
Abstract
Multidrug-resistant (MDR) plasmids play an important role in disseminating antimicrobial resistance genes. To elucidate the antimicrobial resistance gene compositions in A/C incompatibility complex (IncA/C) plasmids carried by animal-derived MDR Salmonella Newport, and to investigate the spread mechanism of IncA/C plasmids, this study characterizes the complete nucleotide sequences of IncA/C plasmids by comparative analysis. Complete nucleotide sequencing of plasmids and chromosomes of six MDR Salmonella Newport strains was performed using PacBio RSII. Open reading frames were assigned using prokaryotic genome annotation pipeline (PGAP). To understand genomic diversity and evolutionary relationships among Salmonella Newport IncA/C plasmids, we included three complete IncA/C plasmid sequences with similar backbones from Salmonella Newport and Escherichia coli: pSN254, pAM04528, and peH4H, and additional 200 draft chromosomes. With the exception of canine isolate CVM22462, which contained an additional IncI1 plasmid, each of the six MDR Salmonella Newport strains contained only the IncA/C plasmid. These IncA/C plasmids (including references) ranged in size from 80.1 (pCVM21538) to 176.5 kb (pSN254) and carried various resistance genes. Resistance genes floR, tetA, tetR, strA, strB, sul, and mer were identified in all IncA/C plasmids. Additionally, blaCMY-2 and sugE were present in all IncA/C plasmids, excepting pCVM21538. Plasmid pCVM22462 was capable of being transferred by conjugation. The IncI1 plasmid pCVM22462b in CVM22462 carried blaCMY-2 and sugE. Our data showed that MDR Salmonella Newport strains carrying similar IncA/C plasmids clustered together in the phylogenetic tree using chromosome sequences and the IncA/C plasmids from animal-derived Salmonella Newport contained diverse resistance genes. In the current study, we analyzed genomic diversities and phylogenetic relationships among MDR Salmonella Newport using complete plasmids and chromosome sequences and provided possible spread mechanism of IncA/C plasmids in Salmonella Newport Lineage II.
Introduction
S
The emergence and spread of MDR Salmonella Newport is a long-standing public health concern in the United States (Welch et al., 2007; Fricke et al., 2009; Lindsey et al., 2009; Fernandez-Alarcon et al., 2011; Tadesse et al., 2016; Valenzuela et al., 2017). Extended-spectrum cephalosporin-resistant, for example, AmpC-MDR Salmonella Newport carrying blaCMY-2 shows resistance to multiple antibiotics, including ampicillin, chloramphenicol, streptomycin, cefoxitin, and ceftiofur (Sharma et al., 2017). The AmpC-MDR Salmonella Newport exclusively belongs to sequence types (STs) 45 and 116 in lineage II (Sangal et al., 2010). The AmpC-MDR Salmonella Newport we selected originated from the same most recent common ancestor in ST45 (Cao et al., 2013).
The A/C incompatibility complex (IncA/C) plasmids have been widely distributed (Kim et al., 2008; Pan et al., 2008). Such IncA/C plasmids have been found in various Gram-negative species, including Salmonella and Escherichia coli, playing important roles in the dissemination of resistance genes (Welch et al., 2007; Harmer and Hall, 2015). The IncA/C plasmids harbored by different bacterial species have a similar backbone, but often carry different resistance genes because of mobile elements (Welch et al., 2007; Fricke et al., 2009; Call et al., 2010; Wiesner et al., 2011). The typical carriers of the resistance gene blaCMY-2 in Salmonella enterica are IncA/C and IncI1 plasmids (Call et al., 2010; Folster et al., 2014, 2017; Tagg et al., 2014). Our plasmids belong to ST 1 IncA/C2 plasmids (Harmer and Hall, 2015). There are two resistance regions in IncA/C plasmids: ARI-A (antibiotic resistance island) and ARI-B, which were also known as aadA (aadA, aacC, qacE, and sul1) and floR (floR, tetAR, strAB, and sul2) regions, respectively. ST2, ST12, and ST23 are the major types in IncI1 plasmids carrying blaCMY-2 (Tagg et al., 2014). The blaCMY-2 region had been mobilized from an IncA/C plasmid through insertion sequence IS1294 (Tagg et al., 2014).
Clustered regularly interspaced short palindromic repeats (CRISPRs)/cas system has been identified in ∼45% of bacteria, including Salmonella, which are adaptive immune systems against foreign mobile elements, such as plasmids and phages (Stout et al., 2017). A functional CRISPR/cas system has the ability to target and degrade certain plasmids containing resistance genes. Bacteria carrying nonfunctional CRISPR/cas system could acquire plasmids carrying resistance genes (Palmer and Gilmore, 2010; Price et al., 2016; Gawryszewska et al., 2017). The complete function of CRISPR/cas system reversely correlates to antimicrobial resistance in bacteria (Palmer and Gilmore, 2010; Price et al., 2016; Gawryszewska et al., 2017).
The application of Whole Genome Sequencing (WGS) affords the opportunity to reveal diversity and genetic characteristics of plasmids and chromosomes of bacterial pathogens (Lienau et al., 2011; Ju et al., 2012; Zhang et al., 2016). WGS database provides comprehensive data with a broader horizon on research, such as GenomeTrakr network established by the U.S. Food and Drug Administration (FDA) and Pathogen Detection initiated by National Center for Biotechnology Information (NCBI).
We characterized the evolutionary relationships and diversities of Salmonella Newport lineages II and III, including the six Lineage II MDR strains, which we further characterized in the current study (Cao et al., 2013). However, the sequence diversity, evolution, and mobilization mechanism of Salmonella Newport IncA/C plasmids are not totally understood. The objectives of this study were to further characterize the sequence diversity of Salmonella Newport IncA/C plasmids harboring blaCMY-2 by comparative analysis and to understand the evolutionary relationship of IncA/C plasmids.
Materials and Methods
Bacterial strains
Six MDR Salmonella Newport strains were selected, including one from ground beef (CVMN1543, Georgia) and five from sick animals (CVM22425, cattle, Arizona; CVM22462, canine, Arizona; CVM22513, cattle, North Carolina; CVM21538, chicken, Georgia; and CVM21550, swine, Texas) (Cao et al., 2012, 2013; Cao et al., 2015).
Antimicrobial susceptibility
The minimum inhibitory concentration (Sharma et al., 2017) values of 16 antimicrobials were determined using a Sensititre system with custom plates (catalog no. CMV2AGNF; Thermo Scientific, Westlake, OH). Antimicrobial resistance was interpreted according to the Clinical and Laboratory Standards Institute (CLSI) criteria with exception of streptomycin (Institute, 2012). The antimicrobials (resistance breakpoint) included amoxicillin–clavulanic acid (AMC, ≥32 μg/mL), ampicillin (AMP, ≥32 μg/mL), azithromycin (AZI, ≥32 μg/mL), cefoxitin (FOX, ≥32 μg/mL), ceftiofur (TIO, ≥8 μg/mL), ceftriaxone (AXO, ≥4 μg/mL), chloramphenicol (CHL, ≥32 μg/mL), ciprofloxacin (CIP, ≥1 μg/mL), nalidixic acid (NAL, ≥32 μg/mL), trimethoprim–sulfamethoxazole (COT, ≥4 μg/mL), sulfisoxazole (SUL, ≥512 μg/mL), tetracycline (TET, ≥16 μg/mL), gentamicin (GEN, ≥16 μg/mL), and kanamycin (KAN, ≥64 μg/mL). The breakpoint for streptomycin (STR) is ≥64 μg/mL. We included the following as quality control organisms: E. coli ATCC25922, Enterococcus faecalis ATCC 29212, Staphylococcus aureus ATCC 29213, and Pseudomonas aeruginosa ATCC 27853.
Sequencing, assembly, and annotation
The relating methods of sequencing, assembly, and annotation were described as published (Klimke et al., 2009; Cao et al., 2015; Maria Hoffmann et al., 2017). Three published IncA/C plasmid sequences were selected as references: pSN254 (NC_009140), pAM04528 (FJ621587), and peH4H (FJ621586). Plasmid pJIE512b (NC_025198) was used as the IncI1 reference plasmid.
Plasmid sequence analysis
Incompatibility group identification
The Inc type of each plasmid was determined using target sequences as described (Johnson and Nolan, 2009; Folster et al., 2012). Identification of repA for IncA/C and repZ for IncI1 was performed using BLAST 2.2.29 (Altschul et al., 1990) (
Plasmid MLST of IncI1 plasmid
The IncI1 plasmid ST was determined as described (Garcia-Fernandez et al., 2008) and in PubMLST database (
IncA/C plasmids analyses
We compared the genetic organization of nine IncA/C plasmids in the BLAST Ring Image Generator (upper limit: 90%; lower limit: 70%) by using pSN254 as reference sequence (Alikhan et al., 2011). We built a presence/absence matrix (omitting nonresistance genes shared by all compared plasmids and/or plasmid-specific genes) based on gene clusters using USEARCH with 90% sequence identity as the threshold (Edgar, 2010). A maximum likelihood tree was constructed using RAxML (ML+thorough bootstrap with 10 runs and 1000 bootstrap replicates) based on the binary matrix (Stamatakis, 2006; Daniele Silvestro, 2012). Plasmid peH4H was used as the outgroup.
Insertion sequence mobile elements
The ISFinder (
Conjugation experiments
We used E. coli JM109 (Promega, Madison, WI) (nalidixic acid resistant) as recipient strain. Nalidixic acid (25 μg/mL) and tetracycline (16 μg/mL) were used as selection agents. The conjugation experiments and efficiency calculation were conducted as described (Phornphisutthimas et al., 2007). One triparental mating experiment was performed using JM109 as recipient and E. coli HB101 carrying plasmid pRK2013 (ATCC37159) (kanamycin resistant) as helper (Van Meervenne et al., 2012).
Chromosomal sequence analysis
Phylogenetic analysis of complete genomes
Genomic data of strain AM04528 was not publicly available. Multiple sequence alignment for complete chromosomal nucleotide sequences was performed using progressiveMauve (Darling et al., 2010) with Salmonella Newport SL254 (CP001113) as a reference genome. Salmonella Newport CVMN18486 (ground beef, New Mexico) was outgroup for the phylogenetic analysis (Cao et al., 2013). A maximum likelihood tree was constructed using RAxML based on the single nucleotide polymorphisms (SNPs), which were identified from multiple genome alignments (ML+thorough bootstrap with 10 runs and 1000 bootstrap replicates) (Stamatakis, 2006; Daniele Silvestro, 2012).
Phylogenetic analysis of additional draft genomes from Pathogen Detection
We selected additional 30 draft genomes from Pathogen Detection by downloading SRA and/or FASTQ data (Supplementary Table S2; Supplementary Data are available online at
Pairwise distance matrix
Pairwise distance matrix (counting the number of nucleotide differences with standard deviation) was calculated for complete chromosomes using Molecular Evolutionary Genetics Analysis v7 with 1000 bootstrap replicates (Kumar et al., 2016).
cas3 gene sequences
The nucleotide sequence of the cas3 gene from CVMN18486 was used as reference to identify variations. The alignment was performed using MUSCLE (Edgar, 2004) in SeaView4 (Gouy et al., 2010). Meanwhile, we used BLAST 2.2.29 (
Nucleotide sequence accession numbers
The nucleotide sequences were deposited in GenBank (Cao et al., 2015). The accession numbers used in this study are as follows: CVM22425 (CP010279), CVM22462 (CP010280), CVM21550 (CP010283), CVM22513 (CP010281), CVMN1543 (CP010284), CVM21538 (CP010282), CVMN18486 (CP009561), pCVM22425 (CP009560), pCVM22462 (CP009567), pCVM22462b (CP009566), pCVM21550 (CP009564), pCVM22513 (CP009562), pCVMN1543 (CP009570), and pCVM21538 (CP009563).
Results
Antimicrobial susceptibility profiles
The six MDR Salmonella Newport strains exhibited different antimicrobial resistance phenotypes (Table 1 and Supplementary Table S1). CVM22425, CVM22462, CVMN1543, and CVM21550 exhibited resistance to nine antibiotics. CVM22513 exhibited additional resistance to kanamycin (Liakopoulos et al., 2016). CVM21538 was only resistant to four antibiotics. All strains were susceptible to azithromycin, ciprofloxacin, nalidixic acid, and trimethoprim–sulfamethoxazole.
Strain H4H is Escherichia coli strain.
All plasmids contained the mer operon (merA, merB, merD, merE, merP, merR, merT), conveying resistance to mercury ions.
AMC, amoxicillin–clavulanic acid (β-Lactamase inhibitors, ≥32 μg/mL); AMP, ampicillin (β-Lactamase inhibitors, ≥32 μg/mL); FOX, cefoxitin (Cephamycin antibiotic, ≥32 μg/mL); TIO, ceftiofur (Cephalosporin, ≥8 μg/mL); AXO, ceftriaxone (Cephalosporin, ≥4 μg/mL), STR, streptomycin (Aminoglycosides, ≥64 μg/mL); KAN, kanamycin (Aminoglycosides, ≥64 μg/mL); GEN, gentamicin (Aminoglycosides, ≥16 μg/mL); TET, tetracycline (Tetracyclines, ≥16 μg/mL); CHL, chloramphenicol (Chloramphecicol, ≥32 μg/mL); SUL, sulfonamide (Sulfa antibiotics, ≥512 μg/mL). The values presented are breakpoints.
IncA/C, A/C incompatibility complex.
Characteristics of IncA/C plasmids
All six MDR strains contained an IncA/C plasmid (Table 1). The nine IncA/C plasmids ranged from 80.1 to 176.5 kbp (Table 2) and the number of resistance genes ranged from 12 (pCVM21538) to 23 (peH4H), including six mercury resistance genes. All six sequenced plasmids contained ARI-B region, but none of ARI-A gene was present (Supplementary Figure S1). All identified resistance regions were found at the similar locations in these plasmids. Some plasmids were of similar sizes and genetic organization, such as pCVM22425 and pCVM22462. The composition and sizes of pSN254 and pCVM21538 varied significantly. All plasmids but pCVM22462 were not capable of self-conjugation.
E. coli H4H used for comparison.
ORF, open reading frames; HPG, hypothetical protein genes; NA, not available; IncA/C, A/C incompatibility complex.
Besides ARI-A and ARI-B, plasmids carried various resistance genes (Figs. 1 and 2). Genes blaCMY-2, blc, and sugE were mobilized as a group, which had a second copy. Plasmids pCVMN1543 (ground beef) and pCVM22425 (cattle) carried only one functional sugE. pCVMN1543 lacked one copy of blc and sugE. The second copy of sugE in pCVM22425 contained a frameshift (Table 3). pSN254 and peH4H carried both sul1 (ARI-A) and sul2 (ARI-B), whereas the remaining plasmids only carried sul2. The plasmids from Salmonella Newport isolates derived from cattle, canine, and swine carried two copies of blaCMY-2
, whereas pCVM21538 (chicken) did not contain blaCMY-2
. We investigated additional 200 MDR Salmonella Newport belonging to ST45 from the same SNP Cluster (PDS000002504.114) in Pathogen Detection (
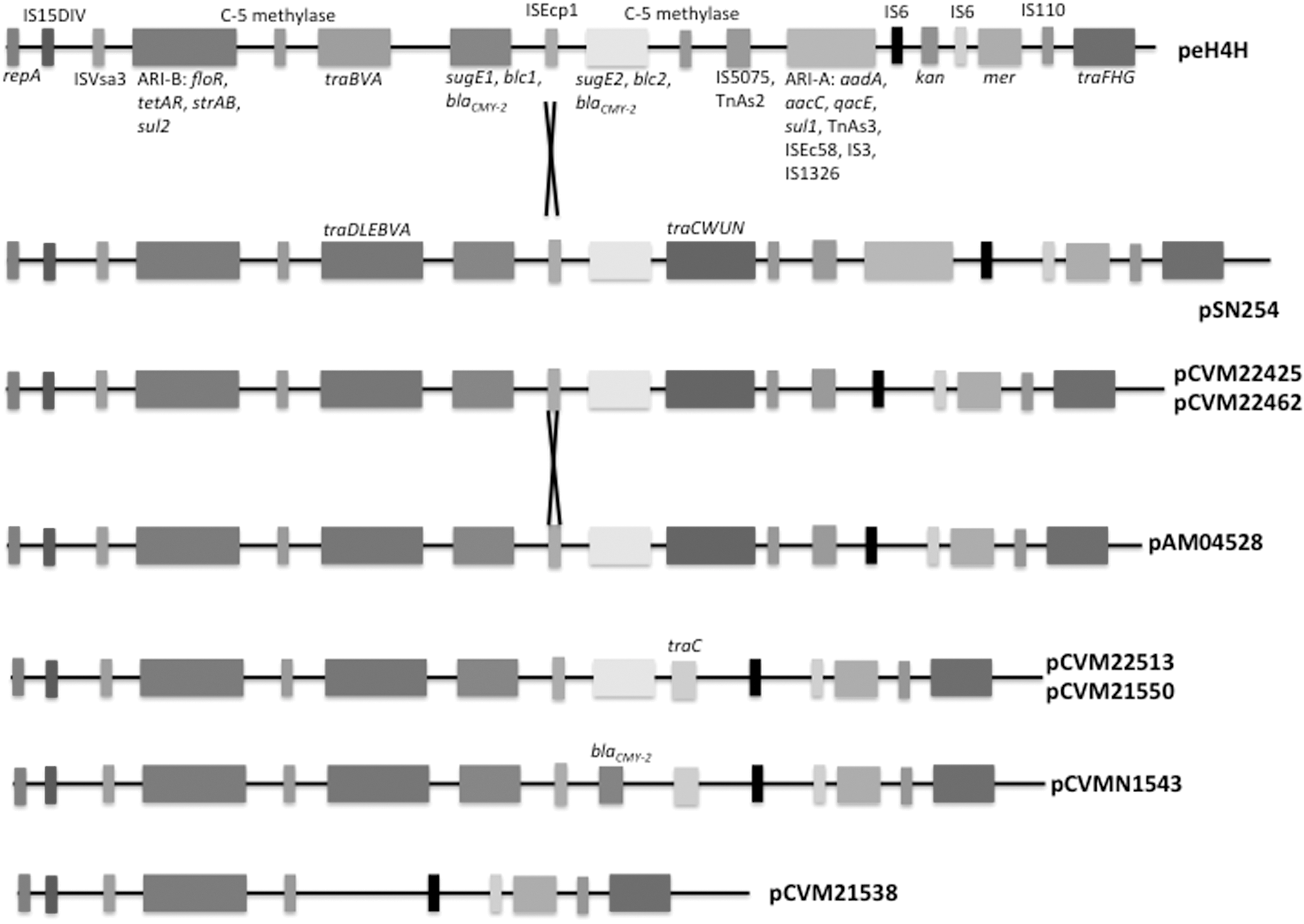
Schematic diagrams of compared complete sequences of IncA/C plasmids. Nine complete IncA/C plasmids analyzed in the current work shared a similar genetic backbone. Multiple resistance clusters having different distribution patterns were identified in the various plasmids. Same genes or gene clusters were marked on peH4H. If there were different compositions in resistance clusters, specific genes were labeled, such as traC in pCVM22513 and pCVM21550. IncA/C, A/C incompatibility complex.
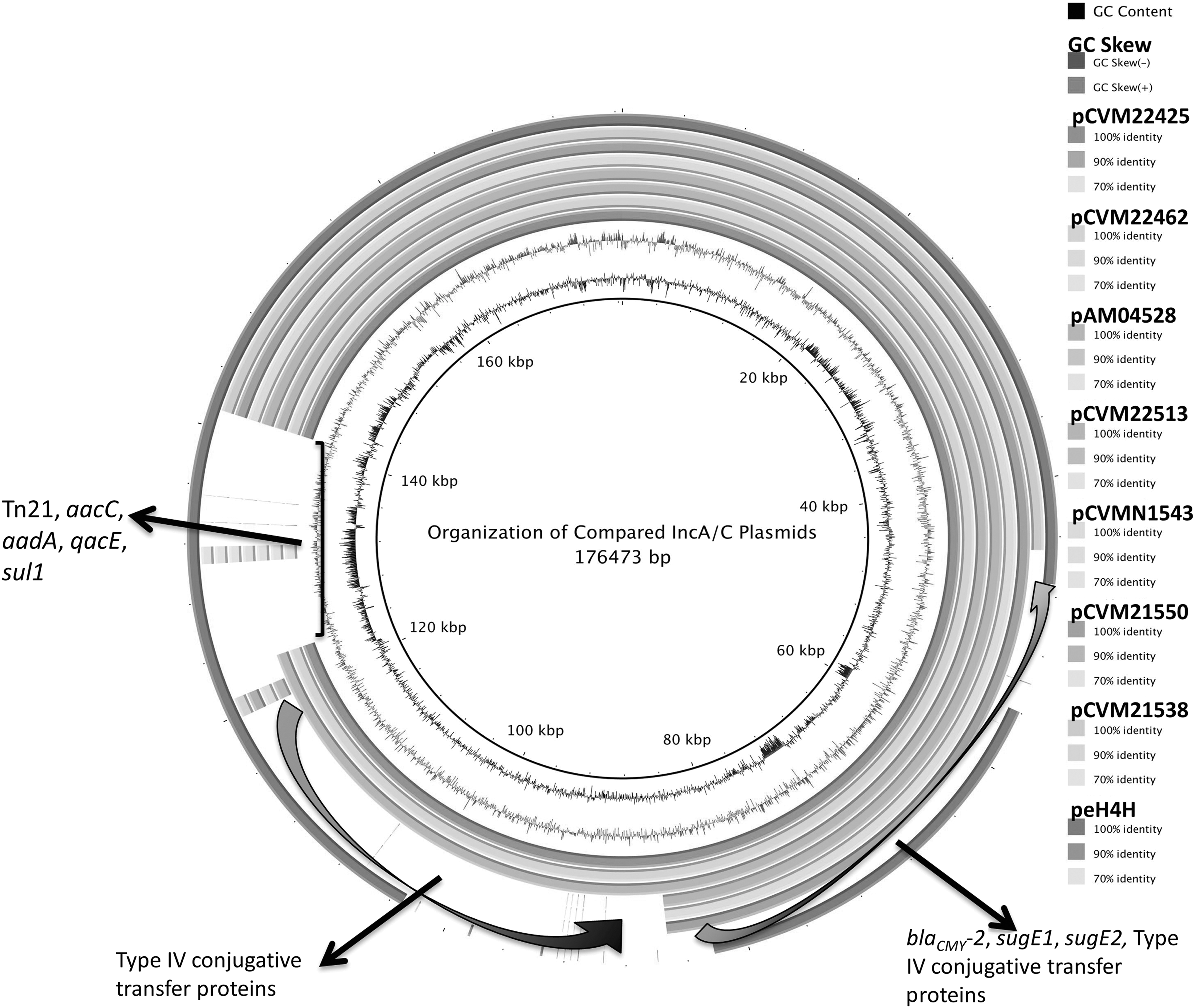
Organizational comparisons among the IncA/C plasmids of Salmonella Newport. Plasmid pSN254, in the inner circle, was used as the reference. Several regions in plasmid pSN254 were absent in other IncA/C plasmids, including blaCMY-2 , aacC, and type IV conjugative transfer proteins. IncA/C, A/C incompatibility complex.
position or locus in pSN254/query plasmid.
I, insertion compare to query plasmid; D, deletion; SNP, single nucleotide polymorphisms.
These plasmids displayed sequences highly conserved compared with pSN254 (Table 3). pCVM22425, pCVM22462, pCVM21538, and pCVMN1543 had no SNP in the nonresistance genes shared by all plasmids. pCVM22513, pCVM21550, and pAM04528 contained SNPs. For example, we identified one G to T mutation in pCVM21550, leading to an alanine to serine change in strB. pCVM22425 and pCVMN1543 contained single or multiple-base deletions resulting in frameshift. In pCVM22425, two deletions were found at positions 67,787 and 68,097 (sugE) and one deletion was identified at position 68,356 (blc).
We constructed one presence/absence matrix (Supplementary Table S3) based on all identified genes in these plasmids (Supplementary Table S4). Binary tree showed the relationships among these plasmids (Fig. 3). IncA/C plasmids with similar sizes, such as pCVM22513, pCVM21550, and pCVMN1543, clustered together.
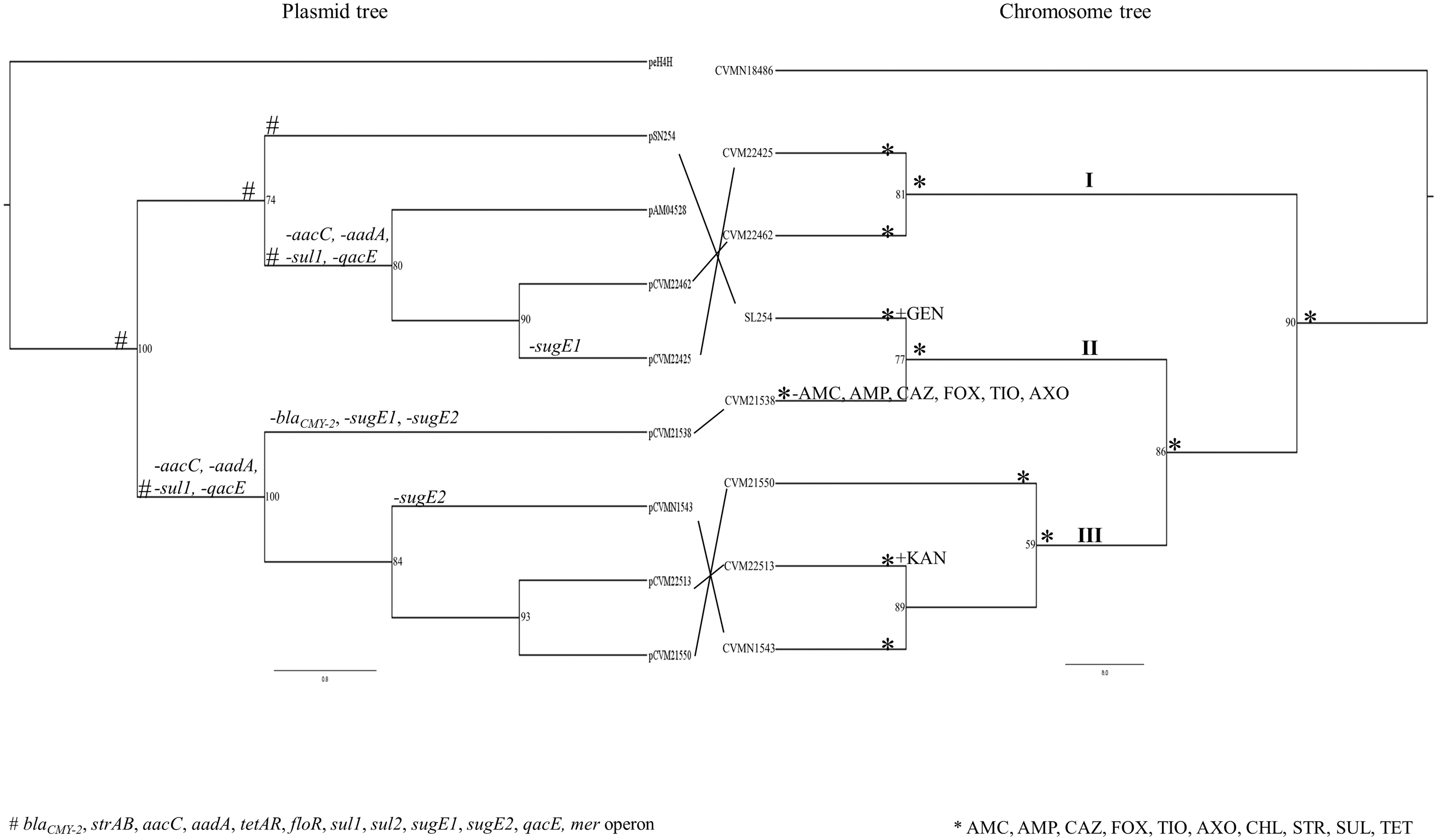
Phylogenetic tree of Salmonella Newport complete chromosomes and binary tree of IncA/C plasmids. The maximum likelihood tree of plasmids was constructed based on the presence/absence matrix of gene clusters in all IncA/C plasmids. Plasmid peH4H from Escherichia coli was selected as outgroup. The maximum likelihood tree of chromosomes was built based on 5448 single nucleotide polymorphisms of multiple genomes alignment. Strain CVMN18486 was selected as outgroup. The multidrug-resistant strains were divided into three subgroups. The IncA/C plasmids in subgroups I and III also clustered together in the plasmid tree, respectively. Antimicrobial resistance profiles and presence/absence of resistance genes in IncA/C plasmids were labeled in each tree. IncA/C, A/C incompatibility complex.
Mobile elements in IncA/C plasmids
We identified three regions of type IV conjugative transfer proteins: traIDLEBVA (I), traCWUN (II), and traFHG (III) (Fig. 1, Supplementary Table S5). Region III was present in all plasmids, which were adjacent to ARI-A usually. Region I was found in all plasmids, excepting pCVM21538. The traDLE genes in region I were missing in peH4H. Complete region II was found in pSN254, pCVM22425, pCVM22462, and pAM04528, but absent in pCVM21538 and peH4H. Plasmids pCVM21550, pCVM22513, and pCVMN1543 contained only traC in region II.
We identified 14 genes encoding transposases in pSN254 (Fig. 1, Supplementary Table S6). The sequenced plasmids did not carry five to eight of these genes. Transposases TnAs2 (insE), ISEc58 (insF), IS3 (insG), and IS1326 (istA and istB) were absent in all sequenced plasmids. IS5075 (insD1) and TnAs2 (tnpA) were absent in pCVM22513, pCVM21550, and pCVMN1543.
Characteristics of IncI1 plasmid
CVM22462 carried one additional ST23 IncI1 plasmid, pCVM22462b. The genetic organization was similar to that of pJIE512b in E. coli (Supplementary Fig. S2). In pCVM22462b, blaCMY-2 , sugE, and blc were adjacent to IS1294 and it carried a tra conjugal transfer cluster (traA through traY) and a trb conjugal transfer cluster (trbABC).
Analyses of Salmonella Newport chromosomes
MDR Salmonella Newport strains shared the most recent common ancestor with small SNP differences (Fig. 3). The smallest SNP difference was 11 SNPs (± 4 SNPs) between CVM22425 and CVM22462; the largest SNP difference was 133 SNPs (± 8 SNPs) between SL254 and CVM22513 (Table 4).
Standard deviation in parentheses.
IncA/C, A/C incompatibility complex.
Same as SL254, all sequenced MDR Salmonella Newport had a mutation in cas3 in CRISPR/cas system, containing a C to T substitution at position 1648 out of 2700 bp leading to a premature stop codon, which was annotated as a pseudogene (SNSL254_RS16015). However, Salmonella Newport lineage II-susceptible strains, including those belonging to ST45 in our published work did not contain this mutation (Supplementary Fig. S3).
Pathogen detection data analyses
All sequenced Salmonella Newport belonged to SNP Cluster PDS000002504.114, an ID of computed cluster based on SNP distance, containing 556 genomes. Distance between these isolates ranges from zero to 96 SNPs. We selected 200 ST45 genomes (Supplementary Table S2). Resistance genotype data showed that four genomes did not contain resistance genes as no plasmid data were available.
Meanwhile, we selected 30 draft and four complete MDR Salmonella Newport genomes (Supplementary Table S7). These genomes carried the identical mutation in cas3 (Supplementary Fig. S3). The phylogenetic tree (Supplementary Fig. S4) showed that closely related genomes contained similar resistance genotype profiles. For example, NY_22697 and NY_22699 carried the same resistance genes (Supplementary Table S2).
Discussion
Salmonella Newport IncA/C plasmids had a conserved backbone structure, but different antimicrobial resistance gene profiles with corresponding phenotypes (Welch et al., 2007; Call et al., 2010; Fernandez-Alarcon et al., 2011; Wiesner et al., 2011) (Figs. 1 and 2, Table 1). These resistance genes and associated mobile elements were grouped together (Welch et al., 2007; Partridge, 2011). ARI-B and ARI-A regions were found adjacent to insertion sequences, ISVsa3 (insA) and TnAs3 (insE), respectively. These two insertion sequences belonged to insertion sequence common region (ISCR) elements and were closely related to IS91 family (Toleman et al., 2006a, 2006b). ISCR elements have been found as important antimicrobial resistance gene capture and movement system (Toleman and Walsh, 2010). TnAs3 was recognized as ISCR2 (Toleman and Walsh, 2008), which was also found in the IncA/C reference plasmid pRA1, indicating ISCR2 associated with resistance gene acquisitions (Toleman and Walsh, 2010). ISCR elements have been key factors during the recent IncA/C plasmid family evolution for acquisitions (Toleman and Walsh, 2010).
As peH4H and Salmonella Newport IncA/C plasmid shared the same backbone structure, they were likely derived from a common ancestor. IncA/C plasmids under different environmental selective pressures could acquire different genetic elements and increase genomic diversities (Call et al., 2010). Different types of antibiotic administration on dosage had significant impact on the dissemination and persistence of IncA/C plasmids carrying blaCYM-2 (Subbiah et al., 2011; Johnson et al., 2015). Conversely, reducing or eliminating the selective pressure from antibiotics could lower the energy demands of the host cell, leading to the decay of unnecessary genes and/or even the loss of the plasmids (Subbiah et al., 2011). Moreover, absence of antibiotics would lead to a population dominated with sensitive organisms indicating that a certain level of selection pressure was necessary to keep MDR plasmids in bacterial population (Subbiah et al., 2011). Mobile elements in these plasmids provide the flexibility to acquire foreign genetic elements when selective pressures change (Fernandez-Alarcon et al., 2011; Johnson et al., 2011). Plasmid pR148 (Aeromonas hydrophila, JX141473) is considered as the precursor of IncA/C2 plasmids carrying blaCMY-2 and lacks ARI-B (Del Castillo et al., 2013; Harmer and Hall, 2014). ARI-B was found at the same locations indicating it was acquired only once (Harmer and Hall, 2015). pCVM21538 carrying ARI-B did not harbor blaCMY-2 and lacked around 40 kbps compared with pR148, suggesting it might subsequently evolve in an ongoing process after originating from pR148.
In Pathogen Detection, we did not find any correlation between the number/distribution of resistance genes and sources/locations. Closely related strains carried same or similar resistance genes. Although there has been no experimental work to elucidate IncA/C plasmids' stability (Harmer and Hall, 2015), resistance genes could be stable and disseminated among clonal groups from common sources or geographical locations. For example, FSIS1400138 and FSIS1400139 carried 12 resistance genes without SNP difference (Supplementary Table S2).
Plasmids in Salmonella Newport, Salmonella Senftenberg, and Salmonella Ohio were not capable of being transferred into a new host (Welch et al., 2007; Evershed et al., 2009). IncA/C plasmids carrying blaCMY-2 had various conjugative abilities. Conjugation efficiency for pAM04528 and peH4H was less than 10−7, whereas pAR060302 has 10−3 (Subbiah et al., 2011). In this study, all IncA/C plasmids failed to self-conjugate, except pCVM22462. We compared pCVM22462 and pCVM22425. Although they carried the same genes encoding conjugation transfer proteins, CVM22462 contained an additional IncI1 plasmid. It was possible that the conjugation of pCVM22462 resulted from the conjugation machinery in IncI1 plasmid. Moreover, we did not find any differences within the transcriptional activator complex AcaCD between pCVM22462 and pCVM22425, which was essential for conjugative transfer (Carraro et al., 2014). Therefore, further experimental work is required to determine the Tra proteins' function and to investigate the role of additional IncI1 plasmid. (Poole et al., 2009; Call et al., 2010; Fernandez-Alarcon et al., 2011; Wiesner et al., 2011).
From an evolutionary perspective, the most recent common ancestor of these MDR ST45 Salmonella Newport strains had acquired IncA/C plasmid through horizontal gene transfer from different serotypes or other species (Sangal et al., 2010; Cao et al., 2013). The closely evolutionary relationship between AmpC-MDR Salmonella Newport in Lineage II and the exclusive presence of IncA/C plasmid carrying blaCMY-2 (except for CVM21538) suggested that these MDR Salmonella Newport share specific genetic advantages or mechanisms for the acquisition, which is not completely understood. One such possibility might lie in the CRISPR/cas systems of this particular Salmonella Newport ST. As CRISPR plays an important role against foreign genetic elements (Price et al., 2016), we investigated the CRISPR/cas system, including additional 34 public genomes (Supplementary Table S2). The cluster-exclusive mutation we identified in cas3 lead to a pseudogene, which may cause incomplete functional CRISPR that could affect DNA targeting and degradation of foreign DNA (Hochstrasser et al., 2014). Consistent with this notion, it was reported that the presence of a functional CRISPR/cas system was inversely correlated to the acquisition and maintenance of antimicrobial resistance in bacterial pathogens (Palmer and Gilmore, 2010). Therefore, the finding offered insights on possible mechanisms by which the common ancestor of AmpC-MDR Salmonella Newport acquired IncA/C plasmid.
Footnotes
Acknowledgments
This work was supported in part by the Joint Institute for Food Safety and Applied Nutrition, the University of Maryland. The authors thank Mrs. Sherry Ayers and Mr. Jason Abbott from the Center of Veterinary Medicine, U.S. FDA for their help in antibiotic susceptibility test. They thank Dr. Lili Fox Velez, Office of Research Sciences at CFSAN, FDA, for her scientific writing and editing support.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.