were prepared, and the lowest concentrations of TbFl-10 that did not allow any visual growth of the tested foodborne pathogenic bacteria were defined as the MIC, expressed in mg/mL. The MIC and the next higher concentration of the sample were selected, spread on the YNA in Petri plates, and incubated at 37°C for 24 h; the lowest concentration of TbFl-10 that did not allow for any growth of the pathogenic bacteria in the 25% YNA plates was defined as the MBC, expressed in mg/mL.
Scanning electron microscopy analysis
The effects of TbFl-10 ethyl acetate extract on the surface morphology of the tested five foodborne pathogenic bacteria were determined by scanning electron microscopy (SEM) analysis. Both the control treated with 5% DMSO and five foodborne bacteria treated with TbFl-10 ethyl acetate extract at MIC were analyzed by SEM, using a standard protocol of Bajpai et al. (2009).
Statistical analysis
All the experimental data are expressed as mean ± standard deviation from three independent replicates in each experiment. Statistical analysis of the results was conducted by one-way analysis of variance followed by Duncan's multiple-range test at p < 0.05, using the Statistical Analysis Software (SAS, version: SAS 9.4; SAS Institute, Inc., Cary, NC).
Results
Isolation of EB
A total of 41 EB isolates were isolated from healthy leaves (23 isolates), seeds, (8 isolates) and the flesh in fruits (10 isolates) of T. brevifolia. (Supplementary Table S1; Supplementary Data are available online at
Antibacterial activity of EB against foodborne pathogens
All the EB isolates were tested for their antibacterial activity against five foodborne pathogenic bacteria, and among them 12 EB harbored at least one positive antibacterial activity in a preliminary screening procedure (Table 1). Out of the 12 EB strains, 5 EB strains, namely T. brevifolia seed (TbS)-8, T. brevifolia fleshy part of fruit (TbFl)-10, T. brevifolia leaf (TbL)-22, TbS-29, and TbL-34 showed positive antibacterial activity against at least 3 foodborne pathogenic bacteria: therefore, these 5 EB were selected for further study (Table 2). The five selected EB showed varying degrees of antagonistic activity (10.23–23.04 mm) against the tested foodborne pathogens (Table 2). TbFl-10 showed the highest antibacterial activity against all foodborne pathogens tested (Table 2 and Fig. 1). The 16S rRNA gene sequencing analysis identified the selected five EB isolates, TbS-8, TbFl-10, TbL-22, TbS-29, and TbL-34, as Bacillus altitudinis, Paenibacillus kribbensis (Pk), Bacillus thuringiensis, Paracoccus marinus, and B. altitudinis, respectively (Table 1). The phylogenetic tree analysis revealed that isolate TbFl-10 shares 99% identity with the 16S rRNA gene sequences of Pk (accession number NR025169).
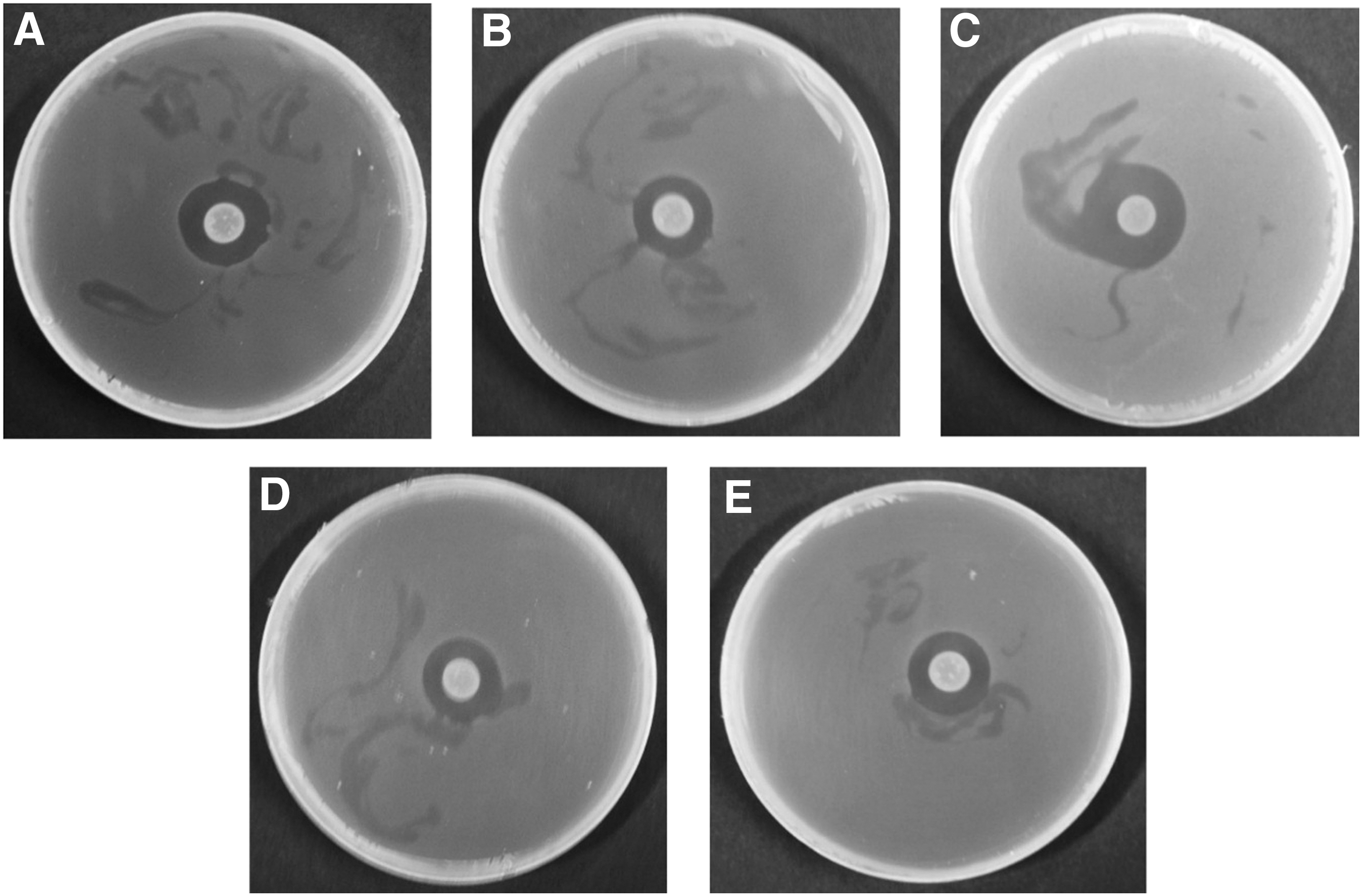
Antibacterial activity of TbFl-10 (Paenibacillus kribbensis) against all the tested foodborne pathogenic bacteria:
B. Cereus, Bacillus cereus ATCC 10876; L. monocytogenes, Listeria monocytogenes ATCC19115; S. aureus, Staphylococcus aureus ATCC 12600; E. coli, Escherichia coli ATCC 43890; S. Typhimurium, Salmonella Typhimurium ATCC 19586. TbS, Taxus brevifolia seed; TbFl, T. brevifolia fleshy part of fruit; TbL, T. brevifolia leaf. +, indicates positive activity, diameter of zones of inhibition less than 10 mm; ++, indicates medium activity, diameter of zones of inhibition between 10 and 13 mm; +++, indicates strong activity, diameter of zones of inhibition greater than 13 mm.
Values with different superscript letters are significantly different, and with the same superscript letters are not significantly different at p < 0.05 according to Duncan's test.
Diameter of the inhibition zone (in mm) of three independent replicates expressed as mean ± standard deviation.
10 μg/disc, reference standard antibiotics.
TbS, Taxus brevifolia seed; TbFl, T. brevifolia fleshy part of fruit; TbL, T. brevifolia leaf; —, no inhibition zone.
The five EB spp. were further tested for their antibacterial activity with the ethyl acetate extracts. In the assay of ethyl acetate extracts of the five selected EB spp., only Pk-TbFl-10 showed antibacterial activity against all the five foodborne pathogenic bacteria (Table 3 and Fig. 2). Especially, the extract of Pk-TbFl-10 exerted the highest antibacterial activity against B. cereus, with the inhibition zone of 15.71 ± 2.85 (Table 3 and Fig. 2).
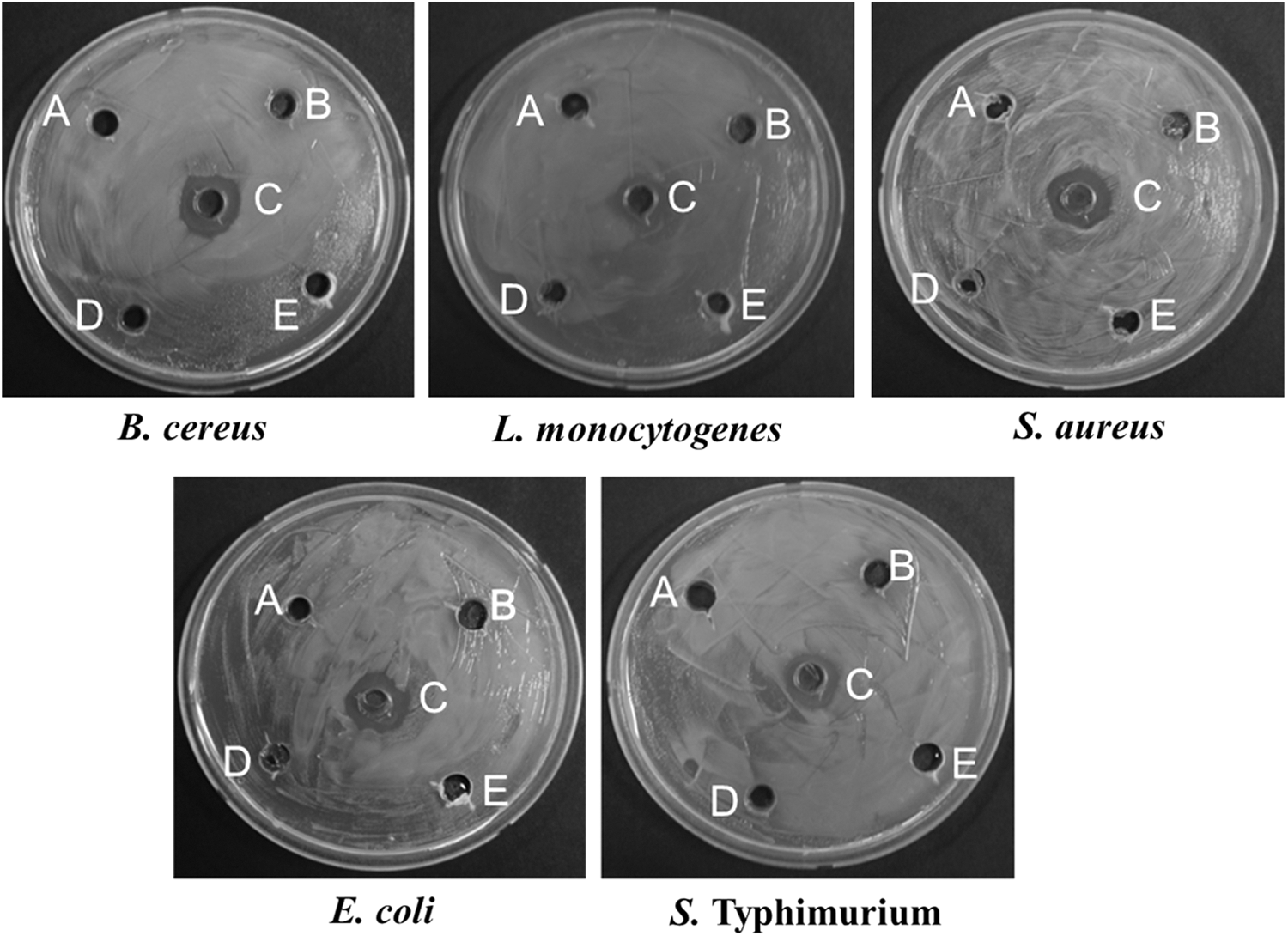
Antibacterial capacity of ethyl acetate extracts of the selected five endophytic bacteria against foodborne pathogenic bacteria. (
MIC and MBC were also determined for the ethyl acetate extract of TbFl-10. Values with the same superscript letters are not significantly different at p < 0.05.
Diameter of the inhibition zone (in mm) of three independent replicates expressed as mean ± standard deviation.
MIC and MBC values are expressed in mg/mL.
MBC, minimum bactericidal concentration; MIC, minimum inhibitory concentration; TbS, Taxus brevifolia seed; TbFl, T. brevifolia fleshy part of fruit; TbL, T. brevifolia leaf; —, no inhibition zone produced.
Determination of MIC and MBC
Among the antagonistic EB spp., only the ethyl acetate extract of Pk-TbFl-10 showed antibacterial activity against all the tested foodborne pathogens; as a result, it was selected for the determination of MIC and MBC. The ethyl acetate extract of Pk-TbFl-10 showed moderate to strong antibacterial effect in the MIC analysis against the tested foodborne pathogenic bacteria. The values of MIC and MBC of the ethyl acetate Pk-TbFl-10 extract ranged from 1.25 to 5.00 mg/mL and from 2.50 to >5.00 mg/mL, respectively (Table 3).
SEM analysis of the solvent extract of TbFl-10
The antibacterial effects of the ethyl acetate extract of Pk-TbFl-10 on the cell morphology were observed against foodborne pathogenic bacteria (Fig. 3). All the tested bacterial spp. were treated with the ethyl acetate extract of Pk-TbFl-10, and the cell surface was found to be ruptured (rough surface), with elongated, shrunk, lysed, and swollen cells compared with the untreated five foodborne pathogenic bacteria showing a regular, uniform, and smooth surface (Fig. 3).

Photomicrographs (scanning electron microscopy images) of foodborne pathogenic bacteria treated with 5% DMSO as a control
Discussion
Food serves as an effective culture and maintenance medium for various microbes that cause foodborne diseases, and it can also allow various microbes to grow and produce several toxins (Ganguly et al., 2012). Among the pathogenic microbes, B. cereus, L. monocytogenes, S. aureus, E. coli, and Salmonella Typhimurium are the major foodborne pathogenic bacteria and may cause life-threatening diseases and symptoms, including pneumonia, meningitis, osteomyelitis, endocarditis, sepsis, bloody diarrhea, and food poisoning (McClements and Decker, 2000; Abuladze et al., 2008; Lim et al., 2010; Afshordi et al., 2016; Seo et al., 2016). Thus, the aim of our study was to find reliable EB strains to develop a new formulation (for an economical and sustainable strategy for control of foodborne pathogens) from EBs effective bioactive natural compounds.
A total of 41 EB isolates were isolated from different tissues of T. brevifolia, including leaves, seeds, and the fleshy part of fruits (Supplementary Table S1). The bacterial population densities shown as CFU per gram of fresh weight varied depending on the tissues (Supplementary Table S1). Similar reports of variation in densities of EB depending on plant tissues, collection locations, and environmental conditions have also been published (Quadt-Hallman et al., 1997; Hallmann et al., 2002; Sturz and Nowak, 2002). In T. brevifolia, the highest EB density was observed in the leaves (Supplementary Table S1).
When all the EB isolates were tested for their antibacterial activity against five foodborne bacterial pathogens, 12 isolates were selected, based on the antibacterial activity (Table 1). In agreement with our results, EB isolated from a medicinal fern Equisetum arvense L. also exerts antibacterial activity against foodborne pathogenic bacteria (Das et al., 2016). According to the second screening data, the five EB isolates exerted antibacterial activity against at least three tested pathogenic bacteria (Table 2). Among the antagonist EB, Bacillus is a well-known and important bacterial heterogeneous taxon, ubiquitous in nature (Alina et al., 2015). Various spp in the Bacillus genus isolated from different crops are economically important because of production of several metabolites and other biotechnological approaches for different purposes, including food, pharmaceutical, environmental, and industrial uses (Tayung et al., 2007, 2011; Melnick et al., 2008; Kumar et al., 2014). Paracoccus sp. can produce important secondary metabolites; P. marinus sp. nov. isolated from coastal seawater in Tokyo Bay was a producer of adonixanthin diglucoside (Khan et al., 2008). Paenibacillus spp. are also important owing to their production of various extracellular enzymes, polysaccharides, amino acids, and secondary metabolites (Seo et al., 1999; Chung et al., 2000). A number of secondary metabolites produced by Paenibacillus sp. act as antibiotic agents (Lorentz et al., 2006). The antagonistic activity has been confirmed against a broad spectrum of microbes (Beatty and Jensen, 2002; Berge et al., 2002; Weid et al., 2003; Ivy et al., 2012).
The ethyl acetate extract of Pk-TbFl-10 exerted robust antibacterial activity against all the tested foodborne pathogenic bacteria among all the isolated EB (Table 3 and Fig. 2). The ethyl acetate extract of Pk-TbFl-10 also showed moderate to strong ranges of values of MIC and MBC against the foodborne bacteria (Table 3). High inhibition zones and the low MIC values have also been reported for Paenibacillus sp. against foodborne pathogenic bacteria, including Bacillus subtilis (NCIM2063), S. aureus (MTCC 737), E. coli (MTCC739), and Pseudomonas aeruginosa (MTCC741) (Lu et al., 2000; Tayung et al., 2007, 2011; Gogoi et al., 2008). In addition, robust antimicrobial activity of Paenibacillus sp. against E. coli and Candia albicans has been reported (Aw et al., 2016).
Metabolites of EB may diffuse in the culture medium and suppress the growth of foodborne pathogenic bacteria. The ethyl acetate extract of Pk-TbFl-10 showed a strong antibacterial activity against all the tested bacteria; the probable explanation may be the bioactive compound size: smaller than that in other antagonist EB spp. The bioactive compound could easily move into the bacterial cell membrane via the general bacterial porins, which might be responsible for several metabolic functions of the cell, or they may form various pores in the outer cell membrane of bacteria, thus resulting in the leakage of internal substances to the outside, causing cell lysis and death (Kotzekidou et al., 2008). This mechanism was verified by SEM analysis, which indicated that Pk-TbFl-10 might contain promising bioactive compounds for deteriorating the cellular membranes of the tested bacteria (Fig. 3). The cell morphology of the foodborne pathogenic bacteria treated with the ethyl acetate extract of Pk-TbFl-10 was found to be irregular, collapsed, and with a rough surface and deformities (Fig. 3). Similar morphological deviations have been observed in Xanthomonas campestris pv. glycine and Pythium myriotylum treated with the metabolites of endophytic Actinomycetes spp. and Fusarium spp. (Mingma et al., 2014; Keerthi et al., 2016).
Conclusions
On the basis of the data and aforementioned explanations, it can be concluded that the identified Pk-TbFl-10 isolated from T. brevifolia has a strong antibacterial activity against foodborne pathogenic bacteria; this effect may be due to bioactive compounds produced by Pk-TbFl-10. Intact EB or the extract containing pure compounds may be useful as natural antimicrobials for food preservation or protection. Further research is needed to find the pure bioactive metabolite(s) amenable to large-scale production. They can be used for pharmaceutical as well as biotechnological applications.
Footnotes
Acknowledgments
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through Agri-Bio industry Technology Development Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA) (117044-3).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.