
Abstract
Nontyphoidal Salmonella enterica is a common cause of illness in humans ranging from gastroenteritis to invasive disease. National surveillance programs continually monitor trends in antimicrobial resistance patterns and mechanisms of resistance to identify emerging public health threats. Our study shows the emergence of nonsusceptibility to both levofloxacin and ceftriaxone, a concerning phenotype that threatens first-line antibiotic therapy, in Salmonella isolates recovered between 2014 and 2015. From 2010 to 2013 the rate of resistance increased from 0.0% (0/1181) to 1.5% (9/593) in 2014 and 2015. The isolates with this phenotype were found to be from multiple serotypes, including Typhimurium, Newport, and Enteritidis. Resistance to ceftriaxone was attributed to the presence of either an AmpC or extended-spectrum β-lactamase, and resistance to fluoroquinolones was attributable to the presence of mutations in the quinolone resistance-determining region or the presence of plasmid–mediated quinolone resistance genes. As this resistance pattern was seen in a variety of Salmonella serotypes harboring varied resistance mechanisms, it indicates a worrying trend in the spread of isolates resistant to both first-line treatment options.
Introduction
N
To monitor antibiotic resistance of Salmonella isolates, all isolates recovered from human specimens in Nebraska are submitted to the Nebraska Public Health Laboratory (NPHL) for antimicrobial susceptibility testing, serotyping, and pulsed-field gel electrophoresis (PFGE) as part of the national PulseNet program (Gerner-Smidt et al., 2006). The performance of antimicrobial susceptibility testing on isolates in Nebraska and throughout the country by the Centers for Disease Control and Prevention (CDC's) National Antimicrobial Resistance Monitoring System (NARMS) provides a mechanism to monitor and combat potential emerging resistance problems in the United States. Beginning in 2014, we observed an increase in the number of isolates that showed reduced susceptibility (disk diffusion ranges interpreted as intermediate or resistant) to ceftriaxone and nalidixic acid, which has conventionally been used as a screen for fluoroquinolone susceptibility. Investigation into the emergence of this phenotype revealed Salmonella isolates of various serotypes that harbored a variety of resistance mechanisms. Previous work from NARMS indicates that from 2004 to 2012 only 38 of the 19,410 Salmonella isolates tested were nonsusceptible to both ceftriaxone and fluoroquinolones (Medalla et al., 2016). This report describes the investigation of the concerning emergence of ceftriaxone and levofloxacin nonsusceptibility in nontyphoidal Salmonella isolates recovered in Nebraska between 2014 and 2015.
Materials and Methods
Isolates
All isolates of Salmonella recovered from human specimens in Nebraska from 2010 to 2015 were reviewed for inclusion in this study. In total, 1783 nontyphoidal Salmonella isolates were submitted to the NPHL for antimicrobial susceptibility testing and strain typing.
Antimicrobial susceptibility testing
Disk diffusion was used to screen for antimicrobial susceptibility to ceftriaxone and nalidixic acid. Testing was performed according to Clinical and Laboratory Standards Institute (CLSI) standards for the appropriate year of testing (CLSI M100 S20, S21, S22, S23, S24, S25). Isolates with reduced susceptibility to ceftriaxone and nalidixic acid had minimum inhibitory concentrations (MICs) for levofloxacin, ceftriaxone, and cefoxitin determined by Etest® (bioMerieux, Durham, NC). Zone sizes and MICs were interpreted according to the current CLSI standards.
Pulsed-field gel electrophoresis
All isolates were tested by PFGE using the protocol established by PulseNet (Gerner-Smidt et al., 2006). PFGE analysis of XbaI chromosomal digests was performed using BioNumerics software to determine strain relatedness (Applied Maths, Sint-Martens-Latem, Belgium).
Serotyping
Serotyping was performed using the xMAP Salmonella Serotyping Assay (Luminex Corporation, Austin, TX) according to manufacturer's instructions. Testing was supplemented with O and H antigen agglutination testing when needed with reagents and protocol supplied by the CDC.
Polymerase chain reaction amplification and DNA sequencing
Genomic DNA was extracted from overnight cultures of Salmonella using the DNeasy Blood and Tissue Kit (Qiagen, Germantown, MD). Polymerase chain reaction (PCR) was used to amplify the quinolone resistance determining regions (QRDRs), including the gyrA, gyrB, parE, and parC genes, followed by sequencing to identify mutations associated with fluoroquinolone resistance. Isolates were also screened by PCR for the presence of plasmid–mediated quinolone resistance (PMQR) genes qnrS, oqxAB, qnrD, and aac(6’)-lb-cr as previously described (Kao et al., 2017). β-lactamase–mediated resistance was evaluated in ceftriaxone–resistant Salmonella isolates using the ARM-D™ AmpC Detection Kit (Streck, Inc., Omaha, NE) to detect plasmid–encoded AmpC β-lactamases and the ARM-D for β-lactamase ID (Streck, Inc.) to detect common extended-spectrum β-lactamase (ESBL) (CTX-M-14 and CTX-M-15), AmpC (DHA and CMY), and carbapenemases (KPC, NDM, IMP, VIM, and OXA-48).
Results
Resistance patterns
A total of 1783 nontyphoidal S. enterica isolates underwent antimicrobial susceptibility testing from 2010 to 2015 (Table 1). Before 2014, no isolates demonstrated reduced susceptibility to both nalidixic acid and ceftriaxone. However, one isolate with this resistance phenotype was identified in 2014, isolate 4360. By 2015, eight additional Salmonella isolates were identified as nonsusceptible to both nalidixic acid and ceftriaxone by the disk diffusion method. These nine isolates had ceftriaxone MICs ranging from 8 to >256 μg/mL when examined by Etest (Table 2). These same isolates had levofloxacin MICs ranging from 1 to >32 μg/mL (Table 2).
CRO, ceftriaxone; NA, nalidixic acid; CIP, ciprofloxacin.
gyrA, gyrB, parC, and parE genes were sequenced for mutations. The presence of qnrS, qnrD, oqxAB, and aac(6’)-lb-cr genes was evaluated by PCR.
LEV, levofloxacin; FOX, cefoxitin; PCR, polymerase chain reaction.
Identification of resistant isolates
To determine if the increase in recovery of the resistant isolates was due to clonal expansion, the serotypes and PFGE patterns of these isolates were examined. Isolate 4360, identified in 2014, was identified as Salmonella Enteritidis, whereas the Salmonella isolates that emerged in 2015 were identified as serotypes Typhimurium (3 isolates), 4,[5],12:i:- (4 isolates), and Newport (1 isolate). PFGE analysis showed five unique DNA restriction patterns based on the presence of at least a three-band difference among the Salmonella isolates (Fig. 1). The three isolates of Salmonella Typhimurium had indistinguishable PFGE patterns as did the three isolates of Salmonella 4,[5],12:i:-, suggesting that they were clonal.
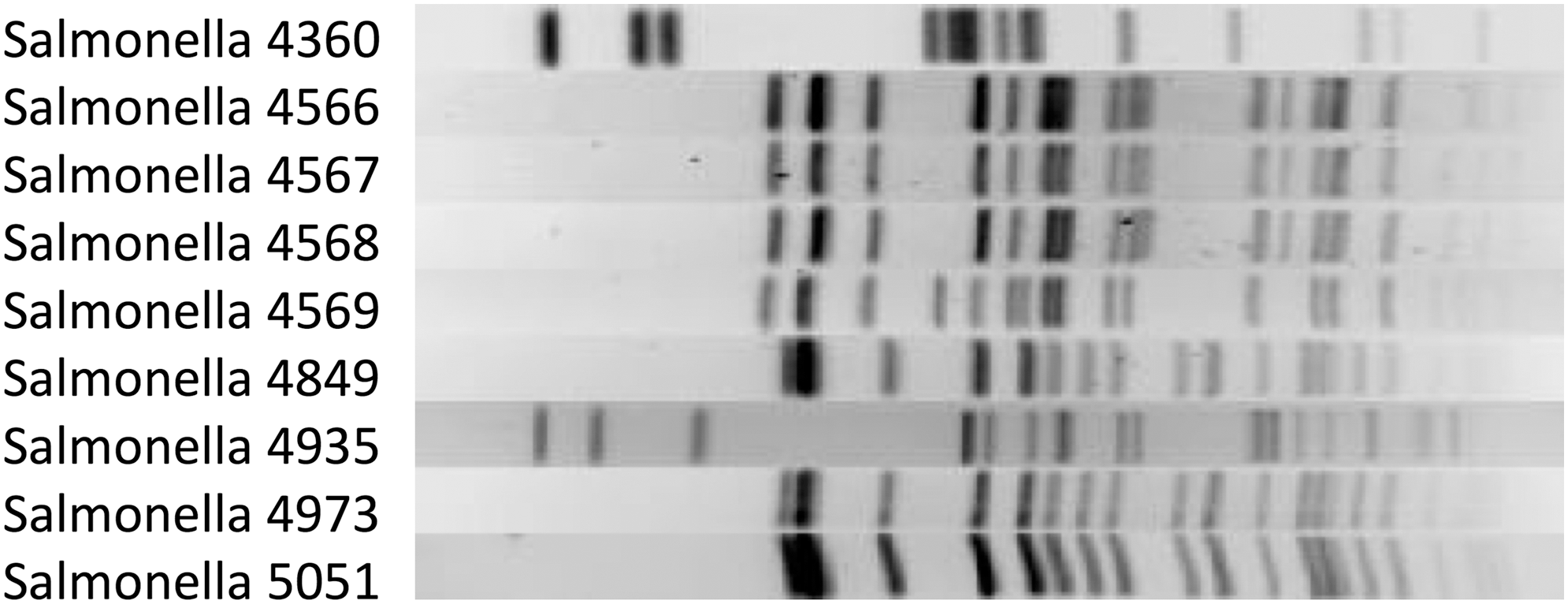
PFGE analysis of XbaI chromosomal digests of nine isolates of Salmonella enterica with nonsusceptibility to ceftriaxone and levofloxacin. PFGE illustrates that 4566, 4567, and 4568 are clonally related (all serotype 4,[5],12:i:-), as are 4849, 4973, and 5051 (all serotype Typhimurium). PFGE, pulsed-field gel electrophoresis.
Resistance mechanisms
Resistance to β-lactams was evaluated using multiplex PCR kits. Salmonella Enteritidis and 4,[5],12:i:- isolates harbored bla CTX-M-15, while the Salmonella Newport and Typhimurium isolates were positive for bla CMY-2 (Table 2). The mechanisms of resistance to the quinolones were determined by sequencing the QRDR in all nine isolates. No mutations were identified within the QRDR of the four Salmonella 4,[5],12:i:- isolates. In the three isolates of Salmonella Typhimurium, one mutation identified in GyrA was an amino acid change Y83S. However, the isolate from 2014, Salmonella Enteritidis, had the only significant changes in the QRDR with a change in parC (S80I) and two mutations in parE (R92K and N100H), which may contribute to the high levofloxacin MIC of this isolate (Table 2). Further analysis of the PMQR in nonsusceptible isolates showed that qnrS, encoding a topoisomerase protection gene, was present in the isolates of Salmonella 4,[5],12:i:- (Table 2). The qnrS gene was not detected in isolates of Salmonella Typhimurium, but they were positive for the presence of the quinolone efflux pump, oqxAB genes. All isolates were also assayed and negative for the detection of qnrD and aac(6’)-lb-cr. Interestingly, no mutations or known fluoroquinolone resistance genes were identified in the Salmonella Newport isolate.
Discussion
In this study, we highlight the concerning emergence of nontyphoidal Salmonella isolates that are nonsusceptible to both ceftriaxone and the fluoroquinolones in Nebraska. A previous report from NARMS indicated that over a 9-year period, only 38 isolates with this pattern were seen nationwide (Medalla et al., 2016), indicating that the nine isolates found in a 2-year period in one state alone may suggest a growing public health threat. To further characterize this trend, we examined the phenotypic properties and antibiotic resistance mechanisms associated with these isolates.
Examining the resistance patterns of nontyphoidal Salmonella isolates collected in Nebraska from 2010 to 2015 showed that the prevalence of ceftriaxone and fluoroquinolone nonsusceptible isolates progressively increased from 0.0% during 2010–2013 to 0.3% in 2014 and 2.8% in 2015. The nontyphoidal Salmonella isolates in which this pattern was observed were from serotypes Enteritidis, Typhimurium, Newport, and 4,[5],12:i:-. In addition to serotype differences, these isolates also had five distinct PFGE patterns. Three isolates of Salmonella 4,[5],12,i:-. had indistinguishable XbaI restriction patterns. Epidemiological investigations suggested that these isolates were represented in an associated outbreak. Unfortunately, these serotypes make up four of the five most common serotypes seen in human infections in the United States (CDC, 2016). Previous reports of multidrug-resistant Salmonella that showed resistance to ceftriaxone and decreased susceptibility to the fluoroquinolones have been shown in Salmonella Albert recovered from turkeys, retail ground turkey, and human infections from individuals with poultry exposure (Folster et al., 2015). The serotypes in this study are not geographically restricted to the midwestern United States like Salmonella Albert. Given the various serotypes and PFGE patterns seen suggest that these isolates were potentially acquired from a diversity of sources making the clonal expansion of isolates with this resistance pattern difficult to control.
Cephalosporin resistance has been well documented in Nebraska. One of the first cases of domestically acquired infection with ceftriaxone-resistant Salmonella in the United States was recorded in Nebraska in 2000, presumably through infection from cattle (Fey et al., 2000). This first isolate identified in Nebraska had developed ceftriaxone resistance through the acquisition of CMY-2 AmpC β-lactamase. Multiple studies have since shown that CMY-2 is the most common β-lactamase in Salmonella infections from human and animal sources in the United States (Dunne et al., 2000; Sjölund-Karlsson et al., 2013; CDC, 2016). The CMY AmpC β-lactamase was detected in four isolates of interest in this study. In the five other isolates, resistance to ceftriaxone was found to be attributable to an ESBL, CTX-M-15 (Table 2). Historically this β-lactamase has not been seen as frequently in Salmonella as a cause of cephalosporin resistance, but recently its incidence has been increasing (Sjolund-Karlsson et al., 2011).
Fluoroquinolone resistance in S. enterica has traditionally been associated with mutations in the QRDR. Since the late 1990s additional mechanisms of resistance to quinolones have expanded to include the PMQR genes such as qnrs which encode proteins that protect DNA gyrase from the quinolones, AAC(6’)-Ib-cr, which is capable of reducing ciprofloxacin activity and quinolone efflux pumps (Sjolund-Karlsson et al., 2009). The most recent report from NARMS showed that isolates with decreased susceptibility to ciprofloxacin frequently had mutations in gyrA (48/77 isolates), in addition to isolates containing PMQR genes qnrB (13/77), qnrS (5/77), oqxAB (4/77), or aac(6’)lb-cr (1/77) (CDC, 2016). Interestingly, whole genome sequencing (WGS) showed that 15.9% (12/77) of the isolates with reduced susceptibility to ciprofloxacin had no known genes or mutations associated with ciprofloxacin resistance. The inability to identify resistance mechanisms by WGS highlights a limitation of this method to correlate phenotypic results with genotypic information. In our study 11.1% (1/9) of the isolates we examined had no attributable mutations or genes associated with reduced susceptibility to the fluoroquinolones (Table 2). The most commonly detected mechanism was the topoisomerase protection gene, qnrS, followed by a quinolone efflux pump encoded by the gene oqxAB.
Conclusions
The findings from this study highlight the emergence of a concerning resistance pattern in Salmonella isolates recovered from human specimens in Nebraska in 2014 and 2015. In addition, these data add to a growing body of evidence that resistance to fluoroquinolones in the United States and worldwide is increasingly driven by the presence of PMQR genes and not solely by mutations in the QRDR. This report demonstrates that multiple mechanisms of resistance to ceftriaxone and the fluoroquinolones can emerge in different Salmonella serotypes associated with human infections. The emergence and continued spread of isolates with nonsusceptibility to first-line antibiotics such as ceftriaxone and the fluoroquinolones is a public health concern. Continued nationwide surveillance programs using both phenotypic and genotypic methods are needed to monitor resistance patterns and mechanisms of resistance in nontyphoidal Salmonella.
Footnotes
Acknowledgments
None of this work could have been done without the Clinical Microbiology Laboratory staff at Nebraska Medicine for performing antimicrobial susceptibility testing and staff at the Nebraska Public Health Laboratory for performing serotyping and pulsed-field gel electrophoresis. Special thanks to Dianna Bannister of Nebraska Medicine for recognizing this initial resistance pattern that prompted this investigation and to Amanda Heeg of the Nebraska Public Health Laboratory for compiling yearly susceptibility data.
Disclosure Statement
No competing financial interests exist.