
Abstract
Enteropathogen colonization and product contamination are major poultry industry problems. The emergence of antibiotic resistance, and associated risks to human health, is limiting the use of antibiotics as first-line defense against enteropathogens in poultry. The chitin derivative, chitosan, has drawn substantial attention for its bactericidal properties. Different molecular weight (MW) chitosans can have varied effects against different bacteria in monoculture. In the current study, cecal contents from each of three market-age broilers and Salmonella Typhimurium, as indicator enteropathogen, were exposed to in vitro anaerobic culture to three chitosan preparations (0.08%, wt/vol), low (LMW), medium (MMW), and coarse (CMW). Effects of chitosan and the carrier solvent acetic acid, on cecal bacteria and Salmonella, were examined by denaturing gradient gel electrophoresis (DGGE) and Salmonella enumeration. Bacterial profiles for the three cecal contents were shown by DGGE to be very different. Each of the three cecal contents grown in the presence of 0.08% acetic acid was very different from the same contents grown without the chitosan solvent. Culturing cecal contents in the presence of chitosan altered the bacterial DGGE profiles from the control and acetic acid-only cultures. The DGGE chitosan-treated profiles for all three cecal sources were identical to each other regardless of the MW chitosan in the culture medium. Compared with Salmonella in monoculture, Salmonella decreased (p < 0.05) by about 1.5 log CFU/mL when grown in mixed culture with cecal contents. Salmonella monocultures in the presence of 0.08% of the chitosan solvent acetic acid decreased (p < 0.05) counts by almost 3.5 log CFU/mL. Combining acetic acid and cecal contents reduced (p < 0.05) Salmonella by 7 log CFU/mL. Adding the chitosan preparations to the mixtures reduced (p < 0.05) Salmonella by 8 log CFU/mL.
Introduction
A
Conflicting reports indicate the effectiveness of chitosan based on molecular weight (MW) (No et al., 2002; Zheng and Shu, 2003; Liu et al., 2006; Raafat et al., 2008; Mellegard et al., 2011). Raafat et al. (2008) and Zheng and Shu (2003) gave conflicting effects of MW on Staphylococcus aureus. No et al. (2002) reported increasing Gram-negative bacterial death with decreasing MW, whereas a similar effect was not seen with Gram-positive bacteria. Jeon et al. (2001) reported chitosan as more effective against pathogens than nonpathogens.
Broiler feed supplemented with 3% chitosan were highly resistant to Salmonella Gallinarum infection (Balicka-Ramisz et al., 2007). Chitosan at 0.2% significantly reduced Salmonella Typhimurium in broiler ceca (Menconi et al., 2014). Chitosan of different MW have varied effects on different bacteria. The objective of the current report was to determine in vitro if three different chitosan MW preparations had different effects on cecal bacterial profiles as examined by denaturing gradient gel electrophoresis (DGGE). Salmonella was included as an antienteropathogen (Salmonella) indicator.
Materials and Methods
Bacterial strain
A Salmonella Typhimurium was previously cultured from a naturally infected broiler and maintained in frozen stock as experimental inoculum. The Salmonella Typhimurium was previously selected for resistance to nalidixic acid (minimum inhibitory concentration [MIC] ≥16 mg/L) (Sigma Chemical Company, St. Louis, MO) for mixed-culture recovery with selective media.
Frozen (80°C) stock was streaked onto Brilliant Green Agar (BGA; Difco; Becton, Dickinson and Company, Sparks, MD), containing 20 mg/L nalidixic acid and 25 mg/L novobiocin (Salmonella Typhimurium are naturally resistant to novobiocin and novobiocin inhibits background growth, facilitating enumeration), and cultured aerobically (5% carbon dioxide, 5% hydrogen, 80% nitrogen; Coy Laboratory Products, Grass Lake, MI) overnight at 40°C (broiler body temperature). Five colonies were transferred to 5 mL of cation-adjusted Muller-Hinton broth (MHB; Difco) and cultured anaerobically overnight for three passes before use.
In preliminary experiments (data not shown), Salmonella Typhimurium monoculture MIC was 0.04% to 0.06% for LMW and 0.6% to 0.07% for MMW and CMW. Chitosan solutions (1%) were prepared in 1% acetic acid. Bactericidal effects of acetic acid in Salmonella monoculture began to be seen at 0.07% to 0.08% and the acetic acid MIC was 0.12% to 0.16%, with 0.16% acetic acid consistently killing all Salmonella. Therefore, 0.08% chitosan was used to examine the effects of the three MW chitosans on anaerobic cecal cultures.
Cecal contents
Cecal contents from each of three market-age broilers were used in in vitro experiments to analyze treatment effects on more uniform bacterial populations, in contrast to using different cecal contents from a variety of live birds for each treatment. Ceca were aseptically excised from three (n = 3) on-feed 6-week-old market-age Cobb × Ross broiler females (Sanderson Farms, Bryan, TX), then transferred to the anaerobic chamber (Coy). Cecal contents from each broiler were expressed into separate sterile beakers and mixed (1:1, wt/vol) thoroughly in sterile 40% glycerol in Butterfield's buffer (0.62 mM KH2PO4, pH 7.2). Aliquots of contents were stored at −80°C until used.
Chitosan
Three chitosan preparations (Sigma Chemical Company) were used in these experiments: low MW (LMW, 50–190 kDa, Cat. No. 448869), medium (MMW, 190–310 kDa, Cat. No. 448877), and coarse (CMW, 310–375 kDa, Cat. No. 419419). Stock preparations (1% wt/vol) were mixed into solution in 1% (vol/vol) acetic acid (Sigma), sterilized by autoclaving (Roller and Covill, 1999; Raafat et al., 2008), and stored at 4°C until used. A 1% acetic acid stock solution without chitosan added served as a control treatment.
All experiments were conducted in cation-adjusted MHB. The absence of chitosan precipitation with cation-adjusted MHB at one or two pH units below neutral, unlike chitosan precipitation seen with other Salmonella-compatible growth media at near neutral pH (unpublished), made it highly suitable for these experiments (Raafat et al., 2008; Dina Raafat personal communication). Each 1% chitosan and acetic acid control preparation was diluted in MHB to 0.16% as starting working solutions.
In vitro anaerobic culture
Experiments were conducted in three separate trials and cecal contents in each treatment group were run in triplicate in each trial. All Salmonella inocula and experiments were cultured at 40°C to mimic broiler body temperature. Overnight anaerobic Salmonella cultures grew to ∼108 CFU/mL and the final inoculum level in each assay tube was ∼105 CFU/mL.
Components were combined to provide the following treatments: no acetic acid and no cecal contents control (Exp Sal Con); 0.08% acetic acid and no cecal contents control (AA Sal Con); cecal contents control and no acetic (No AA Con); cecal contents plus 0.08% acetic acid solvent control (AA Con); and each chitosan preparation at 0.08% final concentrations of low (LMW), medium (MMW), and coarse (CMW) MW chitosans. All tubes were incubated 18–24 h, anaerobically at 40°C. One milliliter from each culture was 10-fold serially diluted, plated on BGA, and incubated 18–24 h, aerobically at 40°C. Colony counts were expressed as log10 CFU/mL (referred to as CFU/mL and mean CFU/mL ± SD). One milliliter of cultures from AA Con, LMW, MMW, and CMW was added to 10 mL of Rappaport-Vassiliadis R10 Broth (RV; Difco), incubated aerobically overnight at 42°C for Salmonella enrichment, and streaked onto BGA followed by incubation overnight at 40°C.
Samples of Salmonella negative following serial dilution and plating, but positive following RV enrichment, were given an arbitrary 0.30 log10 CFU/mL for Salmonella. Colony counts (log10 CFU/mL) for the three trials for each of the cecal contents were similar and were combined (n = 9) for comparison using ANOVA and mean separation by Duncan's multiple range test (SAS 9.4 for Windows; SAS Institute Inc., Cary, NC).
DGGE and pH
One milliliter of each culture was centrifuged at 8000 × g for 10 min. Pellets were suspended in 180 μL of lysis buffer (the QIAmp DNA Mini Kit; QIAgen Incorporated, Valencia, CA) containing 20 mg/mL of lysozyme (Sigma) and DNA was isolated as instructed. Bacterial 16S rDNA genes were PCR amplified using 30 ng of sample DNA and DGGE was conducted as previously described (Muyzer et al., 1993; Hume et al., 2003). Dendrogram band pattern relatedness was analyzed using GelCompare II, v6.6 11 (Applied Maths, Austin, TX) based on the Dice similarity coefficient and the unweighted pair group method using arithmetic averages (UPGMA) for cluster analysis and comparison according to percentage similarity coefficient (%SC). Supernatants were used to determine culture pH and treatment mean were compared using ANOVA and mean separation by Duncan's multiple range test (SAS 9.4 for Windows, SAS Institute Inc., Cary, NC).
Results and Discussion
Denaturing gradient gel electrophoresis
Comparisons by DGGE were based on the presence of identifiable bands within each profile. Band intensity was not a consideration in these comparisons, since PCR results can be biased by a number of sample and solution features (Suzuki and Giovannoni, 1996; Chandler et al., 1997; Wintzingerode et al., 1997; Kanagawa, 2003). The main consideration was whether treatment resulted in the appearance or elimination of specific bacterial bands.
Cecal bacteria DGGE profiles segregated mainly according to treatment with No AA Con sharing 68.3%SC, AA Con sharing a 66.7%SC, and all the chitosan groups sharing a 95.5%SC, which indicated the chitosan groups as essentially sharing the same profile (Fig. 1). Profiles for the three cecal samples in the No AA Con groups separated from each other as did the three cecal samples within the AA Con group. This result was expected due to the high variability between digestive microbial contents found in different broilers, even those broilers within the same treatment group. The 79.5%SC for cecal sample 2 and 3 and the 68.3%SC between these two samples and cecal sample 1 indicate that the three cecal contents shared no initial profile similarities. Additionally, no profile similarities were seen for the three cecal samples given the AA Con treatment. The effect of acetic acid on cecal bacteria can be seen in the degree of separation (60.1%SC) of profiles from the No AA Con group and the AA Con group.

Dendrogram of broiler in vitro anaerobic cecal bacteria plus Salmonella cultures following denaturing gradient gel electrophoresis PCR analysis of cecal contents from broilers 1, 2, and 3 and with three replicate samples for each treatment. Treatments: No acetic acid+cecal contents control (No AA Con), acetic acid+cecal contents control (AA Con), low-molecular-weight chitosan+cecal contents (LMW), medium-molecular-weight chitosan+cecal contents (MMW), coarse-molecular-weight chitosan+cecal contents (CMW).
Deleterious effects of organic and nonorganic acids on bacteria are well documented (Levine and Fellers, 1940; Eklund, 1983; Hinton and Linton, 1988; Cherrington et al., 1991; Abdul-Roauf et al., 1993; Huyghebaert et al., 2011). Although acetic acid at 0.08% as used in these experiments was well below the MIC of 0.16%, the acetic acid solvent had a tremendous antibacterial effect as determined by Salmonella cell count and DGGE. However, a greater change in Salmonella numbers and DGGE profiles was seen in all groups treated with chitosan. There was no profile similarity (54.9%SC) between the three chitosan treatment groups and the control groups, No AA Con and AA Con.
The DGGE comparison showed that chitosan changed the profiles from the three cecal content sources to resemble each other. Additionally, cecal contents from the three sources were not segregated into clads based on source as seen with the No AA Con and the AA Con treatments. The expected outcome was differences in how the low, medium, and coarse MW chitosan affected the three bacterial populations based on their varied bactericidal effect against different bacteria in monoculture (Jeon et al., 2001; No et al., 2002; Zheng and Shu, 2003; Liu et al., 2006; Raafat et al., 2008; Ganan et al., 2009; Kong et al., 2010). Contrary to expectations, the outcomes for the three chitosans were similar in the extent to which they affected general cecal bacterial populations as well as their bactericidal effects on in vitro Salmonella reduction.
Culture pH
Bacterial inhibition by chitosan is reported to increase with lowering pH (No et al., 2002). Chung et al. (2004) demonstrated greater chitosan adsorption on bacteria with decreased solution pH, whereas adsorption with decreasing pH was related to the increased chitosan-positive charge and the increased attraction to cell surface-negative charges.
Supernatant pH for cultures per treatment in each of the three trials for each of the cecal contents were similar and were combined (n = 9) for comparisons. Mean pH for the two acetic acid control cultures, AA Sal Con and AA Con, were the lowest (p < 0.05) for all cultures (Fig. 2). Mean pH values were 5.42, 5.44, and 5.46 for all LMW, MMW, and CMW, respectively, whereas those for AA Sal Con and AA Con were 4.71 and 4.99, respectively. Although the pH values within these two groups are significant, the small differences in pH values in a biological sense within the two groups are likely insignificant.

Culture pH (Mean ± SD) for individual (n = 9) and pooled cecal (n = 27) sources in trials 1, 2, and 3. Exp Sal Con = Salmonella control with no acetic acid and no cecal contents, AA Sal Con = Salmonella control plus acetic acid and no cecal contents, No AA Con = cecal contents control and no acetic acid, AA Con = cecal contents control plus acetic acid, LMW = cecal contents plus low-molecular-weight chitosan in acetic acid, MMW = cecal contents plus medium-molecular-weight chitosan in acetic acid, CMW = cecal contents plus coarse-molecular-weight chitosan in acetic acid. All Ceca treatments with the different letters are significantly different (p < 0.05).
Low pH in the AA Sal Con and AA Con cultures likely contributed to decreased Salmonella growth. Acetic acid, as are other volatile fatty acids (VFA), is bactericidal with Gram-negative bacteria. One mode which affects bacteria is thought to occur from undissociated VFA in the acid environment diffusing across the cell membrane, losing the carboxyl proton once in the cytoplasm, and the newly charged molecule contributing to disruption of cell function (Cherrington et al., 1991).
Culture pH for AA Con is slightly higher (p < 0.05) than that for AA Sal Con. However, Salmonella numbers were lower (p < 0.05) by at least 3 log for the three cecal contents (Fig. 3). These lower cell counts may suggest that the cecal contents had a more negative effect on Salmonella viability. Although pH values for cultures containing chitosan were higher than those for AA Con, Salmonella CFU/mL were lower for cecal contents 1 and 3 than in respective AA Con treatments. An assessment of all trials' pH values (Fig. 2) for the three chitosan groups revealed they were about pH 0.45 more than that for AA Con, whereas CFU/mL were about one log less (Fig. 3).
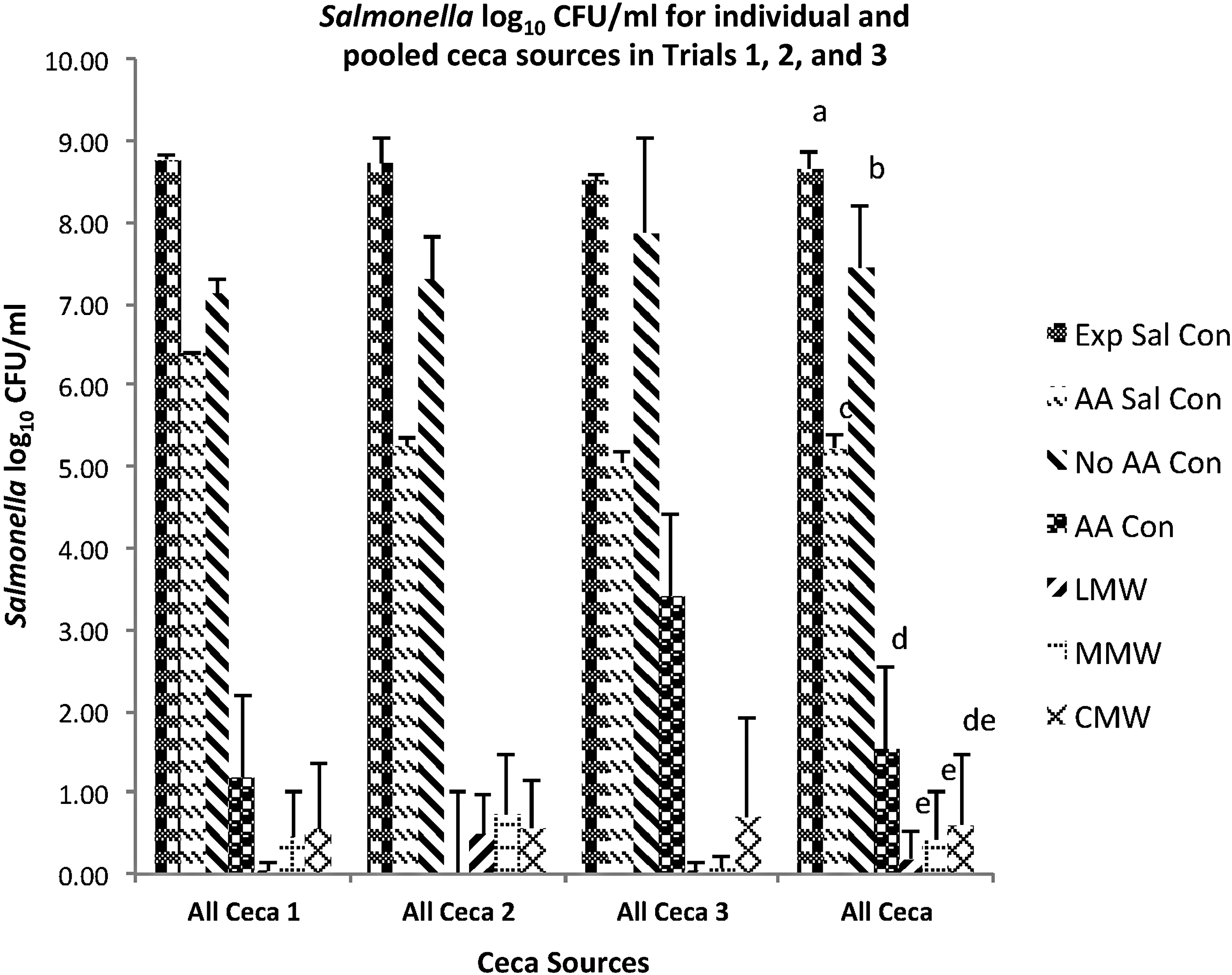
Salmonella log10 CFU/mL for individual and pooled cecal sources in trials 1, 2, and 3. (Mean ± SD) for individual (n = 9) and pooled cecal (n = 27) sources in trials 1, 2, and 3. Exp Sal Con = Salmonella control with no acetic acid and no cecal contents, AA Sal Con = Salmonella control plus acetic acid and no cecal contents, No AA Con = cecal contents control and no acetic acid, AA Con = cecal content control plus acetic acid, LMW = cecal contents plus low-molecular-weight chitosan in acetic acid, MMW = cecal contents plus medium-molecular-weight chitosan in acetic acid, CMW = cecal contents plus coarse-molecular-weight chitosan in acetic acid. All Ceca treatments with the different letters are significantly different (p < 0.05).
Salmonella CFU/mL
Salmonella was included in the study as an antienteropathogen indicator of the chitosan preparations in these mixed bacterial population anaerobic cultures. From preliminary MIC results, it was obvious that chitosan had a greater bactericidal on Salmonella in monoculture than what was seen in mixed cecal culture. Chitosan levels could not be increased in these in vitro cultures due to the need for putting chitosan into an acetic acid solution to exhibit antienteropathogen effects and the bactericidal effects of acetic acid at higher levels. This anti-Salmonella solvent effect did not come into play as demonstrated by Balicka-Ramisz et al. (2007), in their very limited study, when Salmonella Gallinarum was eliminated in broilers given feed containing 3% chitosan.
Salmonella inocula for the three trials had a mean of 5.50 CFU/mL and mean cell numbers in Salmonella Typhimurium control (Exp Sal Con) for the three trials following overnight incubation of 8.66 CFU/mL (Fig. 3). Acetic acid is known to have bacteriostatic and bactericidal effects on Salmonella and other bacteria (Goepfert and Hicks, 1969; Cherrington et al., 1991; McHan and Shotts, 1993; Van Immerseel et al., 2006). Culturing Salmonella in monoculture in the presence of 0.08% acetic acid (AA Sal CON) resulted in Salmonella counts nearly 3.5 log values lower (p < 0.05) compared with the Exp Sal Con. Salmonella counts in AA Sal Con for Ceca 3 in Figure 3 were statistically lower than those for Ceca 1 and Ceca 2; however, the difference likely has no biological significance, since the counts are within approximately a 1 log value.
The Nurmi concept, also known as competitive exclusion, competitive inhibition, and the barrier effect (Nurmi and Rantala, 1973; Lloyd et al., 1977; Impey et al., 1987), addresses the inhibition and exclusion of Salmonella colonization by broiler-beneficial digestive and fecal bacteria. Salmonella CFU/mL in cecal contents 1 and 2 for the treatment No AA Con decreased by nearly 1.5 log values (p < 0.05) when compared with the Exp Sal Con (Fig. 3). These decreases were likely a competitive effect of beneficial bacteria against Salmonella. However, there was no difference (p > 0.05) in Salmonella CFU/mL in cecal contents 3 compared with Exp Sal Con.
The lack of complete Salmonella exclusion in the No AA Con by cecal bacteria and the lack of consistent exclusion among the three cecal bacteria sources are indicative of the variability possible when working with competitive exclusion preparations. The inclusion of acetic acid in the mixed cultures (AA Con) resulted in Salmonella decreases (p < 0.05) of 8.58 and 5.09 log CFU/mL with cecal contents 1 and 3, respectively. No Salmonella was detected following serial dilution spread plating and after enrichment for cecal contents 2 given the AA Con treatment. These dramatic decreases in Salmonella counts seen with the combined acetic acid and cecal bacteria may suggest that acetic acid created an environment more conducive to anti-Salmonella bacteria. Adding LMW, MMW, and CMW chitosan to the culture mixtures reduced Salmonella by ∼8 log values below those in the Exp Sal Con. The LMW, MMW, and CMW treatment groups did not statistically differ from the AA Con. Additionally, there were no consistent differences (p > 0.05) among the three cecal contents in Salmonella reduction for the chitosan preparations.
Conclusions
Bacterial DGGE profiles for the three cecal contents were very different from each other. The addition of acetic acid solvent to cecal contents control cultures shifted profiles from those of cecal contents grown without acetic acid and profiles continued to be very different from each other. This result cannot rule out a combined effect of chitosan with acetic acid as the chitosan solvent in these experiments. Culturing the contents in the presence of chitosan shifted profiles from those of the control cultures. However, profiles from all three cecal sources were identical regardless of the MW chitosan in the culture medium. Cecal contents alone from two broilers decreased Salmonella by about 1.5 log CFU/mL when compared with the control Salmonella in monoculture. The LMW and MMW chitosan in some trials resulted in the reduction of Salmonella to undetectable levels. Salmonella at undetectable levels in those trials contributed to the low mean values for LMW and MMW treatments. The three chitosans were very effective in reducing Salmonella counts. Results of these in vitro experiments will be translated in vivo with Salmonella in broilers.
Footnotes
Acknowledgment
No outside funding supported this research.
Disclosure Statement
No competing financial interests exist.