
Abstract
Staphylococcus aureus is a frequent and major contagious mastitis bacterial pathogen. The antibiotic treatment cure rates vary considerably from 4% to 92%. Staphylococcus aureus readily becomes resistant to antibiotics, resulting in persistent noncurable intramammary infection that usually results in culling of infected animals. Because of its notorious ability to acquire resistance to the commonly used as well as last resort antimicrobials such as methicillin and vancomycin and the development of multidrug-resistant strains, antimicrobial resistance (AMR) in S. aureus is of paramount importance in human medicine. The objective of this study was to evaluate the prevalence of AMR and genetic diversity of S. aureus isolates from milk of dairy cattle. Staphylococcus aureus isolates (n = 239) from 33 dairy farms in Tennessee were tested against 10 antimicrobials by broth microdilution method using the Sensititer system. Genetic diversity of resistant isolates was evaluated by pulsed-field gel electrophoresis (PFGE). Overall, AMR of the S. aureus isolates varied from as low as 1.3% for ceftiofur to as high as 25% for sulfadimethoxine. Out of 239 S. aureus isolates, 82 (34.3%) of them were resistant to at least 1 of the 10 antimicrobials. The AMR isolates belonged to two major PFGE types, indicating the presence of dominant clonal patterns among the resistant isolates. In general, there was a variation of prevalence of AMR within and among farms over time, with an increasing trend in tetracycline resistance. Judicious use of antimicrobials in dairy cattle farms can reduce the development of antimicrobial-resistant S. aureus.
Introduction
I
Data from the 2007 USDA National Animal Health Monitoring System (USDA APHIS, 2008b) indicate that the percentages of farms that treated cows with any antibiotic were 85.4% for mastitis, 58.6% for lameness, 55.8% for respiratory, 52.9% for reproductive, 25.0% for diarrhea or other digestive problems, and 6.9% for all other conditions (Seegers et al., 2003; Petrovski et al., 2006; USDA APHIS, 2008a; Hogeveen et al., 2011; Oliver et al., 2011; Rollin et al., 2015). Therefore, mastitis is the most common disease of dairy cows treated with antimicrobials (Oliver et al., 2011).
Staphylococcus aureus mastitis is the most frequently encountered disease of dairy cattle throughout the world. Prevalence of bovine S. aureus mastitis ranges from 40% to 50% in lactating cow herds, in which two or more udder quarters are infected (Mellenberger and Keirk, 2001). Dairy farmers rely on antimicrobials for the treatment and prevention of mastitis (Saini et al., 2012a, 2012b).
The prophylactic use of antibiotics as a blanket dry cow therapy exposes large number of healthy animals to antibiotics, thus increasing selective pressure on commensal bacteria. More than 90% of U.S. dairy farms use intramammary antibiotics at dry off (USDA APHIS, 2008a) and 80% of them treat all cows on the farm (Wagner and Erskine, 2013). Antibiotics are administered to dairy cows through two primary routes: intramuscular and intramammary (USDA APHIS, 2009), which can exert selective pressure both to the mammary pathogenic bacteria and nonmammary commensal bacteria in the gastrointestinal tract. Nonprudent use of antimicrobials in dairy farms increases selection pressure on mastitis-causing and commensal bacteria (Barbosa and Levy, 2000; Barber et al., 2003). Once resistant bacteria emerge on the farm, the resistant strains persistently circulate on the farm and farm environment (Normanno et al., 2007), building up reservoirs of antimicrobial resistance (AMR), which may result in distribution of resistance genes in human pathogens (Wichmann et al., 2014). Therefore, regular monitoring of AMR is important not only to evaluate the development of AMR bacteria but also for the selection of effective treatment and assessment of prognosis (van Duijkeren et al., 2003; CLSI, 2015).
Resistance of S. aureus to antimicrobials is mainly because of its inherent ability to form biofilms (Melchior et al., 2006), encapsulated lesions, and acquisition of AMR genes through horizontal transfer of mobile genetic elements, such as plasmids, bacteriophages, and genomic islands (Brussow et al., 2004; Owens et al., 1988, 1997). The emergence of aggressive antibiotic-resistant S. aureus strains (such as methicillin-resistant S. aureus [MRSA]) has become a significant problem, as these strains have spread beyond their typical hospital milieu to the environment and are commonly isolated from cattle (Holmes and Zadoks, 2011; Fitzgerald, 2012a, 2012b).
Some reports indicate that only a few strains are largely responsible for mastitis (Fitzgerald et al., 1997; Zadoks et al., 2002; Anderson et al., 2006; Chambers and Deleo, 2009; Sakwinska et al., 2011). The genetic diversity of S. aureus isolates can be evaluated by phage typing, multilocus metabolic-enzyme typing, multilocus sequence typing, and pulsed-field gel electrophoresis (PFGE) methods (Peacock et al., 2002; Zadoks et al., 2002; van Leeuwen et al., 2003; O'Brien et al., 2004; Robinson and Enright, 2004; Sabour et al., 2004; Anderson et al., 2006; Cookson et al., 2007; Li et al., 2009; Smyth et al., 2009; Larsen et al., 2012; Ritchie et al., 2014). Although many antimicrobial-resistant S. aureus isolates were identified by phenotypic and genetic methods, the differences in genetic structure among these resistant isolates remain largely unknown.
Knowledge of the AMR patterns and genetic diversity of resistant S. aureus isolates from cases of bovine mastitis will provide pivotal epidemiological information on (1) development and persistence of resistant isolates in dairy farm environment, (2) distribution patterns of resistant clones within and between farms, (3) identification of sources of resistant clones and critical control points in dairy farms, (4) treatment strategies, and (5) development of effective AMR mitigation strategies in dairy farms. Therefore, the objectives of this study were (1) to evaluate the AMR of S. aureus isolates obtained from dairy cows and (2) to analyze genetic diversity of resistant isolates to determine the scope of distribution of AMR traits among S. aureus isolates.
Materials and Methods
Bacterial isolation, identification, and susceptibility testing
A total of 239 S. aureus isolates were isolated from 187 dairy cows with mastitis from 33 dairy farms in Tennessee during 2004–2016. The isolates were identified and stored by Tennessee Quality Milk Laboratory at the University of Tennessee, Department of Animal Science, Knoxville, TN. S. aureus isolates were tested for their sensitivity to a panel of 10 commonly used antimicrobials for the treatment of mastitis. All milk samples were collected and microbiologically analyzed following the National Mastitis Council guidelines (Boerlin et al., 2003; Oliver et al., 2004). Isolates were tested for their susceptibility against ampicillin, penicillin, erythromycin, oxacillin +2% NaCl, pirlimycin, penicillin–novobiocin (P/N) combination, tetracycline, cephalothin, ceftiofur, and sulfadimethoxine using the Sensititer system (ThermoFisher Scientific, Cleveland, OH) with broth microdilution method according to Clinical Laboratory Standard Institute (CLSI, 2015, 2016) guidelines. The antibiotic concentration ranges on the Sensititer plates are shown in Table 1. Minimum inhibitory concentrations (MICs) of the antimicrobials were determined as described elsewhere (Saini et al., 2011). The MIC was defined as the lowest concentration of antimicrobial that prevents visible bacterial growth after incubation for 18 h at 37°C.
The MICs were interpreted according to CLSI (2015) M31-A3.
MICs, minimum inhibitory concentrations.
Molecular fingerprinting of the S. aureus isolates by PFGE
Molecular fingerprinting of S. aureus isolates was done using PFGE as described (McDougal et al., 2003; Gillespie et al., 2009; Sawant et al., 2009) with minor modifications. In brief, each bacterial isolate was inoculated and incubated overnight on blood agar plate. A single pure colony was inoculated to 5 mL of Brain Heart Infusion broth and incubated at 37°C for 24 h. Bacterial concentration was adjusted to an OD600 of 0.9 to 1.1 with phosphate-buffered saline using spectrophotometer (Bio-Rad, Hercules, CA). A 200 μL aliquot of the culture was pelleted and resuspended in 300 μL of Tris-EDTA buffer (pH 8.0). The suspension was mixed with 1.8% (w/v) InCert agarose (Lonza, Rockland, ME) in Tris-EDTA buffer (ThermoFisher Scientific), dispensed into the wells of disposable plug mold (Bio-Rad) and digested with lysostaphin (1 mg/mL in 20 mM sodium acetate, pH 4.5; Sigma Aldrich, St. Louis, MO). The plugs were washed four times in 4 mL of Tris-EDTA buffer at 37°C for 20 min and further digested with SmaI enzyme (McDougal et al., 2003; Gillespie et al., 2009; Sawant et al., 2009).
Chromosomal DNA digestion by SmaI enzyme
Agar plugs were cut into a 2 × 2 mm size, equilibrated in 1 × SmaI restriction buffer for 30 min, and digested with SmaI (New England BioLabs, Inc., Ipswich, MA) in a total volume of 200 μL (3 μL SmaI + 197 μL of 1 × buffer) at 25°C for 3 h. A single plug was loaded on to each tooth of 15 combs with the control S. aureus strain NCTC 8325 was loaded into the 1st, 8th, and 15th combs and incubated at room temperature for 20 min. The comb was placed in the gel-casting platform and the 1% SeaKem agarose was poured into it and kept at room temperature for 20 min until solidified. The gel electrophoresis was conducted using the CHEF Mapper at initial switch of 5 s, final switch of 40 s, and running time for 21 h at 200 V (6 V/cm) at temperature of 14°C using ramp angle of 120°. The gel was stained by ethidium bromide (1.25 μg per mL of water; Invitrogen, Carlsbad, CA) for 25 min and washed twice for 30 min with fresh distilled water. Images were taken by ChemiDoc® Touch Imaging System (Bio-Rad) and exported to PulseNet by Image Lab 5.2.1 Software (Bio-Rad) and saved as a TIFF file.
Data analysis and interpretation
S. aureus isolates were classified as susceptible or resistant according to the interpretative criteria (CLSI, 2015) of the MIC values (Table 1). Intermediate values were considered resistant. The prevalence of AMR isolates was compared in two time intervals (2004–2011 and 2012–2016).
The TIFF images of PFGE were imported and analyzed using GelCompar II software version 6.6 (Applied Maths, Kortrijk, Belgium). The intra- and intergel PFGE runs were normalized using control S. aureus strain NCTC8325. The bands ranging between 10 and 674 kb were used for analysis. The band patterns of the isolates were used for grouping as follows: (1) same strains (no band difference), (2) closely related strains (differed by two to three bands), (3) possibly related strains (differed by four to six bands), and (4) unrelated strains (differed by seven or more bands). PFGE types and subtypes were defined by groups formed at seven or more band differences (to define PFGE types) and four to six bands differences (to define PFGE subtypes) (Tenover et al., 1995). For band pattern comparisons within and between different gels for isolates, the following settings were used: optimization of 0.5% and position tolerance of 1.25%. PFGE types and subtypes were defined by group differed by seven or more bands and four to six bands similarity cutoffs, respectively, on a dendrogram constructed by the unweighted-pair group matching algorithm (UPGMA) (Faria et al., 2008).
Results
Prevalence of antimicrobial-resistant S. aureus
The prevalence of AMR among the 239 isolates tested is shown in Figure 1. In total, 82 (34.3%) isolates were resistant to one or more of the antimicrobials. Overall, AMR among S. aureus isolates varied from as low as 1.3% for ceftiofur to as high as 25% for sulfadimethoxine. The prevalence of AMR was widely distributed throughout the farms at a varying proportion for different antimicrobials tested. About two-third of the farms had sulfadimethoxine-resistant isolates, one-third had penicillin and erythromycin, a quarter had ampicillin, penicillin–novobiocin, and one-fifth had tetracycline-resistant isolates (Fig. 2). Seven (21.2%) out of the 33 farms did not have AMR S. aureus isolates, whereas 26 (78.8%) of the farms had at least one S. aureus isolate resistant to one or more drugs. Sulfadimethoxine-resistant S. aureus were the most widely distributed resistant isolates across 21 (63.6%) farms followed by penicillin-resistant isolates in 12 (36.4%) of the 33 dairy farms (Fig. 2). The prevalence of AMR declined from 2004 to 2016 for all antimicrobials except for tetracycline resistance (Fig. 3). However, the significance of the time on AMR was not evaluated statistically due to small sample size.
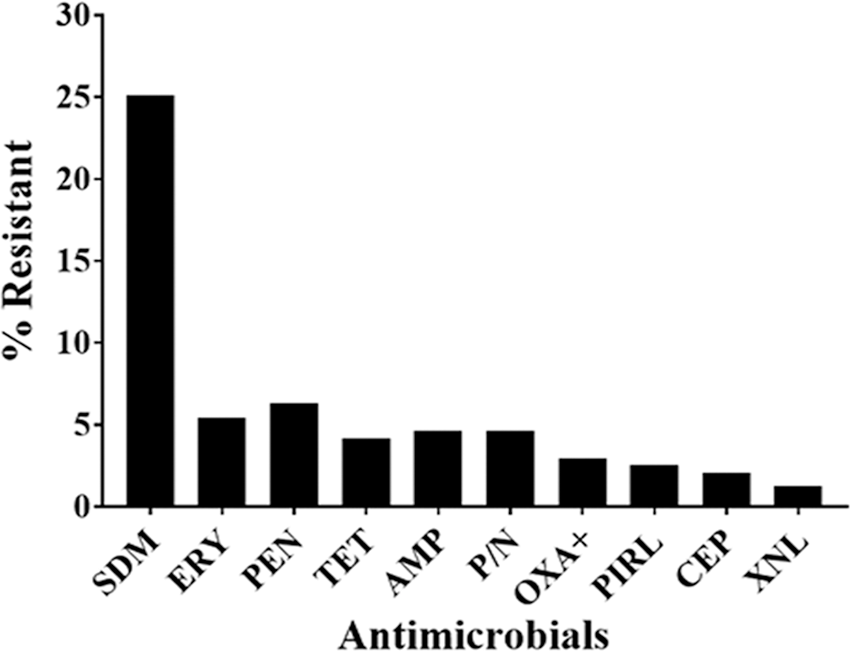
Prevalence of antimicrobial-resistant Staphylococcus aureus isolates against commonly used antimicrobials in dairy cattle. Out of 239 S. aureus isolates, the prevalence of resistance to ampicillin (AMP) was 4.6% (n = 11), penicillin (PEN) 6.3% (n = 15), erythromycin (ERY) 5.4% (n = 13), oxacillin +2% NaCl (OXA+) 2.9% (n = 7), pirlimycin (PIRL) 2.5% (n = 6), penicillin/novobiocin (P/N) 4.6% (n = 11), tetracycline (TET) 4.2% (n = 10), cephalothin (CEP) 2.1% (n = 5), ceftiofur (XNL) 1.3% (n = 3), and sulfadimethoxine (SDM) 25% (n = 60).
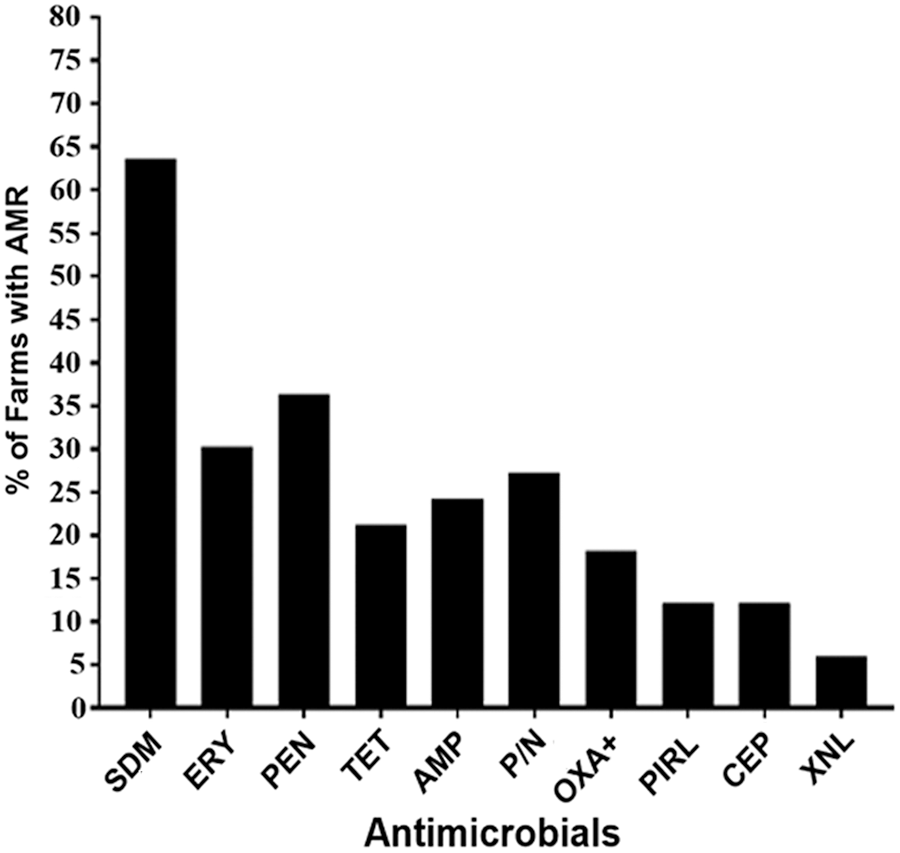
The distribution of antimicrobial-resistant Staphylococcus aureus isolates among 33 dairy farms. Out of 33 dairy farms, 8 (24.2%) had ampicillin (AMP), 12 (36.4%) penicillin (PEN), 10 (30.3%) erythromycin (ERY), 6 (18.2%) oxacillin +2% NaCl (OXA+), 4 (12.1%) pirlimycin (PIRL), 9 (27.3%) penicillin/novobiocin (P/N), 7 (21.2%) tetracycline (TET), 4 (12.1%) cephalothin (CEP), 2 (6.1%) ceftiofur (XNL), and 21 (63.6%) sulfadimethoxine (SDM)-resistant isolates.
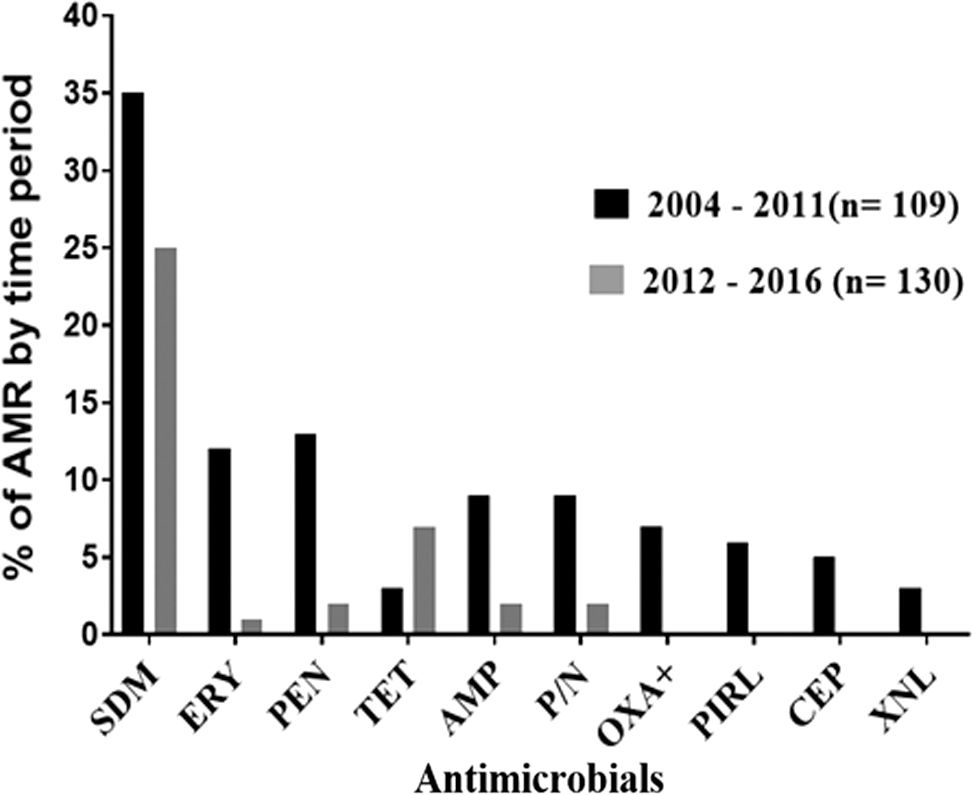
Antimicrobial resistance among Staphylococcus aureus isolates over two time periods (2004–2011 and 2012–2016). Of 109 isolates during 2004–2011, the prevalence of resistance to ampicillin (AMP) was (8.3%), penicillin (PEN) (11.9%), erythromycin (ERY) (11%), oxacillin (OXA+) (6.4%), pirlimycin (PIRL) (5.5%), penicillin–novobiocin (P/N) (8.3%), tetracycline (TET) (2.8%), cephalothin (CEP) (4.6%), ceftiofur (XNL) (2.8%), and sulfadimethoxine (SDM) (32.1%). Of 130 isolates during 2012–2016, resistance to AMP was (1.5%), PEN (1.5%), ERY (0.8%), OXA+ (0%), PIRL (0%), P/N (1.5%), TET (5.4%), CEP (0%), XNL (0%), and SDM (19.2%). The prevalence of SDM-resistant isolates was significantly higher during the 2004–2011 period compared with the 2012–2016 period (95% odds ratio = 1.011–4.235; p < 0.05).
AMR profiles of S. aureus
The AMR patterns of S. aureus vary among tested antimicrobials. In total, 82 (34.3%) of the S. aureus isolates were resistant to at least 1 of the 10 antimicrobials, whereas 157 (65.7%) of the 239 isolates were pansusceptible. Out of all tested isolates, 25.5% were single drug resistant, 4.6% were double drug resistant, and 4.2% were multidrug resistant. Eighty-two resistant isolates showed 21 different phenotypic patterns. Three-fourths (61; 74.4%) of these resistant isolates showed six patterns of single AMR, whereas 11 (13.4%) of the isolates showed double AMR patterns. The remaining 10 (12.2%) S. aureus isolates were resistant to ≥3 antimicrobials (Table 2). Of the multidrug-resistant isolates, seven (70%) were also oxacillin-resistant isolates. AMR was not detected in 7 (21.2%) of 33 farms. The remaining 26 (78.8%) farms had at least one isolate with single, double, or multiple drug resistance phenotypes. Overall, the 61 isolates with single drug resistance phenotype were obtained from 20 (60.6%) farms. The 11 isolates with double drug resistance phenotypes were obtained from 8 (24.2%) of the 33 farms. Six (18.2%) of the 33 farms had at least one isolate with multidrug resistance (MDR) phenotype. Of the 11 isolates resistant to ampicillin, 10 (90.9%) were also resistant to penicillin (Table 2).
AMP, ampicillin; AMR, antimicrobial resistance; CEP, cephalothin; ERY, erythromycin; OXA+, oxacillin +2% NaCl; P/N, penicillin/novobiocin; PEN, penicillin; PIRL, pirlimycin; SDM, sulfadimethoxine; TET, tetracycline; XNL, ceftiofur.
PFGE analysis
PFGE was conducted on 56 of 82 resistant isolates, showing that 10–16 chromosomal DNA fragments ranging from 10 to 674 kb in size were generated. Forty-six distinct PFGE patterns (clones) were generated from the 56 AMR isolates (Fig. 4). A clone was defined as a set of isolates identical in all banding patterns. Based on estimates of genetic relationship using the UPGMA cluster analysis method, the 46 PFGE patterns were clustered into nine major clonal complexes (PFGE types) that were designated by the letters A through I. The PFGE type I was predominantly observed in 31 (55.4%) of 56 isolates followed by type G, which was detected in 11 (19.6%) of 56 isolates. The PFGE types A, C, E, and H were rare among the tested isolates, since only 1 belongs to each of them out of 56 isolates tested (Fig. 4 and Table 3). Subclonal types under clonal types D, F G, and I were designated with Roman numbers (Fig. 4). Isolates with the PFGE type I were the most complex in genetic diversity and had 11 different subtypes. Those under PFGE type G had seven subtypes. There was a specific PFGE type and AMR patterns distribution among the resistant isolates from different farms (Fig. 4 and Table 3).

PFGE types of antimicrobial-resistant Staphylococcus aureus isolates from 33 dairy cattle farms in Tennessee. A tree calculation (dendrogram) of genetic diversity was constructed using the UPGMA clustering method that is based on average number of different bands of SmaI digested chromosomal DNA to evaluate PFGE patterns of 56 antimicrobial-resistant S. aureus isolates. The S. aureus strain NTTC 8325 was used as control for the experimental variations between duplicate experiments (intra-and intergel variation). Numbers under Staph PFGE indicate band numbers, numbers under farm indicate dairy farm ID, letters under PFGE indicate PFGE types, and Roman numbers indicate PFGE subtypes. PFGE, pulsed-field gel electrophoresis; UPGMA, unweighted-pair group matching algorithm.
AMP, ampicillin; AMR, antimicrobial resistance; CEP, cephalothin; ERY, erythromycin; OXA+, oxacillin +2% NaCl; P/N, penicillin/novobiocin; PEN, penicillin; PFGE, pulsed-field gel electrophoresis; PIRL, pirlimycin; SDM, sulfadimethoxine; TET, tetracycline; XNL, ceftiofur.
Twenty-two (39.3%) out of 56 isolates were detected in four farms. A total of 14 (25%) of 56 isolates with three PFGE types were detected in two farms, and 16 (28.6%) of 56 isolates with four PFGE types were detected in two other farms. A farm with a single lineage was also detected. A total of 4 (7.1%) of 56 isolates with one PFGE type were detected in three farms (Fig. 4 and Table 3). The abundance and distribution patterns of the nine PFGE types in 11 farms showed hierarchy of genetic structure. Of the nine PFGE types, types G and I were abundant and widely distributed in 7 of 11 farms. Both types G and I comprised dominant strains accounting for 42 (75%) of the 56 isolates. Similarly, out of 11 farms, 3 farms had type F and 2 farms had type D. A unique single PFGE type was observed in three separate farms, PFGE types A and B in farm 4, type C in farm 8, and types E and H in farm 2. Four different PFGE types consisting of D, E, H, and I were detected in farm 2, whereas PFGE types comprising C, F, G, and I were detected in farm 8. No farm had more than four PFGE types.
Certain PFGE types were found to be associated with AMR particularly to two PFGE types, that is, I followed by G. An isolate with single PFGE type was found resistant to multiple antimicrobials in this study. In this regard, those from both G and I types were linked with seven AMR phenotypes. The sulfadimethoxine-resistant isolates showed seven PFGE types, that is, B, D, E, F, G, H, and I, indicating multiple PFGE types with a single AMR pattern. Tetracycline-resistant isolates showed F, G, and I PFGE types, whereas erythromycin-resistant isolates grouped into G and I types (Table 2). Changes of PFGE profiles over time indicated that some PFGE types were farm specific such as PFGE type B that was persistently endemic over time in farm 4, whereas some PFGE types such as G and I were distributed overtime between different farms. Interestingly, some subtypes within PFGE type I such as I-iv and I-x were persistently endemic over time in specific farms, particularly in farms 3 and 5, respectively.
Discussion
Evaluation of S. aureus isolated from dairy cows for sensitivity to antimicrobials showed that 34.3% of the isolates were resistant to at least one antimicrobial and the majority (65.7%) of the isolates was pansusceptible. Of the AMR isolates (n = 82), 74.4% showed resistance only to one antimicrobial as compared with 13.4% resistance to two antimicrobials and 12.2% MDR (resistance to more than three antimicrobials). The resistant isolates showed 21 different resistance patterns. These findings indicated that about a third of the S. aureus isolates were AMR and MDR was not apparently widespread. Even though it was not possible to obtain accurate information on antibiotics use patterns in the dairy farms included in this study, occurrence of high-level resistance to single antibiotics and low MDR may indicate variations in the antibiotic selection and use practices. Similar observations were made from Wisconsin dairy cattle in which 25% (n = 116) of S. aureus isolates were AMR, and 0.9% were MDR (Oliveira et al., 2012). However, in that study only five different resistance patterns were reported, suggesting that AMR in the S. aureus tested in our study had more diversity in their resistance profile.
A relatively high prevalence (25%) of sulfadimethoxine resistance was observed among the isolates, which agree with a previous study (Pol and Ruegg, 2007). Higher prevalence (49%) was also reported from Wisconsin (Sato et al., 2004). Sulfadimethoxine is approved in the United States for the treatment of septicemia caused by coliform mastitis, respiratory pathogens, and necrotic pododermatitis (Oliveira and Ruegg, 2014). We did not evaluate antimicrobial usage patterns in each farm, particularly sulfametoxide in treating mastitis (Oliveira and Ruegg, 2014). The high sulfadimethoxine resistance prevalence requires further study to elucidate the level of sulfadimethoxine use in dairy cattle production for treatments other than mastitis, and its association with sulfadimethoxine-resistant S. aureus. Interestingly, resistance to antimicrobials that belong to β-lactams (ampicillin, penicillin, ceftiofur, and cephaprin) and lincosamide (pirlimycin), the two antimicrobial classes most commonly used for the treatment of mastitis in the United States, was generally low. For the β-lactams, it ranged from 1.3% (ceftiofur) and 2.5% (pirlimycin) to 6.3% (penicillin). These findings are similar to reports from other parts of the United States (Pol and Ruegg, 2007; Oliveira et al., 2012; Oliveira and Ruegg, 2014; Ruegg et al., 2015). Resistance to tetracycline (4.2%), erythromycin (5.4%), penicillin–novobiocin (4.6%), and oxacillin +2% NaCl (2.9%) was generally comparable with reports from Wisconsin (Oliveira et al., 2012), but lower than a more recent study in Wisconsin (Ruegg et al., 2015). Oxacillin +2% NaCl is not prescribed for veterinary use, particularly for mastitis in the United States. The 2.9% oxacillin resistance observed in this study is comparable with the 2.1% reported for Minnesota dairy herds (Haran et al., 2012). Some authors suggest that oxacillin resistance is indicative of MRSA (Aarestrup and Schwarz, 2006) with potential public health implication. In this study, all the oxacillin-resistant isolates were MDR (Table 2), supporting the idea that methicillin/oxacillin resistance is associated with multidrug-resistant S. aureus (Chambers and Deleo, 2009).
AMR in S. aureus isolates detected in this study was broadly distributed across the 33 farms. Anderson et al. (2006) reported a 12.5% (3 of 24 farms) prevalence in North Carolina and Virginia, which was lower than that in this study. In general, specific antibiotic resistances were widely distributed (Fig. 2). Sulfadimethoxine resistance was the most widespread (occurred in 63.6% of the farms), and ceftiofur resistance was the least widespread (6% of the farms). These results revealed that the percentage of farms that had resistant isolates varied among the 10 antimicrobials tested. It is possible that different dairy farms use different antimicrobials and hence the percentage of resistant isolates was expected to vary with farm and type of commonly used antibiotics (Saini et al., 2012b; Oliveira and Ruegg, 2014).
The prevalence of antimicrobial-resistant S. aureus showed some variations over the years. With the exception of tetracycline resistance, which increased during the 2012–2016 period, resistance to all other antibiotics declined from 2004–2011 to 2012–2016 (Fig. 3). A decreasing trend for penicillin from 49% in 1994 to 30% in 2001 (Makovec and Ruegg, 2003) to 20% in 2015 (Ruegg et al., 2015) was reported in Wisconsin. Over the 6-year period, from mid-1990 to early 2000, S. aureus isolates from bovine mastitis showed either a decreasing trend or did not change their AMR level (Erskine et al., 2002). Despite the fact that in this study we did not evaluate the 33 farms over time, the AMR profile of isolates in recent years seems lower than in previous years. These could be due to several reasons, including (1) rising awareness of farmers about prudent use of antimicrobials, (2) aggressive culling of chronically infected cows, showing no response to antibiotic treatments, (3) changes in treatment protocols and intensity of drug use in dairy farms, (4) increasing regulation and availability of guidelines on antimicrobial usage from FDA, and continuous monitoring of milk quality standard from milk buyers and dairy farmers association. Despite a general trend of reduction in AMR for most antibiotics over recent years, we noticed that resistance to tetracycline increased during 2012–2016 than during the previous years. This might be due to increased usage of tetracycline as the most preferred antibiotic to treat other health problems of dairy cows such as lameness, pneumonia, and reproductive infections (USDA APHIS, 2008a; Oliver et al., 2011). Therefore, variation in the prevalence of AMR among farms and over years emphasizes the need for continuous monitoring of AMR at the farm level overtime to determine effective mitigation measures.
Among the 56 AMR S. aureus isolates tested, 46 PFGE patterns were observed that were clustered into 9 major PFGE types, suggesting genetic diversity of the isolates. Studies from different parts of the world indicate that S. aureus isolates obtained from dairy cows tend to be genetically diverse (Kapur et al., 1995; Fitzgerald et al., 1997; Zadoks et al., 2002; Sabour et al., 2004; Anderson et al., 2006; Li et al., 2009). Lineage I that occurred in 55.4% of the resistant isolates (n = 56) was the most prevalent PFGE type followed by lineage G that occurred in 19.6% of the resistant isolates. PFGE types D and F together accounted for 14.2% (7.1% each) of the 56 isolates tested and the remaining PFGE types (A, B, C, E, and H) were less abundant. The predominance of PFGE type I may suggest clonal expansion of the isolates with this PFGE type between and within the farms. Previous reports support this observation (Anderson et al., 2006; Sakwinska et al., 2011). The PFGE typing showed that multiple lineages coexisted in a particular farm. Of the 56 resistant isolates tested, two lineages coexisted in four farms, three lineages coexisted in two farms, and four lineages coexisted in another two farms, accounting for 39.3% (22/56), 25% (14/56), and 28.6% (16/56) of the resistant isolates tested, respectively. This indicates that majority (92.9%) of the resistant isolates coexisted in multiple farms. Our findings and other reports from Pennsylvania and Ohio (Kapur et al., 1995) showed that although few lineages dominate, the S. aureus population in a particular farm/herd can be multiclonal. No farm was found to have more than four lineage types. Results of this study showed that the types of lineages involved in coexistence varied with farm. Coexistence of the heterogeneous lineages that accounted for AMR of 41.1% within a specific farm has been reported from Canadian dairy herds (Sabour et al., 2004). Existence of a single lineage was detected in three farms, accounting for AMR of 7.1% (4/56). Single lineage existence per farm accounting for 58.6% of the isolates in Canada (Sabour et al., 2004) and in Pacific regions (Ritchie et al., 2014) was reported.
Interestingly, the phenotypic assay discriminated resistant S. aureus isolates into 21 AMR patterns, whereas PFGE displayed 46 patterns. This indicated a better discriminatory power of PFGE than the phenotypic AMR assay method (Zadoks et al., 2002; McDougal et al., 2003; Hallin et al., 2007). Our results showed that the majority of the AMR patterns were linked to a few PFGE types, particularly G and I. The G and I PFGE types were linked to multiple (seven) AMR patterns, thus indicating that these are the major lineages responsible for AMR in the study area. Similarly, reports from different geographical areas indicated that frequently one or a few clones of S. aureus are responsible for resistance to antimicrobials (Anderson et al., 2006; Chambers and Deleo, 2009; Sakwinska et al., 2011). In contrast, majority of the S. aureus genotypes detected in this study seem to acquire resistance to sulfadimethoxine. Sulfadimethoxine resistance was widely disseminated in seven PFGE types (B, D, E, F, G, H, and I) of the nine lineages detected, indicating multiple lineages had resistance to a single antimicrobial drug. Tetracycline resistance was detected in F, G, and I PFGE types and erythromycin resistance was observed in G and I PFGE types.
Conclusion
The prevalence of antimicrobial-resistant S. aureus isolates from dairy cows varied with farms, time, and types of antimicrobials tested. With the exception of sulfadimethoxine, resistance to most antimicrobials was low and there was gradual decline in resistance to commonly used antimicrobials such as cephalothin, ceftiofur, and pirlimycin from 2004–2011 to 2012–2016. AMR S. aureus showed both phenotypic and genotypic diversity, and most AMR patterns were associated with few PFGE types. Studies are required to further understand the widespread occurrence of sulfadimethoxine and tetracycline resistance in S. aureus in dairy cattle production systems despite the limited use of these antibiotics for the treatment of bovine mastitis.
Footnotes
Acknowledgment
This project was funded by the University of Tennessee, College of Veterinary Medicine, Center of Excellence in Livestock Diseases and Human Health (UT CVM-COE).
Disclosure Statement
No competing financial interests exist.