
Abstract
Salmonella enterica serovar I 4,[5],12:i:- has emerged as a common nontyphoidal Salmonella serovar to cause human foodborne illness. An interesting trait of serovar I 4,[5],12:i:- is that it only expresses the fliC gene for bacterial motility (i.e., monophasic), while most Salmonella strains alternately express two flagellin genes (fliC and fljB). The goal of this study was to characterize the porcine response following inoculation with a multidrug-resistant (MDR) serovar I 4,[5],12:i:- isolate associated with a multistate pork outbreak to determine if the increased prevalence of serovar I 4,[5],12:i:- in swine is due to enhanced pathogenicity. Pigs were inoculated and subsequently evaluated for the ability of the isolate to colonize intestinal tissues, cause clinical symptoms, induce an immune response, and alter the fecal microbiota over a 7-day period. Pigs exhibited a significant increase in rectal temperature (fever) (p < 0.01) and fecal moisture content (diarrhea) (p < 0.05) at 2 days postinoculation (d.p.i.) compared with preinoculation (day 0). Serum analyses revealed significantly increased interferon-gamma (IFN-γ) levels at 2 (p ≤ 0.0001) and 3 (p < 0.01) d.p.i. compared with day 0, and antibodies against Salmonella lipopolysaccharide (LPS) were present in all pigs by 7 d.p.i. Serovar I 4,[5],12:i:- colonized porcine intestinal tissues and was shed in the feces throughout the 7-day study. Analysis of the 16S rRNA gene sequences demonstrated that the fecal microbiota was significantly altered following MDR serovar I 4,[5],12:i:- inoculation, with the largest shift observed between 0 and 7 d.p.i. Our data indicate that the pork outbreak-associated MDR serovar I 4,[5],12:i:- isolate induced transient clinical disease in swine and perturbed the gastrointestinal microbial community. The porcine response to MDR serovar I 4,[5],12:i:- is similar to previous studies with virulent biphasic Salmonella enterica serovar Typhimurium, suggesting that the absence of fljB does not substantially alter acute colonization or pathogenesis in pigs.
Introduction
S
Numerous animal species are reservoirs for Salmonella, including reptiles, cattle, poultry, and pigs (Stevens et al., 2009). In swine, infection with Salmonella enterica serovar Choleraesuis can cause systemic disease, but other serovars, such as Salmonella enterica serovar Typhimurium, are opportunistic pathogens that often lead to asymptomatic colonization resulting in unrecognized transmission and contamination (Stevens et al., 2009). Reports have identified the rapid emergence and global dissemination of a monophasic variant, Salmonella enterica serovar I 4,[5],12:i:- (Arguello et al., 2014; Barco et al., 2014; Yang et al., 2015; Elnekave et al., 2017). This monophasic variant expresses the first-phase flagellar antigen encoded by the fliC gene, but not the second-phase flagellar antigen, FljB (Echeita et al., 2001). Pork products are a primary source of serovar I 4,[5],12:i:- contamination (Hauser et al., 2010). In the United States, an outbreak of serovar I 4,[5],12:i:- linked to pork products occurred in 2015, whereby 188 individuals were infected resulting in 30 hospitalizations. Investigation of 10 clinical outbreak isolates revealed that all strains were multidrug-resistant (MDR; resistant to ≥3 antimicrobial classes), including resistance to ampicillin, streptomycin, sulfisoxazole, and tetracycline (R-type ASSuT) (Kawakami et al., 2016). This is a public health concern, as human infection with antimicrobial-resistant Salmonella strains are associated with increased bacteremia, hospitalization, and treatment failure (Varma et al., 2005).
As pigs have been identified as a primary reservoir of serovar I 4,[5],12:i:- strains, we evaluated the ability of an MDR serovar I 4,[5],12:i:- outbreak isolate to colonize and cause clinical symptoms in swine, induce an immune response, and modify the porcine fecal microbiota to determine if the increased prevalence in swine is due to enhanced acute colonization or pathogenicity. This information could lead to future vaccine design, or other preharvest control measures, to limit and/or eliminate colonization in the host species. Overall, our data indicate MDR serovar I 4,[5],12:i:- induced transient clinical disease, stimulated the production of circulating interferon-gamma (IFN-γ) and Salmonella lipopolysaccharide (LPS)-specific antibodies, and altered the microbial community of the porcine gastrointestinal tract.
Materials and Methods
MDR serovar I 4,[5],12:i:- isolate description
In 2015, a multistate outbreak of MDR serovar I 4,[5],12:i:- was associated with pork products from Washington State (Kawakami et al., 2016); Salmonella isolates have the R-type ASSuT (Kawakami et al., 2016). The USDA Food Safety and Inspection Service (FSIS) isolated 11 serovar I 4,[5],12:i:- strains with the primary PFGE pattern JPXX01.1314 from cecal contents of postslaughter pigs during the outbreak investigation. An isolate, FSIS 1503788, was passaged in swine, isolated from the ileocecal lymph node (ICLN), and designated strain SX 240.
Swine study
Eight Yorkshire crossbred pigs from five Salmonella-fecal-negative sows were weaned at 16 days of age, shipped to the National Animal Disease Center, Ames, IA, and housed in a single isolation room. Pigs tested fecal-negative for Salmonella twice over a 6-week period, as described previously (Bearson et al., 2010). At 8 weeks of age, all pigs were inoculated via the intranasal route with 1 × 109 colony-forming units (CFU) of strain SX 240. Sampling intervals included 0, 1, 2, 3, and 7 days postinoculation (d.p.i.). To evaluate the ability of SX 240 to cause disease in pigs, clinical parameters of infection (fever, diarrhea) were monitored during the 7-day trial via body temperature measurements and fecal moisture content assessments. Body temperatures were monitored using a rectal thermometer. Fecal samples were collected for microbiota analysis, moisture content evaluation, and quantitative and qualitative bacteriology analyses. Blood samples were collected from the jugular vein for serum antibody and IFN-γ ELISA, as previously described (Bearson et al., 2017). Serum samples collected on 0 and 7 d.p.i. were used to determine antibodies to LPS antigen using the IDEXX HerdChek® Swine Salmonella Test Kit (IDEXX Europe B.V., Hoofddorp, Netherlands), following the manufacturer's instructions. Serum samples collected at 0, 1, 2, 3, and 7 d.p.i. were used to determine IFN-γ levels using the Porcine IFN gamma ELISA Kit (Pierce Biotechnology, Rockford, IL), following the manufacturer's instructions. One pig died during the study due to complications unrelated to Salmonella inoculation. At 7 d.p.i., all pigs were euthanized and necropsied to obtain tissue samples of tonsil, cecum, ileocecal lymph nodes (ICLN), and Peyer's patch region of the ileum. Contents of the cecum (1 g) and mucosal scrapings at the ileal-cecal junction (0.01 g in 0.1 mL PBS) were also evaluated by quantitative and qualitative bacteriology analyses.
Bacteriology
Quantitative and qualitative Salmonella culture analyses of fecal and tissue samples were performed as previously described (Bearson et al., 2016), utilizing XLT-4 medium (Becton, Dickinson and Co., Sparks, MD) supplemented with 50% tergitol, ampicillin (100 μg/mL), tetracycline (15 μg/mL), novobiocin (50 μg/mL), and streptomycin (50 μg/mL). Suspected Salmonella colonies were evaluated on BBL™ CHROMagar™ Salmonella (Becton, Dickinson and Co.) for mauve colonies indicative of Salmonella.
Fecal moisture content
To determine fecal moisture content, 1 g of feces was placed in an aluminum pan, weighed, incubated in a drying oven at 70°C for 22 h, and weighed again. Percent moisture was calculated by subtracting the dry weight from the wet weight, dividing by the wet weight, and multiplying by 100.
16S rRNA gene sequencing and analysis
Fecal samples were immediately placed on ice following collection and stored at −80°C until processing. Total DNA was extracted using the PowerMag Microbiome RNA/DNA Isolation Kit (MoBio, Carlsbad, CA) according to the manufacturer's instructions and adapted for use on a Biomek FXP laboratory automation workstation (Beckman Coulter, Indianapolis, IN). Four wells containing water instead of sample were included to test extraction quality.
The 16S rRNA gene libraries were created as previously described (Allen et al., 2016) with the exception that the PCR primers 515-F 5′-GTGCCAGCMGCCGCGGTAA-3′ and 806-R 5′-GGACTACHVGGGTWTCTAAT-3′ were used to amplify the V4 hypervariable region of the 16S rRNA gene using previously published barcodes (Kozich et al., 2013). In addition, a mock community consisting of 1 archaeal and 19 bacterial species (Allen et al., 2016) was amplified in one well of each plate for quality control purposes. Libraries were sequenced on an Illumina MiSeq (Illumina, San Diego, CA) using the MiSeq Reagent Kit v2 (2 × 250).
16S rRNA gene sequences were processed using the QIIME software package v. 1.9.1 (Caporaso et al., 2010b). Paired-end 16S rRNA gene reads were joined using fastq-join (Aronesty, 2013) with a minimum overlap of 200 bp and a maximum percent difference of 3. Joined reads were quality-filtered with reads being truncated following two consecutive base calls of a quality score of less than 25. Sequences with ≥85% of the original sequence length following truncation were retained. Chimeric sequences were removed using the UCHIME algorithm (Edgar et al., 2011) implemented in VSEARCH V. 2.4.0 (Rognes et al., 2016). Remaining sequences were clustered into operational taxonomic units (OTUs) at 97% similarity using a de novo OTU picking method implemented in VSEARCH. Taxonomy was assigned to OTUs using the UCLUST consensus taxonomy assigner (Edgar, 2010) and SILVA SSU database v. 128 (Quast et al., 2013) with a minimum similarity of 0.8 and max accepts of 3. Representative sequences for each OTU were aligned using PyNAST (Caporaso et al., 2010a), and a phylogenetic tree was constructed using FastTree (Price et al., 2010). Based on the results of the mock community OTU clustering, OTUs containing fewer than 10 sequences were excluded from further analysis, as were those classified as chloroplasts and mitochondria. Raw sequences were submitted to the NCBI Sequence Read Archive under BioProject PRJNA382998.
Salmonella invasion assays
IPEC-J2 cells, which were derived from the small intestine of piglets (Rhoads et al., 1994), were grown and maintained as previously described (Bearson et al., 2017). Before use, IPEC-J2 cells were confirmed free of Mycoplasma contamination using the Lookout® Mycoplasma PCR Detection Kit (Sigma-Aldrich, St. Louis, MO). The ability of the MDR serovar I 4,[5],12:i:- strain SX 240 to invade IPEC-J2 cells was compared to virulent, non-MDR, biphasic Salmonella Typhimurium strain χ4232 (SX 117) (Bearson et al., 2014) as previously described (Bearson et al., 2017), with the following modifications: multiplicity of infection of 10 and gentamicin concentration of 100 μg/mL. Experiments were performed in triplicate (each with technical replication), and percent invasion was calculated by dividing CFU/mL recovered by CFU/mL added and multiplying by 100.
Statistical analyses
Statistical analyses of body temperature, fecal moisture content, and serum IFN-γ levels were performed using repeated measures analysis of variance (ANOVA) with a Dunnett's multiple comparison test with comparison to the day 0 control. Analysis of invasion assay data was performed using an unpaired t-test. p-values less than 0.05 were considered significant. Statistical analyses were performed using GraphPad Prism 7.0 (GraphPad Software, Inc., La Jolla, CA).
Alterations in the archaeal and bacterial community structure over the 7-day sampling period were assessed using weighted UniFrac distances (Lozupone et al., 2007). All samples were randomly subsampled to 9000 sequences before analysis to account for unevenness in sequencing depth. Weighted UniFrac distances were calculated and plotted using the R v. 3.3.3 packages phyloseq v. 1.19.1 (McMurdie and Holmes, 2013) and vegan v.2.4.2 (Oksanen et al., 2017). Permutational multivariate ANOVA (PERMANOVA) using the adonis function, with 10,000 permutations in vegan, was used to analyze the weighted UniFrac distances by sampling time. The homogeneity of dispersion for each sampling time was determined using the betadisper function of vegan. Linear discriminant analysis effect size (LEfSe) (Segata et al., 2011) was used to identify genera with a relative abundance of greater than 0.1% that were more abundant at 0 d.p.i. compared with 1, 2, 3, and 7 d.p.i. A minimum linear discriminant analysis (LDA) score of 4.0 was used as the threshold for classifying differentially abundant genera.
Results and Discussion
Salmonella uses host-specific factors to colonize the gastrointestinal tract; therefore, it is important to study host–pathogen interactions in an animal that represents a risk to food safety (Bearson and Bearson, 2011). In this study, the natural porcine host was used to characterize an MDR serovar I 4,[5],12:i:- strain isolated during the investigation of a foodborne outbreak associated with pork products (Kawakami et al., 2016). This study may provide insight on the mechanism by which serovar I 4,[5],12:i:- has increased prevalence in swine and human foodborne illness.
MDR serovar I 4,[5],12:i:- causes transient clinical disease in swine
To evaluate the ability of SX 240 to cause disease in pigs, clinical parameters of infection (fever, diarrhea) were monitored during the 7-day trial. The average body temperature significantly increased at 2 d.p.i. (40.3°C) compared with day 0 (39.4°C) (p < 0.01; Fig. 1). Also at 2 d.p.i., percent fecal moisture content (diarrhea) increased significantly (79.3%) compared with day 0 (74.9%) (p < 0.05; Fig. 2). These results indicate that, like virulent biphasic Salmonella Typhimurium, MDR serovar I 4,[5],12:i:- can induce transient (i.e., mild and self-limiting) clinical disease in swine (Patterson et al., 2016).
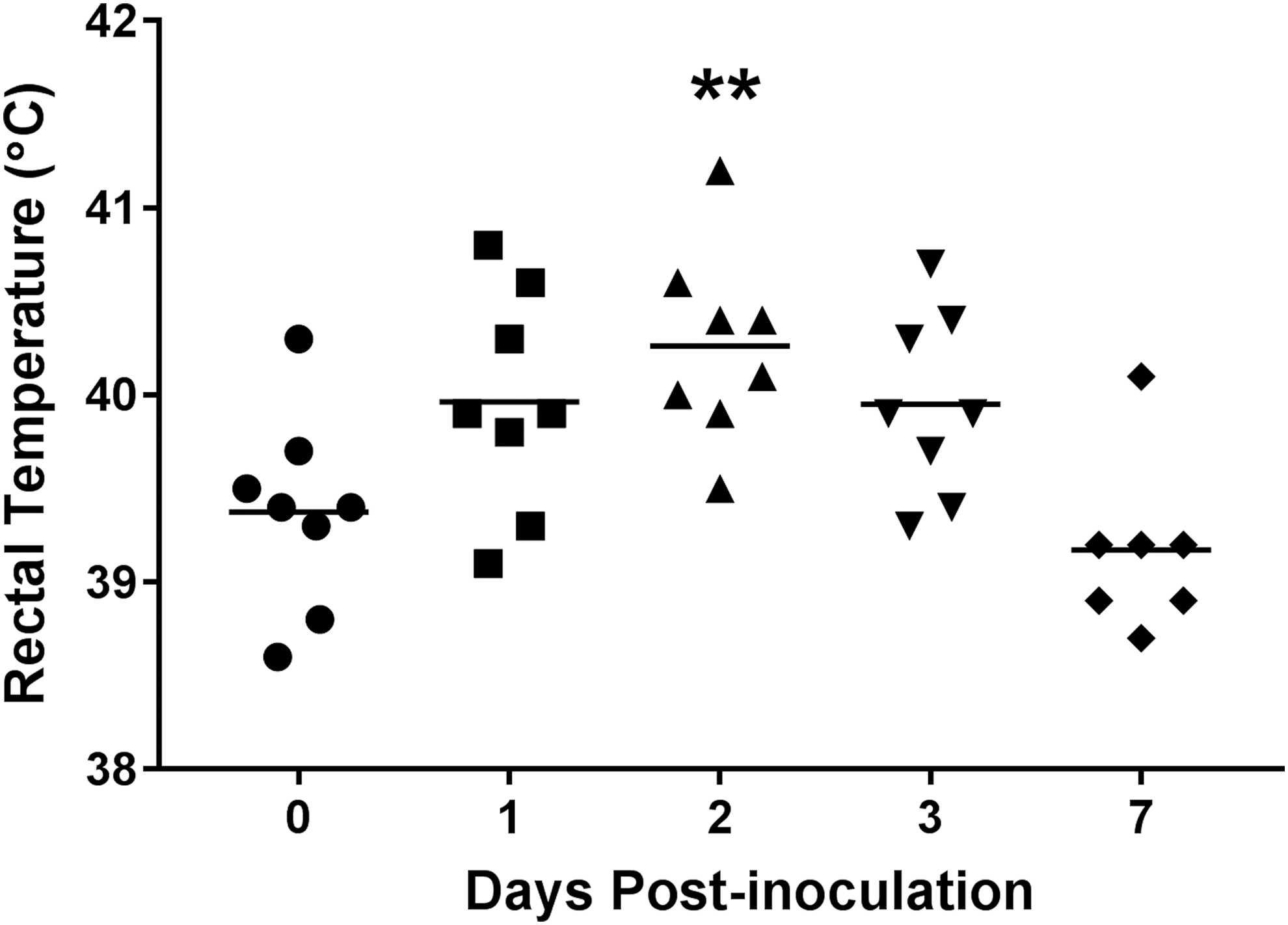
Body temperature is significantly increased in pigs inoculated with MDR Salmonella enterica serovar I 4,[5],12:i:-. At 8 weeks of age, pigs were inoculated with MDR serovar I 4,[5],12:i:-, and rectal temperatures (fever) were recorded at 0, 1, 2, 3, and 7 d.p.i. Each data point represents the temperature of an individual pig with the horizontal line representing the mean for each time point. **Significant difference (p < 0.01) when compared to day 0. d.p.i., days postinoculation; MDR, multidrug-resistant.
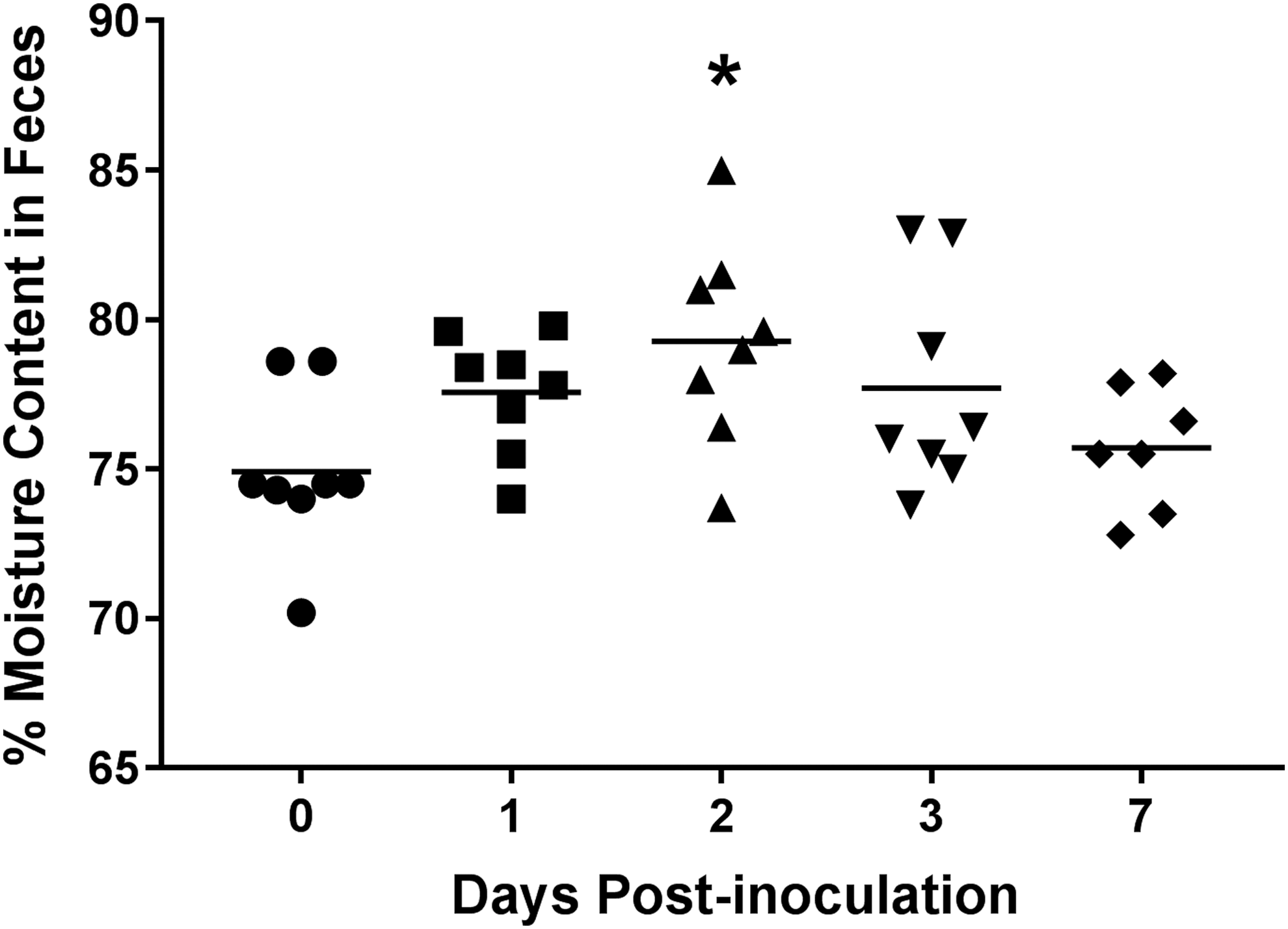
Significantly increased moisture content in swine feces following inoculation with MDR Salmonella enterica serovar I 4,[5],12:i:-. Moisture content (diarrhea) of pig feces collected at 0, 1, 2, 3, and 7 d.p.i. was determined. Each data point represents the fecal moisture content of an individual pig with the horizontal line representing the mean for each time point. *Significant difference (p < 0.05) when compared to day 0.
Fecal shedding and tissue colonization of MDR serovar I 4,[5],12:i:- in swine
MDR serovar I 4,[5],12:i:- fecal shedding and tissue colonization levels were determined to evaluate colonization and transmission potential. Peak fecal shedding levels occurred at 1 d.p.i. (∼104 CFU/g feces) and decreased to ∼102 CFU/g feces by 7 d.p.i. (Fig. 3), indicating that SX 240 is shed at levels typically seen in pigs inoculated with virulent, non-MDR, biphasic Salmonella Typhimurium (Bearson et al., 2014). Transmission studies suggest that exposure of pigs to relatively low numbers of Salmonella (102–103 CFU) is sufficient for colonization (Hurd et al., 2001; Boughton et al., 2007a, b).
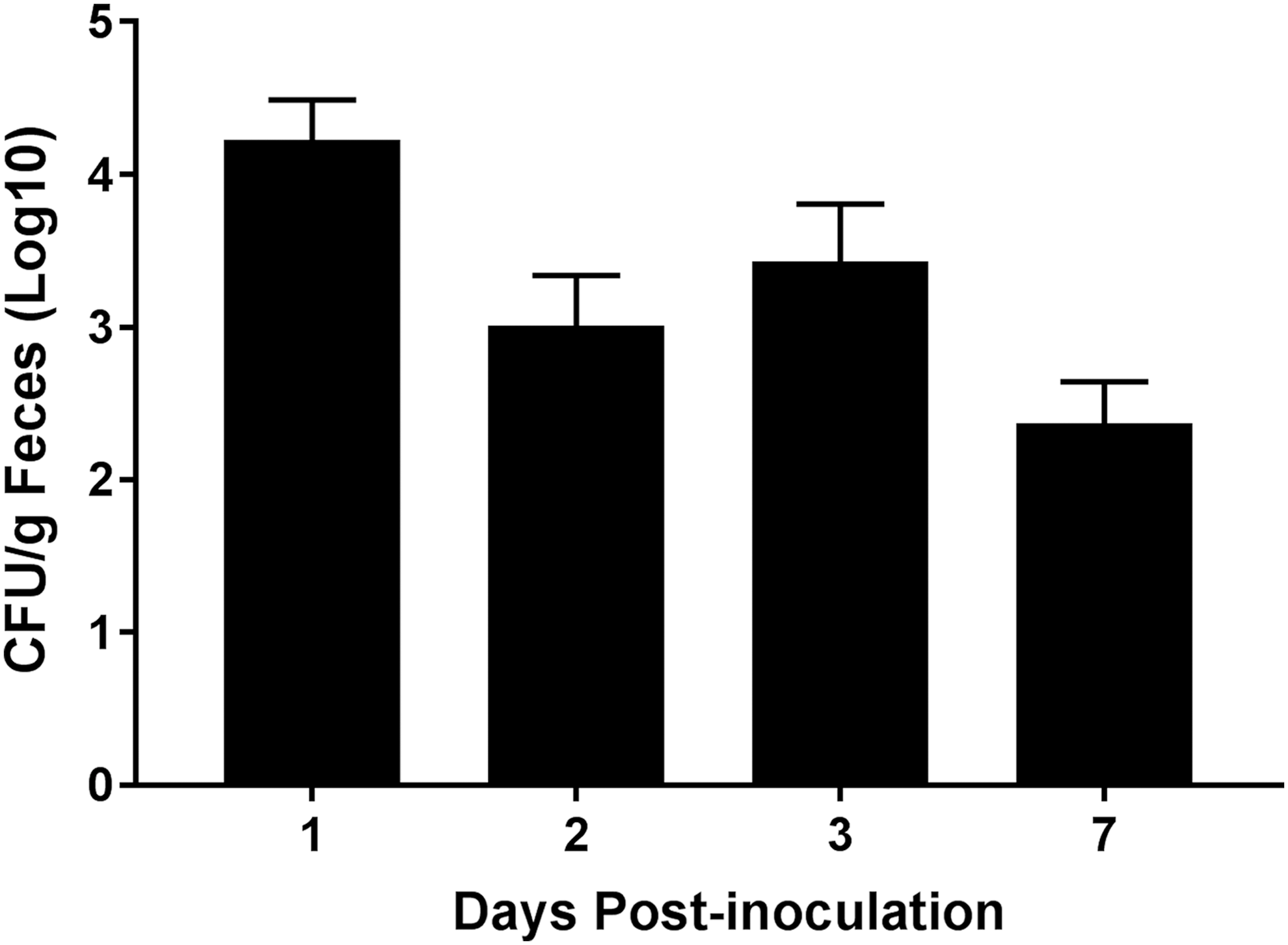
Fecal shedding of MDR Salmonella enterica serovar I 4,[5],12:i:- in pigs. Following inoculation with MDR serovar 1,4,[5],12:i:-, fecal shedding at 1, 2, 3, and 7 d.p.i. was measured. Data are expressed as arithmetic means with error bars indicating the standard experimental mean.
At 7 d.p.i., tissues of the intestinal tract and tonsils were evaluated for SX 240 colonization. Colonization levels of ≥103 CFU/g were observed in the following tissues associated with the ileum: Peyer's patch region, mucosal scrapings at the ileocecal junction, and ICLN (Fig. 4). Colonization levels between 102 and 103 CFU/g were detected in the cecal tissue and contents, whereas the tonsils were colonized at 101 CFU/g. These data suggest that MDR serovar I 4,[5],12:i:- can colonize pig tissues, including the tonsils, which have been implicated in Salmonella carriage (Boyen et al., 2008a, b; Van Parys et al., 2010; Volf et al., 2012). Carrier-status pigs have been shown to be a major reservoir for Salmonella transmission, as stressful events such as transport, farrowing, or weaning can induce pathogen recrudescence, resulting in increased intestinal load and fecal shedding of Salmonella (Callaway et al., 2006; Verbrugghe et al., 2011).
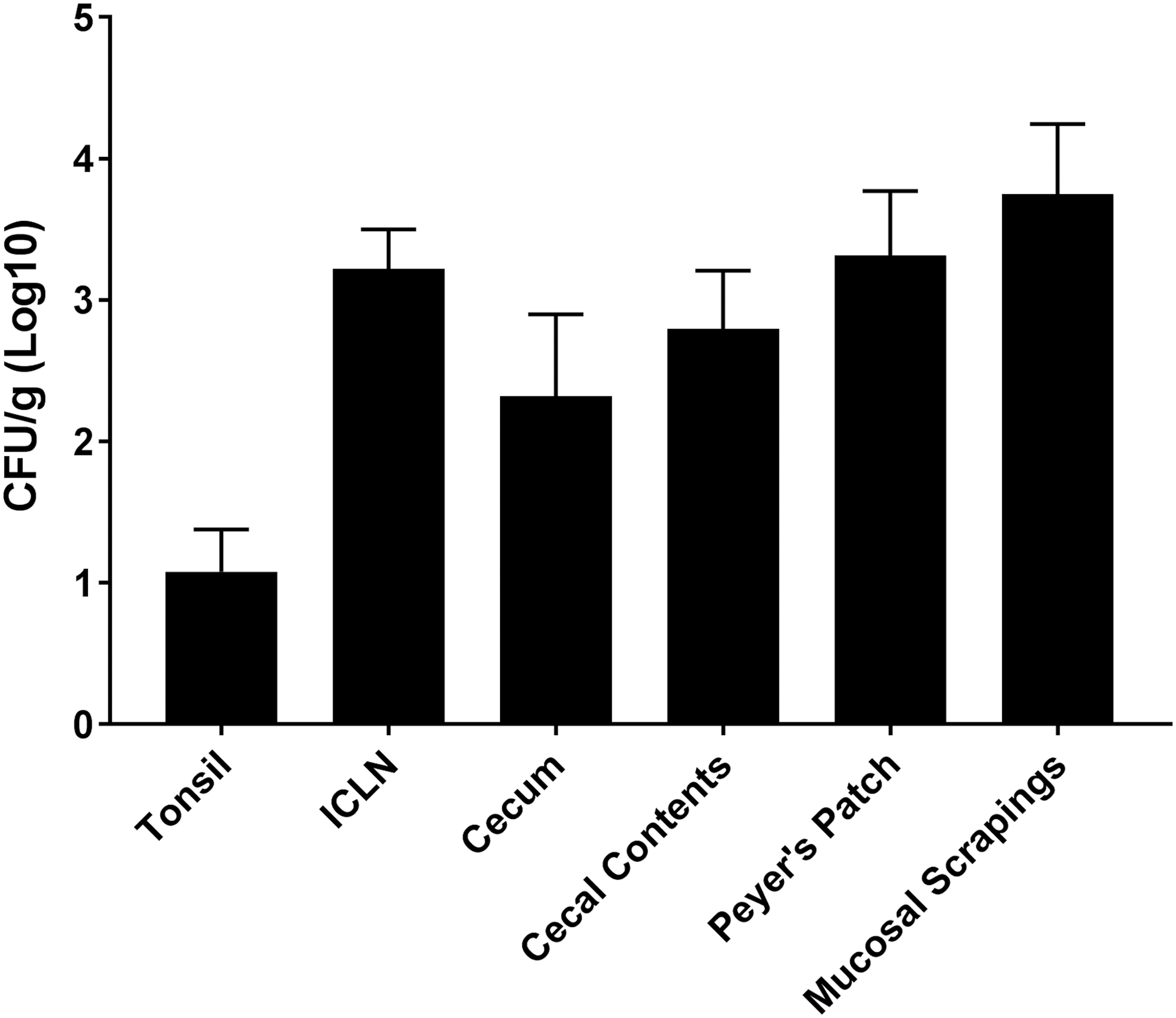
Tissue colonization of MDR Salmonella enterica serovar I 4,[5],12:i:- in pigs. Bacteriological analysis of MDR serovar I 4,[5],12:i:- (CFU/g) recovery from pig tissues at 7 d.p.i. was determined. Data are expressed as arithmetic means with error bars indicating the standard experimental mean. CFU, colony-forming units; ICLN, ileocecal lymph nodes.
Certain Salmonella mutants impaired for bacterial motility are less invasive than wild-type strains, including Salmonella Typhimurium SL1344 fliC fljB mutants that have decreased invasion of porcine intestinal epithelial cells (IPEC-J2) (Horstmann et al., 2017). However, SL1344 expressing either FliC alone or FljB alone had similar invasiveness to wild-type SL1344, indicating that expression of either flagellin is sufficient to permit IPEC-J2 invasion. Because invasion of intestinal epithelial cells is an important Salmonella virulence mechanism, invasion potential of SX 240 was compared to non-MDR, biphasic Salmonella Typhimurium strain χ4232 (SX 117). No significant difference (p = 0.2277) in IPEC-J2 invasion was observed between SX 240 (1.6%) and SX 117 (1.2%). Thus, the absence of fljB in MDR serovar I 4,[5],12:i:- did not reduce invasion of IPEC-J2 cells, supporting a previous in vitro study whereby a monophasic Salmonella DT193 isolate adhered to and invaded porcine intestinal epithelial cells at a comparable level to a biphasic Salmonella DT193 isolate (Crayford et al., 2014).
Overall, these data indicate that SX 240 colonizes porcine gastrointestinal tissues comparable to virulent, non-MDR, biphasic Salmonella Typhimurium (Bearson et al., 2014, 2017) and suggest that the absence of the second-phase flagellar antigen, FljB, does not limit its colonization potential. In this strain and in the European MDR serovar I 4,[5],12:i:- isolate with R-type ASSuT (DT193/STYMXB.0131), a 16-kb deletion of the genetic region encoding FljB is due to the insertion of a 28-kb cassette that carries antimicrobial resistance genes (Garcia et al., 2016). It remains to be determined whether deletion of a gene(s) in the fljB genomic region or presence of a gene(s) on the 28 kb module provides an advantage to serovar I 4,[5],12:i:- for persistent colonization or environmental survival that may account for its increasing prevalence over the last decade.
Porcine immune responses to MDR serovar I 4,[5],12:i:-
Our group has shown an association between variations in the levels of the proinflammatory cytokine IFN-γ with differing Salmonella shedding levels in swine (Huang et al., 2011). Inoculation of pigs with SX 240 significantly increased serum IFN-γ at 2 (369 pg/mL; p ≤ 0.0001) and 3 d.p.i. (230 pg/mL; p < 0.01) compared with day 0 (29 pg/mL) (Fig. 5), indicating that MDR serovar I 4,[5],12:i:- elicits a significant IFN-γ response in swine.
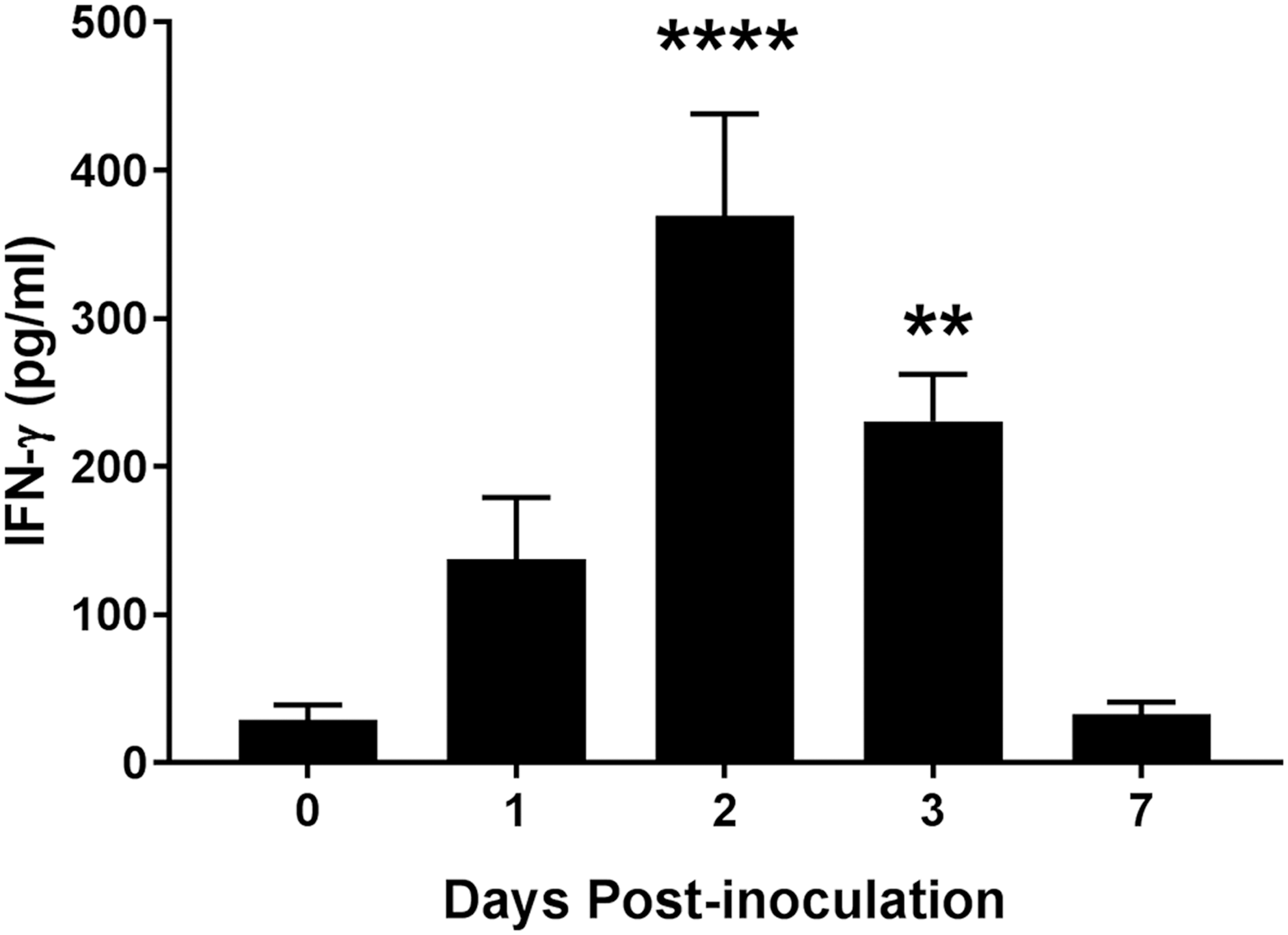
Significantly increased circulating IFN-γ levels in swine inoculated with MDR Salmonella enterica serovar I 4,[5],12:i:-. Serum IFN-γ levels at 0, 1, 2, 3, and 7 d.p.i. were determined by ELISA. Data are expressed as arithmetic means with error bars indicating the standard experimental mean. ****p ≤ 0.0001, **p < 0.01 when compared to day 0. IFN-γ, interferon-gamma.
Serological surveillance is an established method to screen swine herds for Salmonella exposure (Mousing et al., 1997; Alban et al., 2002; Sandberg et al., 2002). To determine if SX 240-inoculated pigs produce detectable antibodies to Salmonella LPS antigen, porcine sera were evaluated using the IDEXX HerdChek Swine Salmonella Test Kit. At 0 and 7 d.p.i., all pigs tested negative and positive for antibodies to Salmonella, respectively (S/P ratios <0.25 compared to S/P ratios of 2.8, 2.9, 1.7, 2.7, 0.8, 1.1, and 2.0), indicating detectable seroconversion in the SX 240-inoculated pigs within 7 days. Furthermore, the standard ELISA assay used for Salmonella surveillance can be used to detect MDR serovar I 4,[5],12:i:- in commercial pig operations.
Alterations in the swine fecal microbiota following MDR serovar I 4,[5],12:i:- inoculation
The effect of SX 240 inoculation on the swine fecal microbiota over the 7-day study was assessed using 16S rRNA gene sequencing. All pigs had a typical taxonomic profile with a predominance of Alloprevotella, Lactobacillus, Prevotella, and Treponema throughout the study (Holman et al., 2017) (Fig. 6). Fecal bacterial community structure analysis showed that pig samples clustered largely by sampling day (R 2 = 0.32; p < 0.001; Fig. 7). The swine fecal microbiota was significantly altered 1-day after Salmonella inoculation (R 2 = 0.19; p < 0.01), and the largest shift, based on weighted UniFrac distances, was observed between 0 and 7 d.p.i. (R 2 = 0.32; p < 0.01). Each sampling time had a similar dispersion as assessed by the betadisper function (p > 0.99). Relative abundance of certain genera was also significantly different between pre- and postinoculation samples. Phascolarctobacterium and Prevotellaceae NK3B31 group were more relatively abundant in preinoculation samples and Prevotella and Succiniclasticum in 1 d.p.i. fecal samples (LDA log10 ≥ 4.0). Moreover, the Phascolarctobacterium and Prevotellaceae NK3B31 group were enriched in the 0 d.p.i. samples compared with all other sampling times (LDA log10 ≥ 4.0). In contrast to our results, previous studies of pigs inoculated with Salmonella Typhimurium reported a decrease in the relative abundance of Prevotella at 2 d.p.i. (Bearson et al., 2013; Drumo et al., 2015). This discordance may reflect differences in the Salmonella strains or may also be a consequence of some Prevotella 16S rRNA gene sequences being classified as part of the closely related Prevotellaceae NK3B31 group.
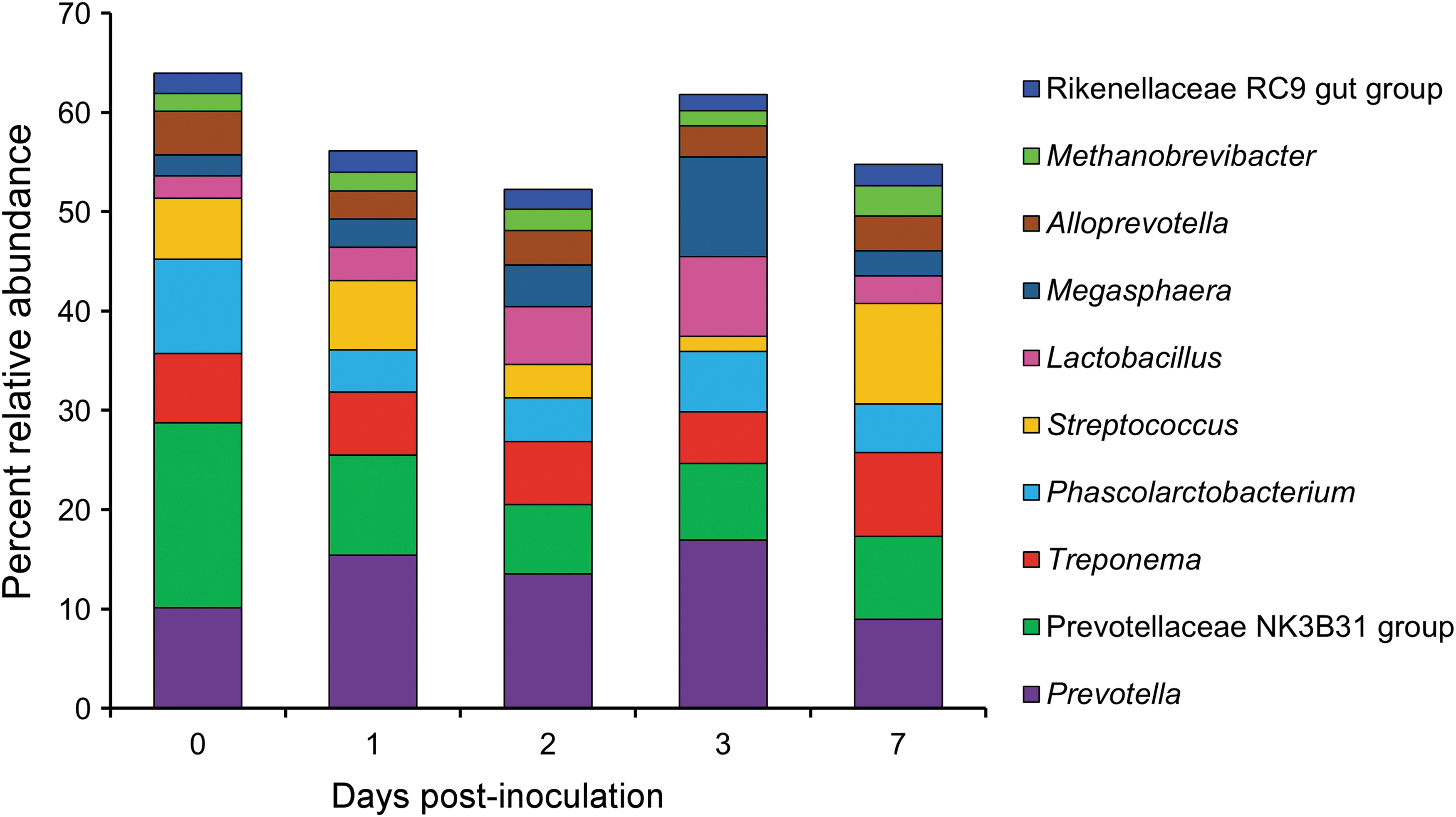
Relative abundance of the 10 most abundant archaeal and bacterial genera over the course of the study.
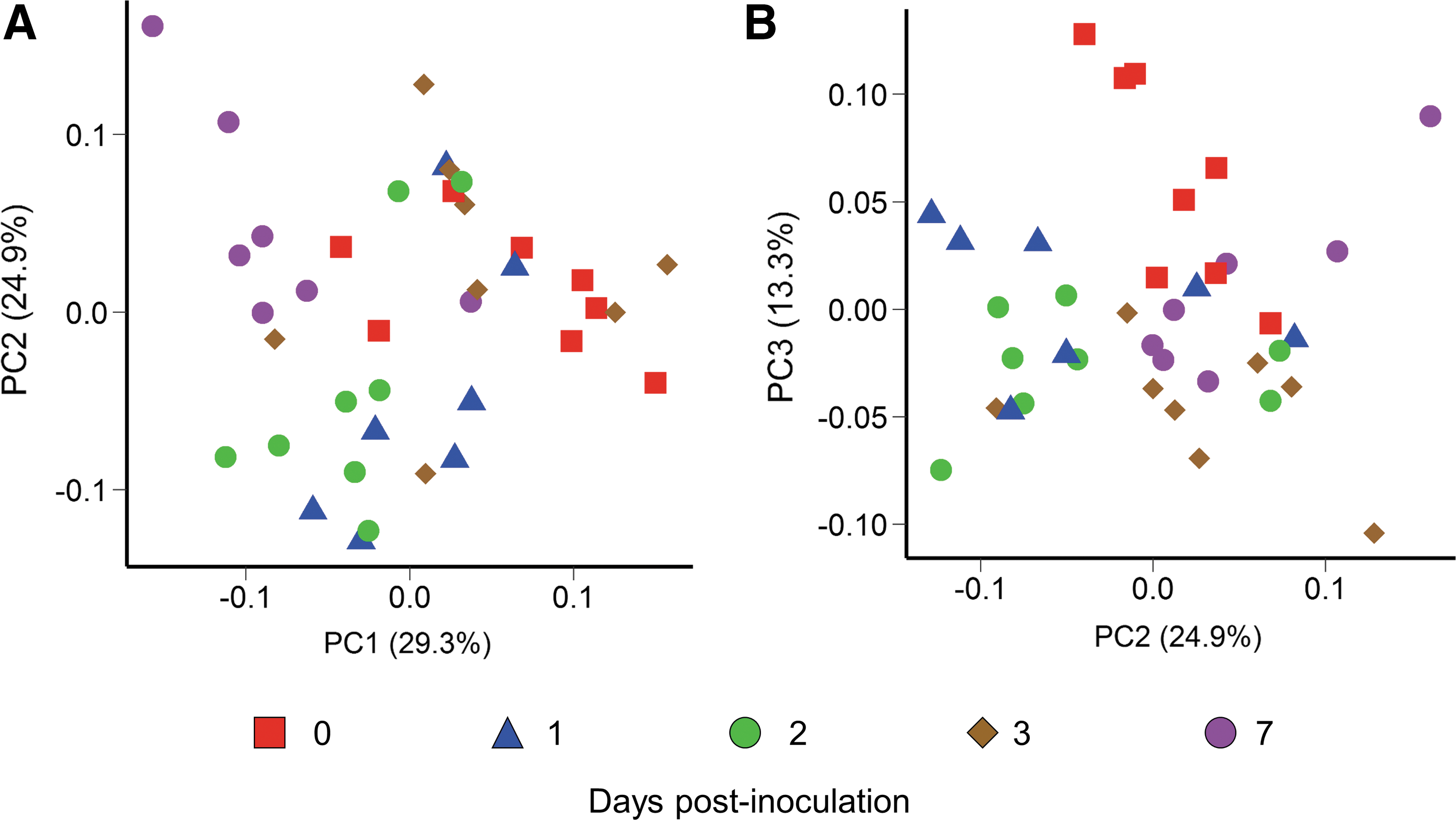
PCoA plot of the weighted UniFrac distances for each fecal sample by sampling time.
The enrichment of Succiniclasticum postinoculation was particularly striking as this genus was not detected in fecal samples before Salmonella inoculation. At 1 d.p.i., the relative abundance of Succiniclasticum was greater than 2.5% in all but two pigs. The genera Phascolarctobacterium and Succiniclasticum are both members of the Acidaminococcaceae family and produce the short chain fatty acid propionate from succinate (van Gylswyk, 1995). Thus, the observed relative increase in Succiniclasticum and decrease in Phascolarctobacterium at 1 d.p.i. may be a result of functional redundancy where membership is less important than the functional capacity of the microbiome.
Given that inoculation with Salmonella resulted in physiological changes within the pig after 1 day, including increased body temperature, diarrhea, and circulating IFN-γ levels, it is not surprising that the fecal microbiota also shifted during this period. Notably, the structure of the fecal microbiota had not returned to its initial or baseline (0 d.p.i.) state at 7 d.p.i., even though body temperature and circulating IFN-γ levels were no longer elevated. This finding is in agreement with a previous study by our group, whereby the fecal microbiota of high Salmonella Typhimurium shedding pigs remained perturbed even at 21 d.p.i. compared to mock-inoculated pigs (Bearson et al., 2013). An altered fecal microbiota may have a negative impact on swine production as this may be related to reports suggesting that Salmonella colonization in swine can reduce average daily gain (Funk and Gebreyes, 2004; Farzan and Friendship, 2010; Lynch et al., 2017).
Conclusions
Serovar I 4,[5],12:i:- has emerged as one of the most common nontyphoidal Salmonella serovars to cause human foodborne illness, and ∼50% of isolates are MDR. Our data indicate that a pork outbreak-associated MDR serovar I 4,[5],12:i:- isolate induced transient clinical disease in swine and perturbed the microbial community of the porcine gastrointestinal tract. In addition, fecal shedding and tissue colonization were consistent with previous swine experiments using virulent biphasic Salmonella Typhimurium, suggesting that the emergence of serovar I 4,[5],12:i:- in pork products is not due to increased virulence, colonization, or shedding characteristics during the acute stage of infection in the natural swine host. Further studies will determine if other mechanisms, such as stress response, metal tolerance, and biofilm formation, are responsible for the increased prevalence of monophasic Salmonella strains in swine.
Footnotes
Acknowledgments
The authors are grateful to Margaret Walker, Kellie Winter, and Jennifer Jones for outstanding technical assistance, and to David Alt for his assistance with the 16S rRNA gene sequencing. We are thankful to Crystal Loving for helpful discussions on experimental design. Salmonella enterica serovar I 4,[5],12:i:- isolates from swine were generously provided by Glenn Tillman (USDA, FSIS, Athens, GA) and John Johnston (USDA, FSIS, Fort Collins, CO). The IPEC-J2 cells were a kind gift from Dr. Bruce Schultz (Department of Anatomy and Physiology, Kansas State University, Manhattan, KS). This research was partially supported by the Iowa Pork Producers Association (NPB #16-113).
Ethics Statement
All animal procedures followed protocols approved by the USDA, ARS, NADC Animal Care, and Use Committee in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health.
Disclosure Statement
No competing or financial interests exist. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendations or endorsement by the U.S. Department of Agriculture. The U.S. Department of Agriculture is an equal opportunity provider and employer.