
Abstract
Campylobacter jejuni and Campylobacter coli play a major role in bacteria-related foodborne illness in humans. Recently, a newly identified species, Campylobacter hepaticus, was shown to be the causative agent of spotty liver disease in chickens. The pathogenic potential of C. hepaticus in humans is unknown. This new species contains genes usually used to detect C. jejuni and C. coli in DNA-based detection methods, such as the hippuricase (hipO) gene and the glyA (serine hydroxymethyltransferase) gene, with a high degree of similarity. Therefore, polymerase chain reaction (PCR) primers used to detect these species need to be evaluated carefully to prevent misidentification of these important Campylobacter species. A multiplex PCR was developed and optimized to simultaneously and specifically identify the presence of C. jejuni, C. coli, and C. hepaticus in chicken samples containing high-complexity microbiota. The assay represents a new diagnostic tool for investigating the epidemiology of Campylobacter colonization in poultry and environmental samples. It may also be applicable to the investigation of Campylobacter contamination in food and in outbreaks of campylobacteriosis.
Introduction
C
On the contrary, the newly identified species Campylobacter hepaticus currently causes significant economic losses to the poultry industry, as it is the cause of spotty liver disease (SLD) in chickens (Van et al., 2016, 2017a). The clinical manifestations of SLD include the formation of gray/white lesions in the liver, an increase in mortality rate in a flock, and reduction in egg production. SLD was first documented in the 1950s when a “vibrio” form of bacteria was suspected as the cause (Peckham, 1958). In more recent years, more specifically identified organisms, such as C. jejuni, C. coli, and Helicobacter pullorum, had been suspected to be the cause, but there was no definitive experimental verification of their potential role (Burnens et al., 1996; Grimes and Reece, 2011; Jennings et al., 2011). Recent studies in England and Australia resulted in the successful isolation and confirmation of C. hepaticus as the causative agent of SLD (Crawshaw et al., 2015; Van et al., 2016). A quantitative polymerase chain reaction (PCR) assay for quick and accurate detection of C. hepaticus in tissue and environmental samples has been developed and is suitable for epidemiology investigations of SLD (Van et al., 2017b). The pathogenicity of C. hepaticus has not been explored in other animals, and its pathogenic potential in humans is unknown. On the contrary, C. jejuni and C. coli are well-known human pathogens. Therefore, it is important to accurately identify C. hepaticus, C. jejuni, and C. coli to help control the spread of these Campylobacter pathogens.
The identification of Campylobacter to the species level, using isolation and subsequent biochemical and molecular testing, is laborious due to the slow growing nature of Campylobacter spp. (Mateo et al., 2005). Therefore, rapid DNA-based identification methods are of value. PCR to detect the hippuricase gene (hipO) has been used to detect C. jejuni as this gene is present in C. jejuni, but absent in most other Campylobacter species, and hippurate hydrolysis is the only biochemical test to differentiate C. jejuni and C. coli (Al Amri et al., 2007; Adzitey and Corry, 2011). However, the newly identified C. hepaticus species also carries this gene and most C. hepaticus strains were hippurate positive (Van et al., 2016). Therefore, PCR assays used to detect these species need to be evaluated and updated to prevent misidentification of these important Campylobacter species.
The study reported here aimed to develop a multiplex PCR assay for specific and simultaneous detection of C. jejuni, C. coli, and C. hepaticus. The assay was applied for detection of these pathogens in samples containing high-complexity microbiota from experimentally as well as naturally infected chickens.
Materials and Methods
DNA preparation
DNA from bacteria was isolated using the Isolate II genomic DNA kit (Bioline), according to the manufacturer's instructions. DNA was isolated from C. jejuni public culture collection strains and RMIT field isolates (C. jejuni NCTC 11168, C. jejuni NCTC 11828, C. jejuni 354, C. jejuni 54/1, C. jejuni 54/2; C. coli, C. coli NCTC 11366T, C. coli 134, C. coli 54/2) and 11 isolates from Australian SLD outbreaks from different farms in the states of Victoria, Queensland, South Australia, and New South Wales, including the type strain HV10 (C. hepaticus NCTC 13823T) (Van et al., 2017b). DNA from other Campylobacter species (C. concisus ATCC 51562, C. mucosalis ATCC 43264T, C. lari, C. upsaliensis, C. sputorum), H. pullorum, and Enterococcus cecorum (RMIT collection) were used to test the specificity of the primers. RMIT isolates were phenotypically and biochemically tested to confirm their identification.
DNA from cecal contents and cloacal swabs of SLD affected (birds with typical clinical indicators of SLD) and healthy birds were prepared using the Isolate Fecal DNA kit (Bioline), according to the manufacturer's instructions.
Primers and multiplex PCR conditions
For multiplex PCR, the primer pairs for each gene must produce different amplicon lengths that enable product differentiation in gel assays. Primers to detect the C. coli glyA gene were designed in this study and produced a 186 bp amplicon. Primers to detect the C. jejuni hipO gene and to produce a 323 bp amplicon were adapted from Wang et al. (2002), and primers for the unique C. hepaticus glycerol kinase gene (GK) were previously designed and produced a 463 bp amplicon (Van et al., 2017b) (Table 1).
PCR conditions were optimized, including varying ratios of primer pairs and optimization of annealing temperature. Multiplex PCR was carried out in a final volume of 20 μL using Q5 High-Fidelity 2 × Master Mix (New England Biolabs), primers at a final concentration of 250 nM for C. jejuni and C. hepaticus and 500 nM for C. coli and 1 μL of template DNA. An Eppendorf Mastercycler Pro PCR instrument was used for amplification with cycling conditions of 98°C for 1 min, 35 cycles of 98°C for 10 s; 59°C for 30 s, and 72°C for 30 s and final extension at 72°C for 10 min. Selected products were Sanger sequenced (Micromon, Monash University, Victoria, Australia) to confirm that the expected fragment had been amplified. PCR with universal primers (8F and 806R) targeting conserved bacterial 16S rRNA gene sequences was carried out as a positive control for PCR to demonstrate appropriate quality of all DNA templates (Turner et al., 1999; Caporaso et al., 2011).
PCR assay specificity, sensitivity, and application of the multiplex PCR assay for the detection of C. jejuni, C. coli, and C. hepaticus in field samples
The three primer sets were evaluated individually for primer specificity using a wide range of Campylobacter species and related organisms. DNA derived from a culture of known colony-forming units (CFUs) of C. hepaticus NCTC 13823T, C. jejuni ATCC 81116, and C. coli NCTC 11366T were diluted in 10-fold increments to determine the limit of detection of the PCR. In addition, relative detection ratios were also evaluated by varying the concentration of each species 100-fold. Thirty-six cecum samples and 12 cloacal swabs from chickens clinically affected with SLD (birds were orally challenged with C. hepaticus as described by Van et al., 2017a) were assayed for carriage of these Campylobacter species.
Bioinformatics to examine primer specificity
The target fragment of the glyA gene for C. coli detection was used as a nucleotide query sequence (186 bp) to compare against the nucleotide collection (nr/nt) and Campylobacter whole genome shotgun (WGS) contigs in the NCBI database using the Basic Local Alignment Search Tool (BLAST). When the identity was less than 100%, the alignment was examined to check the mismatch positions to see whether they are in the primer binding regions.
For primers used to detect C. hepaticus and C. jejuni in this study, the above procedure was not carried out as primers to detect these species were validated from previous studies (Wang et al., 2002; Van et al., 2017b). However, as C. hepaticus also encodes a hipO gene, the target fragment of the C. jejuni hipO gene (323 bp) was compared by BLAST against the genome sequences of C. hepaticus NCTC 13823T ( = CIT 111092T) (GenBank accession no. LUKK01000000), and the genome sequences of 14 C. hepaticus isolates from our collection and mismatch positions were examined.
Results
The designed and optimized multiplex assay showed specificity as only the corresponding strains showed the expected PCR amplification products. The sensitivity limit was 13–23 CFU/reaction (Fig. 1; Lane 1–6). In addition, the assay could detect the presence of all three species, even when they were present at highly different levels in the mixture (Fig. 1; Lane 7–9).
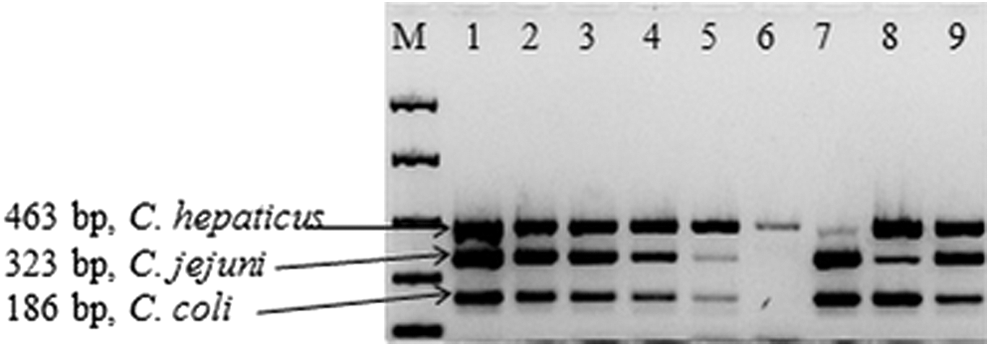
Sensitivity of the multiplex PCR protocol. M: Easy Ladder I, Bioline; Lane 1–6: PCR products from a serially diluted mixture of DNAs (equivalent to 1.3–2.3 × 105 to 1.3–2.3 CFU/reaction) from Campylobacter jejuni, Campylobacter coli, and Campylobacter hepaticus. Lane 5 shows the lowest concentration of DNA that the multiplex protocol was able to detect for these three species. Lane 7–9: Relative detection ratios test. C. hepaticus, C. jejuni, and C. coli DNA were added at 100-fold lower concentration than the DNA of the other two species, respectively. CFU, colony-forming unit; PCR, polymerase chain reaction.
When the 186 bp target fragment of the C. coli glyA gene was BLASTed against the NCBI nucleotide collection (nr/nt) and Campylobacter WGS databases, a total of 744 C. coli complete genomes and WGS entries in the NCBI database contained the full query sequence of 186 bp. Of these, 605, 110, and 29 genomes had sequences with 100%, 99%, and 96% identities to the query sequence, respectively. For the similarities of <100%, the alignment showed that the differences are positioned outside the primer binding sites, therefore, it would not affect the binding of primers to target sequence. The bioinformatics analysis demonstrates that all C. coli strains in the sequence database would be expected to encode a gene that would be amplified by the primers specified in Table 1. In addition, the BLAST results also showed that the primers designed to specifically target C. coli would not bind to the DNA of other Campylobacter species.
The alignment of the 323 bp C. jejuni hipO fragment against the genome sequences of C. hepaticus isolates showed 84% identities and a high level of dissimilarity in the primer binding regions, with only 11/20 base identities in the forward primer binding region and 13/20 base identities in the reverse primer binding region. All 14 C. hepaticus genomes contain identical 323 bp hipO fragments that are dissimilar to the C. jejuni sequence.
The assay was used to detect the presence of these species in field samples (Fig. 2). Out of 31 cecum samples from birds with clinical signs of SLD, 4 of them contained all 3 Campylobacter species; C. hepaticus, C. jejuni, and C. coli; 8 samples showed the presence of C. jejuni and C. hepaticus; 13 had C. coli and C. hepaticus; and 6 carried only C. hepaticus. Of five birds without clinical signs of SLD, none of them contained C. hepaticus; one contained both C. jejuni and C. coli; one contained only C. jejuni; two contained only C. coli; and no specific amplification was observed in one sample. Cloacal swabs from chickens (three control birds and nine SLD affected birds) were also assayed for carriage of these Campylobacter species. The multiplex assay showed that all nine diseased bird cecal swabs contained C. hepaticus and one also contained C. coli and none contained C. jejuni. In the swabs from the three control birds, a C. hepaticus-specific PCR amplicon was not detected in any of them, however, one bird contained C. jejuni and C. coli and one bird contained C. coli. C. hepaticus was isolated from the liver samples of all PCR-positive birds used in this study. The culturing of C. jejuni and C. coli was not undertaken, however, the PCR products were sequenced to confirm that the expected fragment had been amplified.
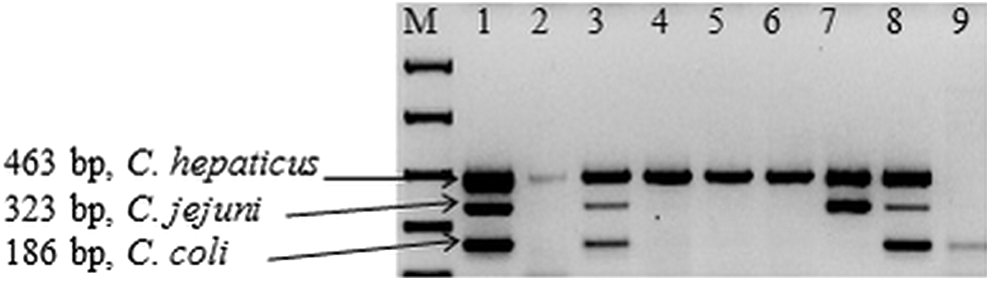
Multiplex PCR of selected field samples. Lane 1–8: multiplex PCR of SLD-infected birds; Lane 9: multiplex PCR of SLD-noninfected bird. PCR, polymerase chain reaction; SLD, spotty liver disease.
Discussion
Multiplex PCR assays have been developed and used for identification and differentiation of various Campylobacter species, such as C. jejuni, C. coli, C. lari, C. upsaliensis, and C. fetus subsp. fetus (Wang et al., 2002; Yamazaki-Matsune et al., 2007), but not yet for the newly identified C. hepaticus species. This new species is most closely related to the major pathogens C. jejuni and C. coli and has ∼144 fewer genes than C. jejuni (Petrovska et al., 2017). Primers designed to amplify the unique glycerol kinase gene from C. hepaticus have been shown to be specific for C. hepaticus and were therefore used in this study (Van et al., 2017b). The primers to detect C. jejuni, targeting the hipO gene, were designed by Wang et al. (2002) and these primers have been used successfully by different groups to examine the presence of C. jejuni in different sources (Wang et al., 2002; Kudirkiene et al., 2010; Madden et al., 2011; Adzitey et al., 2012). Until the discovery of C. hepaticus, the hipO gene was identified as unique to C. jejuni. The current study demonstrates that, although C. hepaticus does carry a hipO gene, the sequence is distinct from that of the C. jejuni gene, and hence, the primers designed to amplify the C. jejuni gene did not amplify the C. hepaticus gene. The analysis of 15 C. hepaticus genomes also supports the finding that there was no cross-reaction with the hipO primers.
Due to rapid development of high-throughput sequencing technologies a growing number of genomic sequences have been shared in public databases, making the search for DNA sequences for primer design that are conserved between isolates of the same species and nonconserved across other species of each genus, easier and more reliable. Presently, more than a thousand C. jejuni genome sequences and more than 800 C. coli genomes are available in the NCBI database. C. hepaticus has been discovered recently and there is currently only a single genome sequence available in the public database. The primers used to detect C. coli, designed in this study, were found to be specific in both the physical isolate screening and the in silico screening against the available sequences in the NCBI database.
The detection limit for the multiplex PCR developed in this study was close to the detection limit of a singlet PCR developed to detect C. hepaticus (13–23 CFU/reaction compared to 7.9 CFU/reaction) (Van et al., 2017b). The utility of the assay was also demonstrated by the ability to detect each of the three Campylobacter species when present at highly different levels in the mixture.
Use of the multiplex assay on clinical samples from birds indicated that there was a high incidence of C. jejuni and C. coli presence. Only 6 out of 31 samples did not contain either or both C. jejuni and C. coli. In addition, 4 out of 31 samples contained all 3 Campylobacter species.
It has been demonstrated that this multiplex assay can be used for both field samples and birds experimentally infected with C. hepaticus. Moreover, it can be used to detect the presence of these pathogens in cloacal swabs of chickens. Cloacal swabs represent a relatively noninvasive way to detect birds infected with pathogenic Campylobacter spp.
Conclusions
This multiplex PCR assay showed both high specificity and sensitivity for simultaneous detection of the human pathogens C. jejuni, C. coli, and the newly identified chicken pathogen, C. hepaticus. It represents a new practical diagnostic tool for rapid and sensitive investigation of the epidemiology of Campylobacter colonization in poultry, environmental samples, and food samples.
Footnotes
Disclosure Statement
No competing financial interests exist.