
Abstract
During 2014–2015, 300 pig carcasses before chilling and 85 food contact surfaces (FCSs) at cutting lines were tested for Salmonella in three slaughterhouses (namely A, B, and C) of northern Italy. In slaughterhouses A and B, four carcass sites of 100 cm2 each (from both the exterior and interior side) were swabbed with a single sponge. In abattoir C, four 100 cm2 sites of the exterior and the interior sides were swabbed with two independent sponges. The population average prevalence of Salmonella-positive carcasses (which takes into account the structure of the study design, with multiple samples collected in a single day) in slaughterhouses A and B was 12.3%, while in slaughterhouse C it was 11.2%. Presence of Salmonella on exterior and interior sides of carcasses showed a low level of concordance (only 3/12 of the contaminated carcasses were positive on both sides). No significant difference was found for FCSs contamination in the three slaughterhouses, with a population average prevalence of Salmonella-positive FCSs of 19.9%. In addition, we found that the clustering due to the day of sampling account for more than 36% and 60% of the overall prevalence variation on carcasses and FCSs, respectively. Eight serovars were identified, with Salmonella Derby as the most common type. The counting of Salmonella on carcasses showed large variability. It was low (<0.0075 most probable number [MPN]/cm2) in 46.6% of the carcasses and as high as 2.7 MPN/cm2 in 4.7%. Specifically, we found that counts on carcasses fit with “heavy tailed” distributions (lognormal and Weibull with a small shape parameter), suggesting not negligible probability of episodes of high Salmonella contamination. The mean values of contamination obtained with the two distributions ranged from 0.235 to 0.435 MPN/cm2.
Introduction
S
Pigs are asymptomatic carriers of several Salmonella serovars, particularly Salmonella Typhimurium, Salmonella Derby, Salmonella enterica 4,[5],12:i:- and Salmonella Rissen (Rostagno et al., 2007; van Hoek et al., 2012; Arguello et al., 2013; EFSA and ECDC, 2016). Subclinically infected pigs may shed Salmonella with their faeces and/or keep the pathogen in the gut, the associated lymph nodes, or the tonsils (Fedorka-Cray et al., 1999) and are considered the primary source of contamination of pig carcasses and the slaughterhouse environment (Berends et al., 1997; Baptista et al., 2010).
Estimating Salmonella burden on pig carcasses is required for consumer exposure assessment. For this purpose, the excision and swabbing techniques may be used and swabbing 400 cm2 is essentially equivalent to excising samples of 20 cm2 (Lindblad, 2007).
Pork meat and pork products have been frequently associated with human disease in the European Union (EU) (EFSA and ECDC, 2015), and in 2015, most cases were from Salmonella Derby (30.3%), the monophasic variants of Salmonella Typhimurium (30.3%) and Salmonella Typhimurium (27.1%) (EFSA and ECDC, 2016). Pork-related Salmonella outbreaks have also been detected in the study area of this work (Scaltriti et al., 2015; Morganti et al., 2018).
The aims of this study were to count Salmonella on pork carcasses in three pig slaughterhouses and to evaluate intraday contamination of food contact surfaces (FCSs) at cutting lines. Another study, conducted in a single slaughterhouse by Bonardi et al. (2016a), evaluated the prevalence of positive carcasses and FCSs at the cutting section with specific interest in the role of carcass dehairing in increasing Salmonella contamination. The present study extended that investigation to two further slaughterhouses, and the newly generated data set was analyzed considering alternative sampling approaches, estimating Salmonella counts on positive carcasses, and comparing the contamination on the outside and inside of carcasses. The study investigated also the relationship between the contamination of carcasses and the slaughterhouse FCSs.
Materials and Methods
Sampling
From February 2014 to May 2015, 300 pig carcasses before chilling and 85 environmental samples at cutting sections were tested for Salmonella in three slaughterhouses of northern Italy (here identified as: A, B, and C). Detection and typing data from slaughterhouse B are reported by Bonardi et al. (2016a). The abattoirs processed ca 380 pigs per hour. Sampled pigs were minimum 9-month-old with an average live weight of 160 kg. Carcasses belonged to 121 batches of pigs and were randomly sampled during 35 visits. After evisceration, 1–10 carcasses per batch (average: 2.48) were selected, the number depending on the possibility of removing the carcasses from the slaughter line because of practical arrangements. After evisceration and splitting, carcasses were sectioned on dressing tables before chilling at temperatures ranging from 2°C to 4°C. For the study, the carcasses were swabbed after splitting, before cutting and chilling. Samples were distributed as follows: abattoir A—100 carcasses from 66 batches (1.51 pigs per batch, on avarage), during 19 visits; abattoir B—90 carcasses from 19 batches (4.73 pigs per batch, on average) during 7 visits; abattoir C—110 carcasses from 36 batches (3.05 pigs per batch, on avarage), during 9 visits (Table 1).
This column applies to slaughterhouse C only.
FCSs, food contact surfaces.
In abattoir A and B, four different carcass sites of 100 cm2 each (back, belly, and jowl externally and the diaphragmatic area internally) were swabbed using a sterile sponge moistened with buffered peptone water (BPW; 3M Sponge Stick; 3M Health Care, St. Paul, MN). In abattoir C, four 100-cm2 sites of both the exterior side (ham, back, belly, and jowl) and the interior side (diaphragmatic and esophagus areas, jowl, and back) of the carcasses were swabbed by using 2 sponges, for a total of 220 samples. The sponges used for the exterior and interior sides were laboratory analyzed as independent samples.
During 30 sampling visits, a total of 85 samples from FCSs (about 3 samples/visit) were collected in the three slaughterhouses (Table 1). Specifically, the environmental samples included swabs from dressing tables and meat conveyors. FCSs were tested after the selected carcasses. Areas of 200–400 cm2 ca. were swabbed using a sterile sponge moistened with BPW (3M Health Care).
All samples were placed in sterile bags, transferred to the laboratory under refrigeration on the day of collection, and tested the same day.
Salmonella detection and typing
Samples from carcasses and FCSs were tested using the 3M™ Molecular Detection Assay 2—Salmonella (MDA Salmonella; 3M Health Care) based on the isothermal amplification and bioluminescence detection of specific sequences of Salmonella DNA. The MDA Salmonella method was validated by AFNOR in 2016. The sponges were suspended in 100 mL of BPW (3M Health Care). The initial suspension was manually shaken for 2 min and left at room temperature for 2 h for resuscitation. After removing 33.3 mL to be used for the MPN counting, as shown in the next paragraph, 66.7 mL of the BPW suspension was preenriched at 37°C ± 1°C for 16 to 20 h. Thereafter, 20-μL aliquots of the broth cultures were tested following the manufacturer's instructions. The MDA Salmonella assay reported the positive results in 20–75 min and the negative results after the run was completed (75 min). Positive results were considered “presumptive positive” to be confirmed by the ISO 6579:2002 method (ISO, 2002). Biochemical identification to the genus was performed by using the API® 20 E microsubstrate system (bioMérieux, Marcy l'Etoile, France).
Salmonella serotyping was performed according to the White-Kauffmann-Le Minor scheme by slide agglutination with O and H antigen-specific sera (DID, Milan, Italy; Biogenetics, Padua, Italy). Definitive identification of S. enterica 4,[5],12:i:–and its differentiation from Salmonella Typhimurium were genotypically obtained with the polymerase chain reaction protocol described by Barco et al. (2011).
Salmonella pulsed-field gel electrophoresis typing
Pulsed-field gel electrophoresis (PFGE) typing of Salmonella isolates was performed according to standard methods (PulseNet, 2010) with XbaI restriction of DNA.
Salmonella MPN counting
The counting of Salmonella was performed following the ISO/TS 6579-2:2012 (ISO, 2012) miniaturized most probable number (MPN) technique, except for the miniaturization of the dilutions. Samples giving a positive result by MDA Salmonella assay were tested by the MPN method. For each sponge, aliquots of 10, 1, and 0.1 mL of the BPW suspension were tested in triplicate. Considering that 100 mL of BPW was added to a sponge used to swab 400 cm2, aliquots of 10 mL corresponded to 40 cm2, 1 mL to 4 cm2, and 0.1 mL to 0.4 cm2. After incubation at 37°C ± 1°C for 16–20 h, 20 μL from each was seeded in a single well (9 wells in total) of a 12-well microtiter plate containing 2 mL per well of Modified semisolid Rappaport Vassiliadis (MSRV; Oxoid). After incubation at 41.5°C ± 1°C for 24 h ± 3 h, the MSRV wells showing a gray–white zone extending out of the inoculum drop were further tested by streaking a 10-μL loopful of the bacterial growth onto XLD (Oxoid) agar plates. Microtiter plates not showing bacterial growth were incubated for further 24 h ± 3 h before being considered negative.
XLD agar plates were incubated at 37°C ± 1°C for 24 h and suspect Salmonella colonies were confirmed following the ISO 6579 method. Positive tubes were converted into MPN counts through the three-replicate MPN table (Blodget, 2010). To estimate the MPN per cm2, the result had to be divided by 400.
Statistical analysis
We assessed whether Salmonella prevalence on pig carcasses and slaughterhouse environments was different between the slaughterhouses by fitting generalized linear mixed models (GLMM) with binomial error distribution with the occurrence of Salmonella as response variable, the slaughterhouse of origin as fixed effect, and the day of sampling as random effect. We introduced the random effect since observations were not independent, but rather clustered within days, with a potentially significant variability in prevalence in different sampling days, as suggested by Corbellini et al. (2016). The significance of the slaughterhouse effect on prevalence was assessed through log-likelihood ratio tests (LRTs) against the null models. To take into account the effect of the sampling day, the estimated Salmonella prevalence from the GLMM was obtained by converting the subject-specific estimates derived from the model to a population average estimate based on the total variance of the random effect (Zeger et al., 1988). We computed the intraclass correlation coefficients (ICC) from GLMM to estimate the effect of clustering within sampling days.
The degree of agreement in Salmonella detection between exterior and interior carcass samples collected in slaughterhouse C was performed through the concordance coefficient, k-Cohen (Cohen, 1960).
To test the differences in the empirical distributions of Salmonella counts, we used two-sample logrank tests with hypergeometric variance based on left-censored data (Prentice and Marek, 1979). We fitted parametric univariate distributions to Salmonella MPN counting on positive carcasses through fitting techniques for semiquantitative data, to take into account nonquantifiable samples. According to Pouillot and Delignette-Muller (2010), we fitted the semiquantitative data by using left-bounded continuous distribution families (lognormal, exponential, and Weibull distributions). We estimated the distribution parameters by using maximum likelihood techniques and the uncertainties in the parameters through nonparametric bootstrap resampling (with 1000 iterations). The best model family describing count distribution, was obtained through a model selection procedure based on Akaike Information Criterion (AIC) (Akaike, 1974). Models were ranked using their differences in AIC (ΔAIC). The best models were selected using the threshold ΔAIC ≤4 (Burnham and Anderson, 2002). Statistical analyses were performed by using the packages “lme4,” “Envstats,” and “fitdistrplus” in the R 3.2.0 environment (R Development Core Team, 2015).
Results
Salmonella contamination on pig carcasses
Tested and Salmonella-positive carcasses and FCSs are reported in Table 1. Prevalence values were calculated separately for abattoirs A and B (one sponge/carcass) and abattoir C (two sponges/carcass). Salmonella was detected in 27/190 carcasses tested in A and B. The analysis of the GLMM showed that no significant difference in prevalence was observed between A and B, where a similar sensitivity is expected as a consequence of the same sampling method (LRT, df = 1, p = 0.383). The population average prevalence of Salmonella-positive carcasses in A and B, computed through the GLMM, was estimated as 0.123 (95% confidence interval [CI]: 0.051–0.205). The ICC within sampling days in A and B was ICC = 0.383. In C, 12/110 carcasses were positive. The population average prevalence on carcasses sampled in C (both inside and outside), computed through GLMM, was 0.112 (95% CI: 0.022–0.233). The ICC within sampling days in C was ICC = 0.362. Considering exterior and interior side samplings, we detected four carcasses contaminated on the exterior side only, five contaminated on the interior side only, and three contaminated on both sides. The analysis of agreement in Salmonella detection between exterior and interior samples showed a concordance k-Cohen = 0.356 (95% CI = 0.028–0.684), which is interpreted as an only “fair” level of concordance according to Altman (1991).
Forty-two Salmonella isolates were detected on pig carcasses. They belonged to five serovars: Salmonella Derby (24/42; 57.1%), Salmonella enterica 4,[5],12:i:- (9/42; 21.4%), Salmonella Rissen (6/42; 14.3%), Salmonella Bovismorbificans (2/42; 4.8%), and Salmonella Brandenburg (1/42; 2.4%).
Nine XbaI PFGE profiles were identified among Salmonella Derby (D1–D9), three among Salmonella enterica 4,[5],12:i:- (MT1–MT3) and three among Salmonella Rissen (R1–R3) (Fig. 1). Isolates belonging to the same pulsotype (D2 and D4) were detected in different carcasses during 8/35 visits. In three cases, the carcasses belonged to the same batch, while in five cases, they belonged to different batches. Pulsotypes D2 and D4 were also found in different slaughterhouses. This is consistent with the fact that genotype D2 of Salmonella Derby was the most common (11/24 of the isolates) (Fig. 1). In slaughterhouse C, the same pulsotypes were detected on both sides of positive carcasses (Table 2).
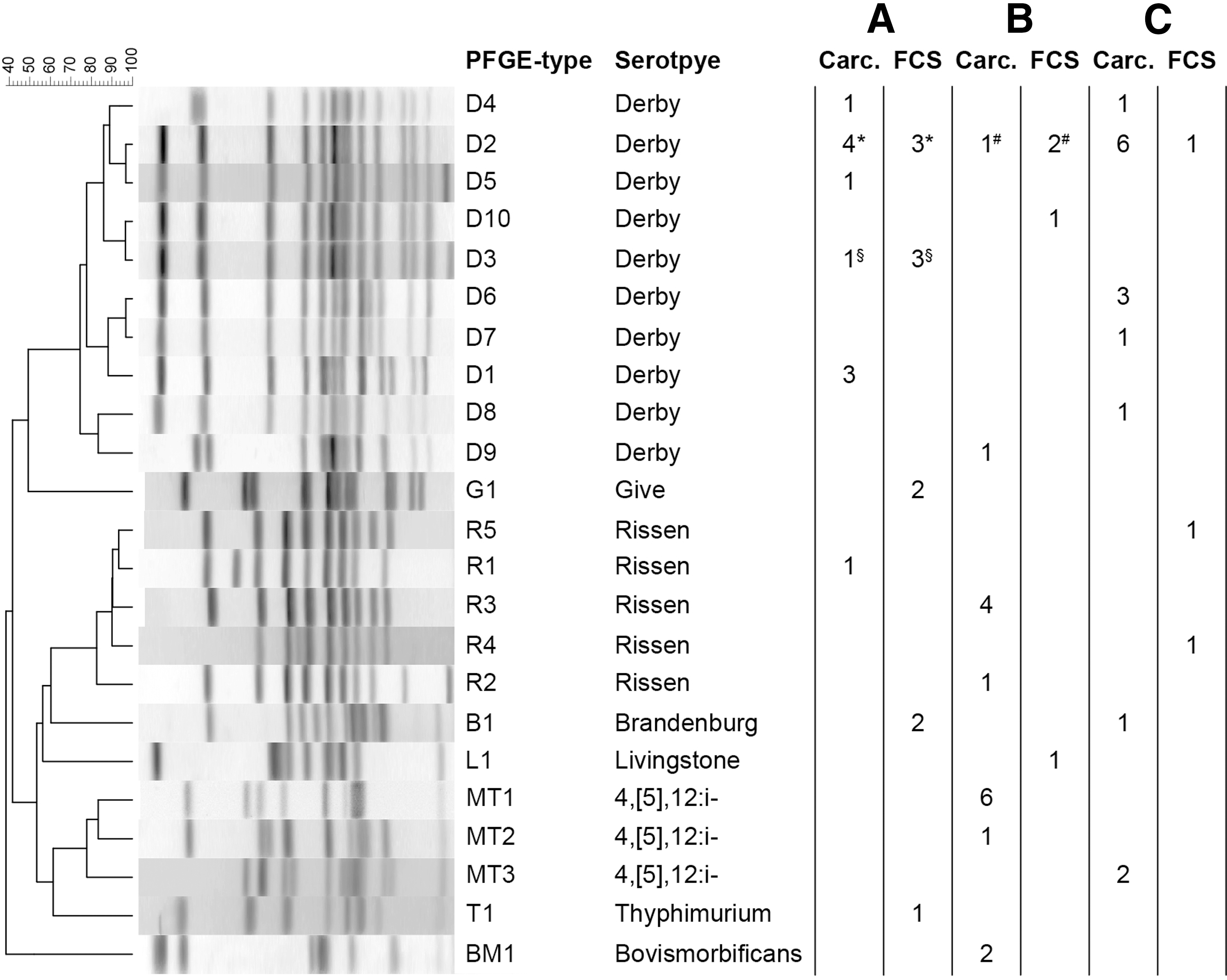
Distribution of serovars and PFGE types of Salmonella enterica detected on pig carcasses. The PFGE types are identified with a different code. For each serovar, the number of isolates belonging to the different genotypes is shown in brackets. A, B, C letters identify the slaugherhouses. Salmonella Derby corresponds to genotypes D1–D9; Salmonella enterica 4,[5],12:i:- to MT1–MT3; Salmonella Rissen to R1–R3; Salmonella Bovismorbificans to BM1; and Salmonella Brandenburg to B1. *,#,§Indicate carcass-FCS intraday cross-contamination. FCS, food contact surface; PFGE, pulsed-field gel electrophoresis.
Pulsotypes are identified with their identification code.
Salmonella counting on pig carcasses
Data are shown in Tables 2 and 3. Quantifiable counts were obtained in 17/27 positive carcasses in A and B (63.0%) and in 11/15 positive carcasses in interior and exterior samples in C (73.3%). MPN counts did not differ significantly between A and B (test statistic ν = −1.626; p = 0.407). MPN counts did not differ significantly between exterior and interior sides in C (test statistic ν = 0.303; p = 0.851). However, significantly higher MPN counts were found on carcasses positive on both sides than in single-side positive carcasses (test statistic ν = 3.91; p = 0.001).
Since A and B did not display significant difference in the empirical distribution of MPN counts, we fitted parametric univariate distributions using counts data of both slaughterhouses. We found that the lognormal (AIC = 12.64) and the Weibull (AIC = 13.6) distributions provided similar fits for data of A and B (ΔAIC <1), while the exponential distribution (AIC = 82.96) was less fitting (Table 4).
Values in the brackets represents the 95% CI computed through nonparametric bootstrap resampling (1000 iterations). Models were ranked using the AIC. The best models were selected using a threshold of ΔAIC ≤4.
ΔAIC, differences in AIC; AIC, Akaike Information Criterion; CI, confidence interval.
The estimated distributions of counts from A and B, obtained with the best models, are displayed in Figure 2. The median values of Salmonella were estimated at 0.0157 MPN/cm2 (95% CI: 0.0048–0.046) in the lognormal and 0.0161 MPN/cm2 (95% CI: 0.0038–0.056) in the Weibull models. The 95th percentile values were estimated at 1.168 MPN/cm2 (95% CI: 0.204–5.075) in the lognormal and MPN/cm2 1.158 (95% CI: 0.219–3.52) in the Weibull models. The mean values of Salmonella counts were estimated at 0.432 MPN/cm2 (95% CI: 0.053–6.4) in the lognormal and 0.235 MPN/cm2 (95% CI: 0.05–0.902) in the Weibull models.
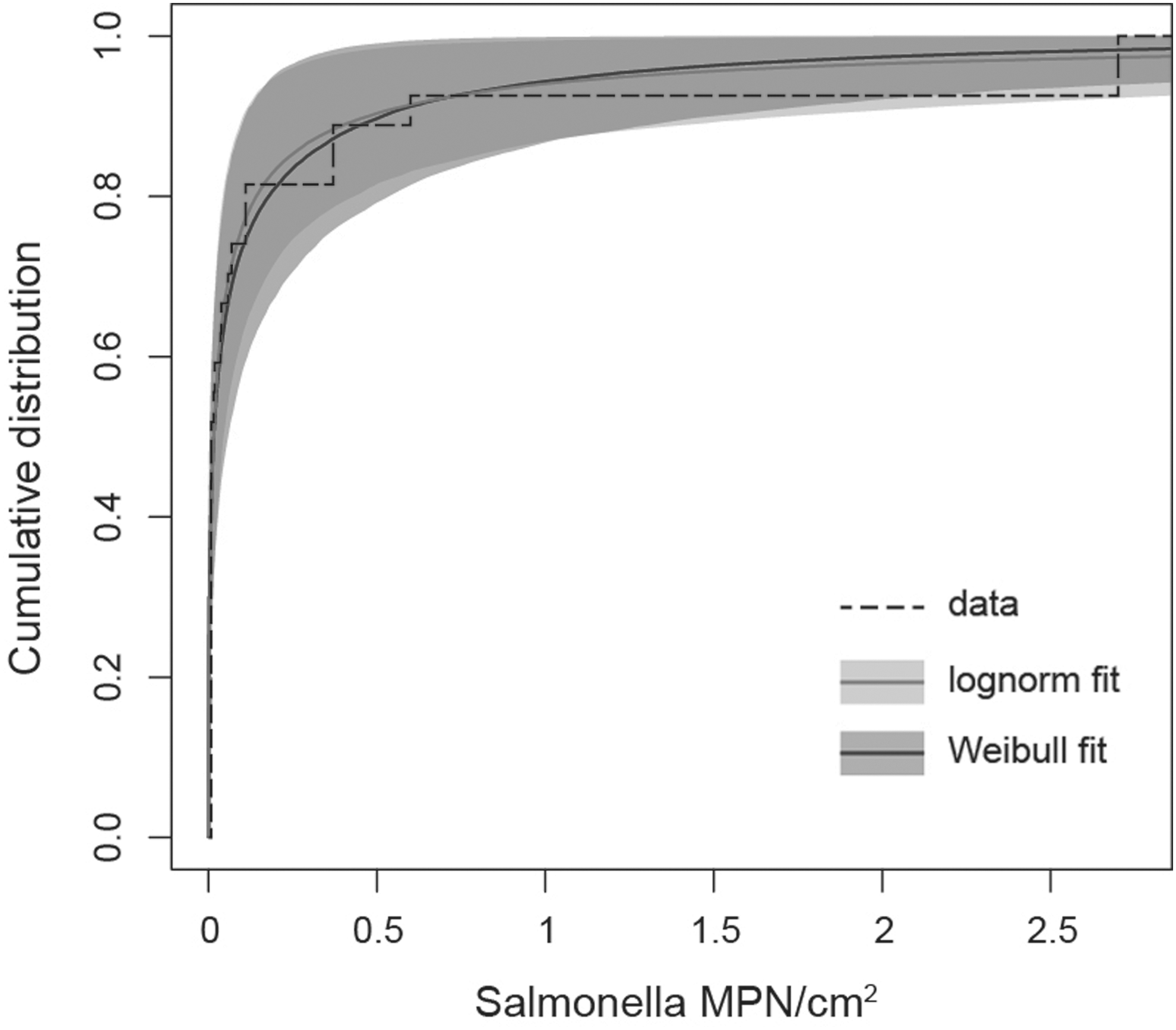
Cumulative distribution plot of Salmonella MPN counts, assuming lognormal distribution (in gray) and Weibull distribution (in dark gray). Shaded regions correspond to the 95% uncertainty range on each quantile of variability. Scale parameters: −4.179 (range: −5.341 to −3.146) lognormal; 0.0462 (range: 0.0129–0.147) Weibull. Shape parameters: 2.584 (range: 1.72–3.534) lognormal; 0.349 (range: 0.254–0.538) Weibull. The dashed line represents the observed data.
Salmonella contamination in the slaughterhouse environment
Salmonella was detected in 18/85 FCS samples (Table 1). No significant differences in prevalence were found between the slaughterhouses (LRT, df = 2, p = 0.751). The population average prevalence of Salmonella-positive samples in FCSs, computed through GLMM following Zeger et al. (1988), was estimated as 0.199 (95% CI: 0.059–0.334). The ICC within sampling days in FCSs was ICC = 0.607.
Six serovars were identified. Salmonella Derby was the most common (10/18; 55.6%), followed by Salmonella Brandenburg (2/18), Salmonella Give (2/18), Salmonella Rissen (2/18), Salmonella Livingstone (1/18), and Salmonella Typhimurium (1/18). XbaI PFGE identified three different pulsotypes for Salmonella Derby (D2, D3, D10), two for Salmonella Rissen (R4, R5), and one pulsotype each for Salmonella Brandenburg (B1) and Salmonella Give (G1). In different sampling sessions, the isolates detected on meat conveyors and dressing tables belonged to pulsotypes detected in the carcasses processed on the same day. However, also genotypes of Salmonella Derby and Salmonella Rissen (D10, R4, and R5) and serovars not detected on carcasses (Salmonella Typhimurium, Salmonella Give, and Salmonella Livingstone) were identified on FCSs (Fig. 1).
Discussion and Conclusions
In this study, the overall average prevalence of Salmonella-contaminated carcasses was 12.3% in the slaughterhouses sampled with one swab per carcass (A and B) and 11.2% in the slaughterhouse sampled with two swabs per carcass (C). However, our analyses evidenced that the contamination of carcasses was critically dependent on the day of sampling. Specifically, we found that a large fraction of the overall variation in the prevalence (more than 0.36 and 0.6 for carcasses and FCSs, respectively) is explained simply by intraday clustering, indicating a higher probability to find multiple positives within the same day. This is in accordance with Corbellini et al. (2016), who observed that the effect of the day of sampling on prevalence of Salmonella-positive carcasses was very large, specifically when pig batches with high prevalence of carriers shedding high numbers of Salmonella were slaughtered.
Despite the small number of carcasses tested per day (5–20) compared with the large number of slaughtered pigs (ca 3000 pigs/day), in two slaughterhouses the same pulsotypes of Salmonella Derby on both carcasses and FCSs were detected on three same sampling days, suggesting that Salmonella cross-contamination from carcasses to cutting lines could have occurred. This finding highlights the role of slaughtered pigs as source of contamination of FCSs involved in postslaughter processing of carcasses and is consistent with Botteldoorn et al. (2003) who observed that cross-contamination at slaughter was influenced by the Salmonella status of the supplied animals (Botteldoorn et al., 2003).
The most commonly detected serovars were Salmonella Derby, Salmonella enterica 4,[5],12:i:- and Salmonella Rissen, known to cause human infections in different countries (Kerouanton et al., 2013; Frank et al., 2014; Arnedo-Pena et al., 2016; Bonardi et al., 2016b; EFSA and ECDC, 2016; García-Fierro et al., 2016). Our results are consistent with the EU baseline survey (EFSA, 2008), except for Salmonella Typhimurium that was never found on carcasses and was detected with low prevalence on cutting lines.
The mean count values ranged from 0.235 to 0.432 MPN/cm2 on carcasses before chilling. These values are comparable to the mean value of 0.48 MPN/cm2 estimated by van Hoek et al. (2012) for carcasses tested by the excision of 5 cm2 areas in four sampling sites. In our study, counts were highly variable. They were low (<0.01 MPN/cm2) in 46.6% of the samples, while they reached 2.7 MPN/cm2 in 4.7% of the carcasses, a high value compared to the estimated mean of the study. This behaviour reflects the “heavy-tailed” distribution of Salmonella counts, as shown by the better fitting of Weibull (with shape parameter smaller than 1) and lognormal distributions than exponential distributions (Fig. 2). This “heavy-tailed” distribution of counts entails a non-negligible probability of observing Salmonella counts several times higher than the mean value. With regard to food-safety, this indicates a not negligible risk of occasional downstream food-chain contaminations and potential consumer infections.
Footnotes
Acknowledgments
The authors gratefully acknowledge Dr. Gisella Pizzin, Mrs. Ida Poli, and Dr. Ilaria Bruini of the University of Parma (Department of Veterinary Science) for technical assistance. E.S. was supported by the Italian Ministry of Health grant PRC 011/2013.
Disclosure Statement
No competing financial interests exist.