
Abstract
The adherence and biofilm formation of Staphylococcus aureus on food contact surfaces are a major concern for the food industry. Development of antibiofilm agents from polyphenols has drawn much attention due to their potent activity. The present study explored the antibacterial and antibiofilm activities of 2R,3R-dihydromyricetin (DMY) against S. aureus ATCC 29213. It was found that DMY exerted excellent antibacterial and bactericidal properties against S. aureus with minimum inhibitory concentration and minimum bactericidal concentration values of 0.125 and 0.25 mg/mL, respectively. Crystal violet staining and 2,3-bis (2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide sodium salt reduction assay demonstrated that DMY significantly reduced the biofilm biomass of S. aureus and decreased the metabolic activity of biofilm cells. Micrographs of light microscope and scanning electron microscope confirmed that DMY inhibited the biofilm formation and caused a disintegration of the complex biofilm architecture. Moreover, DMY was highly efficient in reducing the number of sessile S. aureus cells adhered to stainless steel. These results suggested that DMY could have potential application to control S. aureus contamination in a food processing environment.
Introduction
B
Adequate hygiene and disinfections are common strategies applied to remove attached pathogens and prevent biofilm formation on the food contact surfaces. Various chemical sanitizers have been used in the disinfection process, including chlorine, chlorhexidine, hydrogen peroxide, peracetic acid, and quaternary ammonium compounds (Silva et al., 2017). The ease of application, relative low cost, and broad antimicrobial spectrum of these commercial disinfectants allow for their extensive application in the food processing procedures. However, increasing awareness of their negative effects on human health and environments has led to the development of antibiofilm agents from natural sources.
Cedrus deodara (Roxb.) Loud., which belongs to family Pinaceae, is widely utilized as a traditional Chinese herb medicine. C. deodara has been reported to possess various nutritional and pharmaceutical effects, and its extract showed excellent antimicrobial activities against a broad range of foodborne pathogens (Zeng et al., 2012). 2R,3R-dihydromyricetin (DMY), a flavonoid isolated from needles of C. deodara, showed a variety of bioactivities, including hypoglycemic, hepatoprotective, analgesic, antioxidant, anti-inflammatory, antitumor, and antibacterial activities (Zhang et al., 2007; Cao et al., 2017). Of particular interest, DMY has been reported to exhibit potent antibacterial activity against S. aureus, Bacillus subtilis, Aspergillus flavus, and Vibrio parahaemolyticus (Kou and Chen, 2012; Liu et al., 2016; Wu et al., 2017). Nevertheless, there has been limited literature about the possible inhibitory effect of DMY on bacterial biofilm formation, especially against S. aureus.
Therefore, the aim of this study was to investigate the antibiofilm activity of DMY against S. aureus by assessing the biofilm biomass, metabolic activity of biofilm cells, and morphology of bacterial biofilm. Furthermore, the potential use of DMY to inactivate the sessile S. aureus cells adhered to stainless steel was evaluated.
Materials and Methods
Chemicals, bacteria strain, and culture conditions
DMY (HPLC ≥98%) was purified from needles of C. deodara as our previous reported method (Liang et al., 2014). A stock solution of DMY in 25% dimethyl sulfoxide (DMSO) was diluted with corresponding medium before use to keep the final concentration of DMSO <0.25%. S. aureus ATCC 29213 was obtained from the China Medical Culture Collection Center (Beijing, China). The bacteria were inoculated into tryptone soy broth (TSB) supplemented with 1% glucose and incubated overnight at 37°C to give the logarithmic phase cells for biofilm assays. Bacterial suspension at a cell density of 1 × 109 colony-forming unit (CFU)/mL was prepared with corresponding medium according to OD600 of 0.18.
Determination of minimum inhibitory concentration and minimum bactericidal concentration
The minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) values of DMY against S. aureus ATCC 29213 were determined by the broth microdilution method as previously reported (Wu et al., 2017). The culture medium used in this assay was Mueller–Hinton broth.
Biofilm formation
The biofilm of S. aureus ATCC 29213 was prepared in 96-well polystyrene microplates with a modified method (Nostro et al., 2016). In brief, overnight culture of S. aureus was diluted with TSB supplemented with 1% glucose to obtain a bacterial suspension of 1 × 106 CFU/mL. Then, 100 μL of the bacterial suspension and 100 μL of serial twofold dilutions of DMY in the same culture were added into a sterile 96-well microplate. TSB supplemented with 1% glucose containing 0.25% DMSO was incorporated as the negative control. The plate was incubated at 37°C for 24 h to allow biofilm formation.
Measurement of growth inhibition and biofilm biomass
After incubation, the optical density at 600 nm was measured to evaluate the effect of DMY on the planktonic bacterial growth, using a microplate reader (Spectra MAX-190; Molecular Devices Co.). Then, planktonic cells were discarded and the wells were washed twice with phosphate-buffered saline (PBS) (0.01 M, pH 7.2). The adherent cells were stained with 200 μL of 0.4% crystal violet solution for 5 min, and washed twice with distilled water. Subsequently, 200 μL of 20% glacial acetic acid was added to each well to dissolve the stained biofilm. After 30 min, the biofilm biomass was quantified by measuring the absorbance value at 570 nm. The inhibitory percentages of growth and biofilm formation were calculated according to the following formulas, respectively (Sivaranjani et al., 2016).
Assessment of metabolic activity of S. aureus cells in biofilm
The metabolic activity of S. aureus biofilm cells was evaluated by 2,3-bis (2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide sodium salt (XTT) reduction assay (Luís et al., 2014). After incubation and washing procedures, 120 μL of a combined solution of 100 μg/mL of XTT (Sigma Aldrich Co.) and 10 μg/mL of phenazine methosulfate (Sigma Aldrich Co.) prepared in PBS was added to each well. The plate was then incubated in the dark at 37°C for 3 h. The metabolic activity of biofilm cells was assessed by measuring the optical density at 492 nm with the microplate reader referred previously.
Observation of S. aureus biofilms
Light microscopic observation
The effect of DMY on the biofilm-forming ability of S. aureus was evaluated microscopically (Sivaranjani et al., 2016). In brief, 500 μL of logarithmic phase S. aureus suspension (1 × 106 CFU/mL) and 500 μL of different concentrations of DMY were added to each well of a 12-well microplate containing a round coverslip. TSB supplemented with 1% glucose containing DMSO without DMY was served as the negative control. After incubation at 37°C for 24 h, the coverslips were taken out, washed thrice with PBS, and stained with 0.4% crystal violet solution for 5 min. The coverslips were then washed with sterilized water and air dried. The stained biofilms were observed with a light microscope (Nikon Eclipse E200; Japan) at 400× magnification.
Scanning electron microscopic analysis
The effect of DMY on the architecture of S. aureus biofilm was further validated with a scanning electron microscope (SEM) (Luís et al., 2014). In brief, S. aureus biofilms were developed on the round coverslips in the presence and absence of DMY as already described. After removal of nonadherent cells, the coverslips were submerged with 2.5% glutaraldehyde at 4°C for 3 h. Thereafter, the coverslips were washed with PBS, dehydrated in a graded ethanol series, and dried at the critical point using a critical point dryer (Tousimis Autosamdri-815, USA) for 60 min. The specimens were then sputter-coated with gold and examined using a SEM (JSM-7500F; JEOL, Japan).
Stainless steel model assay
Stainless steel 304 coupons (2 cm × 2 cm × 1 mm thickness) were washed, autoclaved, and dried. The sterile coupons were immersed in the bacterial suspensions of logarithmic phase S. aureus (1 × 108 CFU/mL) at 37°C for 2 h. After incubation, the coupons were rinsed with 0.1% peptone water to remove planktonic cells, and then submerged in DMY solutions (dissolved in 10% ethanol) for 10 min. Each coupon was rinsed again with 0.1% peptone water to remove excess DMY solution. Subsequently, the washed coupon was immersed in 10 mL of 0.1% peptone water and sonicated at 55 kHz for 10 min. The sonicated samples were serially decimal diluted for cell count by plating onto nutrient agar plates and incubated at 37°C for 18–24 h. Two controls treated with 0.1% peptone water and 10% ethanol were conducted in parallel, respectively. The adhered bacterial density (CFU/cm2) was calculated using the following formula (Engel et al., 2017).
Statistical analysis
All the experiments were conducted in triplicate. Statistical analysis was performed on SPSS 19.0 software (IBM Co.). One-way analysis of variance was performed to determine significant differences (p < 0.05) with least significant difference or Games–Howell multiple comparisons according to the results of homogeneity test.
Results
MIC and MBC
DMY exhibited potent antibacterial and bactericidal activities against S. aureus ATCC 29213. The MIC and MBC values of DMY were detected to be 0.125 and 0.25 mg/mL, respectively.
Effect of DMY on the growth and biofilm formation of S. aureus
As shown in Figure 1, treatment with DMY at 0.25, 0.5, and 1 mg/mL slightly affected the planktonic growth of S. aureus, causing a minor (<10%) and no significant (p > 0.05) decrease compared with the control, whereas 2 mg/mL of DMY significantly (p < 0.01) inhibited the growth of S. aureus with inhibition of 27%. In contrast, all tested concentrations of DMY significantly (p < 0.01) reduced the biofilm formation of S. aureus. Moreover, an increased biofilm biomass inhibition was observed along with the increasing concentration of DMY, ranging from 54% to 89%.
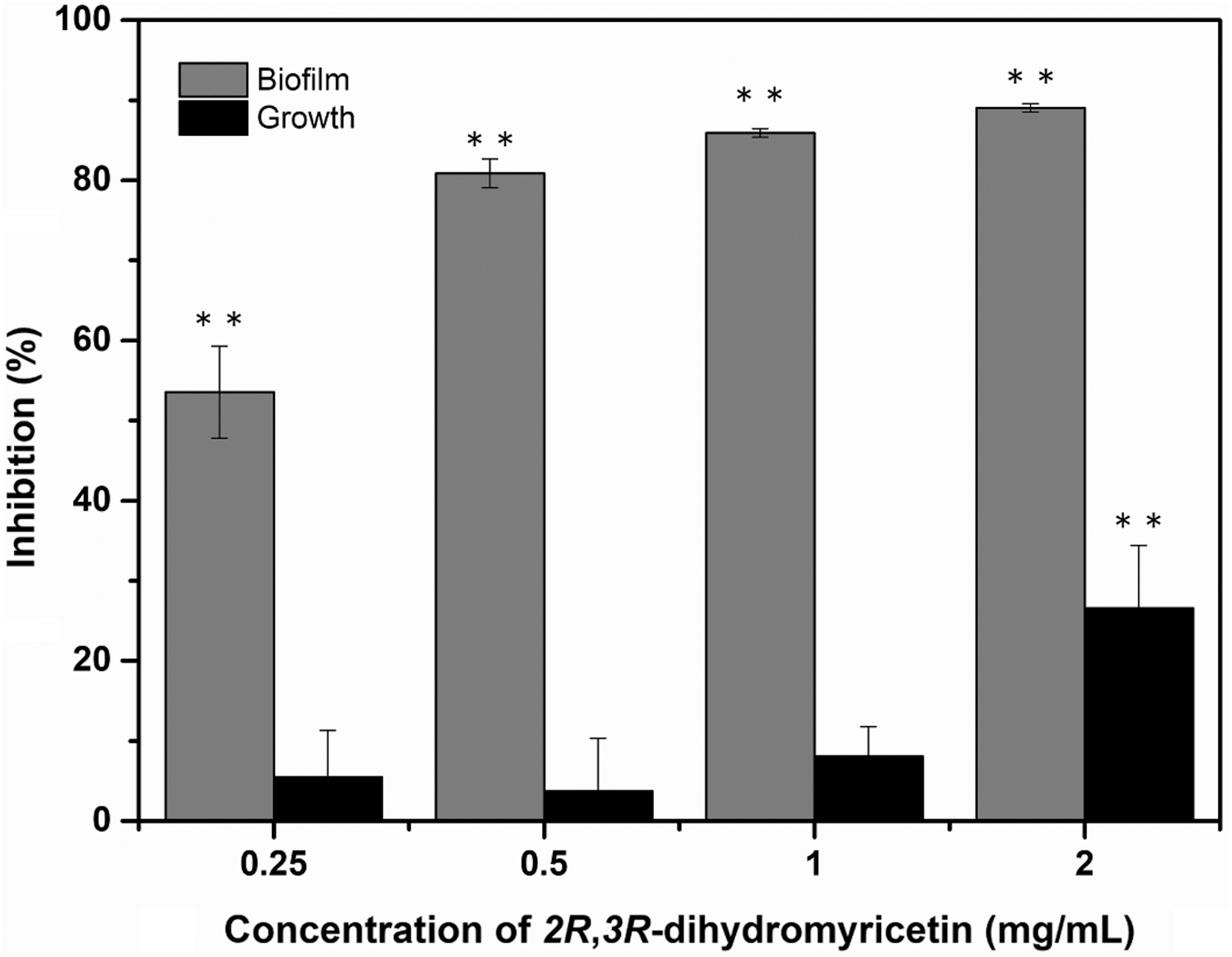
Effect of DMY on the planktonic growth and biofilm biomass of Staphylococcus aureus ATCC 29213. Bars represent standard deviation (n = 3). **p < 0.01. DMY, 2R,3R-dihydromyricetin.
Effect of DMY on the metabolic activity of S. aureus biofilm cells
Bacterial cells with metabolic activity are able to reduce XTT to a water-soluble orange formazan, which is easily quantified at 492 nm by a spectrophotometer. The absorbance value implies the total cellular viability of bacteria. As shown in Figure 2, the absorbance values at 492 nm of S. aureus cells treated with DMY at 0.25, 0.5, and 1 mg/mL were detected to be 0.23 ± 0.02, 0.14 ± 0.03, and 0.08 ± 0.01, respectively, whereas the control cells had an absorbance value of 0.27 ± 0.01. As a result, the absorbance value decreased significantly (p < 0.05) with increasing concentration of DMY from 0.25 to 1 mg/mL, which indicated that DMY caused a significant (p < 0.05) reduction in the metabolic activity of S. aureus biofilm cells.
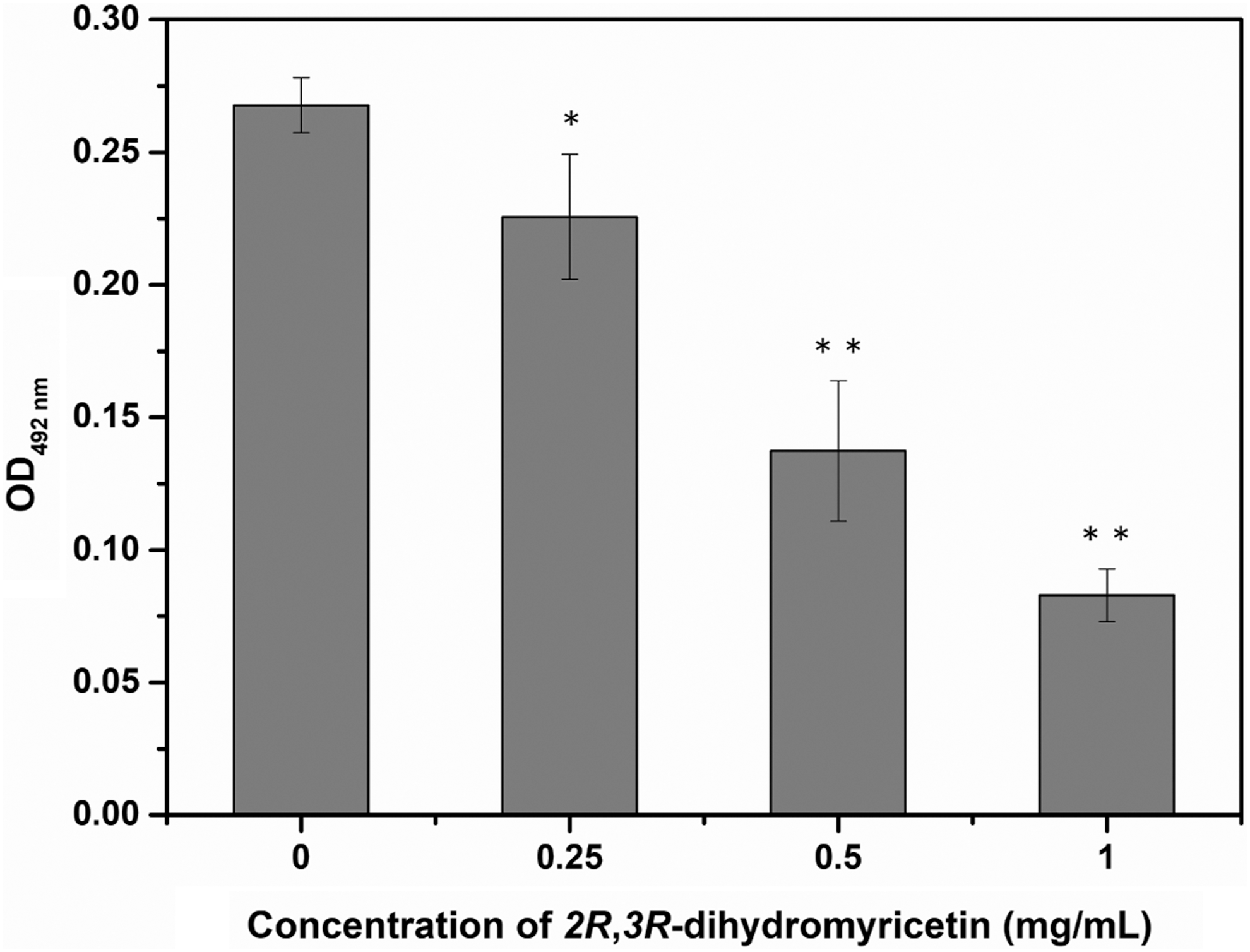
Effect of DMY on the metabolic activity of Staphylococcus aureus ATCC 29213 biofilm. Bars represent standard deviation (n = 3). *p < 0.05, **p < 0.01. DMY, 2R,3R-dihydromyricetin.
Effect of DMY on the morphology of S. aureus biofilm
The micrographs captured after crystal violet staining displayed the overall morphology of S. aureus biofilm, which showed that DMY disrupted the structure of bacterial biofilm and reduced the cell density in a dose-dependent manner (Fig. 3). Moreover, deep insights into the modifications in the biofilm architecture of S. aureus were further examined at higher magnification with SEM. In the control group, numerous S. aureus cells were embedded in an extracellular polymeric matrix and presented multilayered cell clusters. In contrast, the number of adherent bacterial cells was significantly reduced and dissolution of polymeric matrix was also observed after treatment with DMY. Scant bacterial biofilm composed of a few S. aureus cells was formed as small cell clusters or short chains.

Light microscope images
Effect of DMY on the S. aureus cells adhered to stainless steel
The initial count of the adherent cells in S. aureus biofilm on stainless steel was 5.08 log CFU/cm2, which was significantly (p < 0.01) reduced to 2.95, 1.22, and 0.37 log CFU/cm2 after treatment with 2, 4, and 8 mg/mL of DMY, respectively (Fig. 4). The reductions of S. aureus cells in biofilm were 2.13, 3.86, and 4.71 log CFU/cm2 when treated with DMY at 2, 4, and 8 mg/mL, respectively. In addition, no significant difference (p > 0.05) in the population of S. aureus biofilm cells was observed between the 10% ethanol-treated and control group.
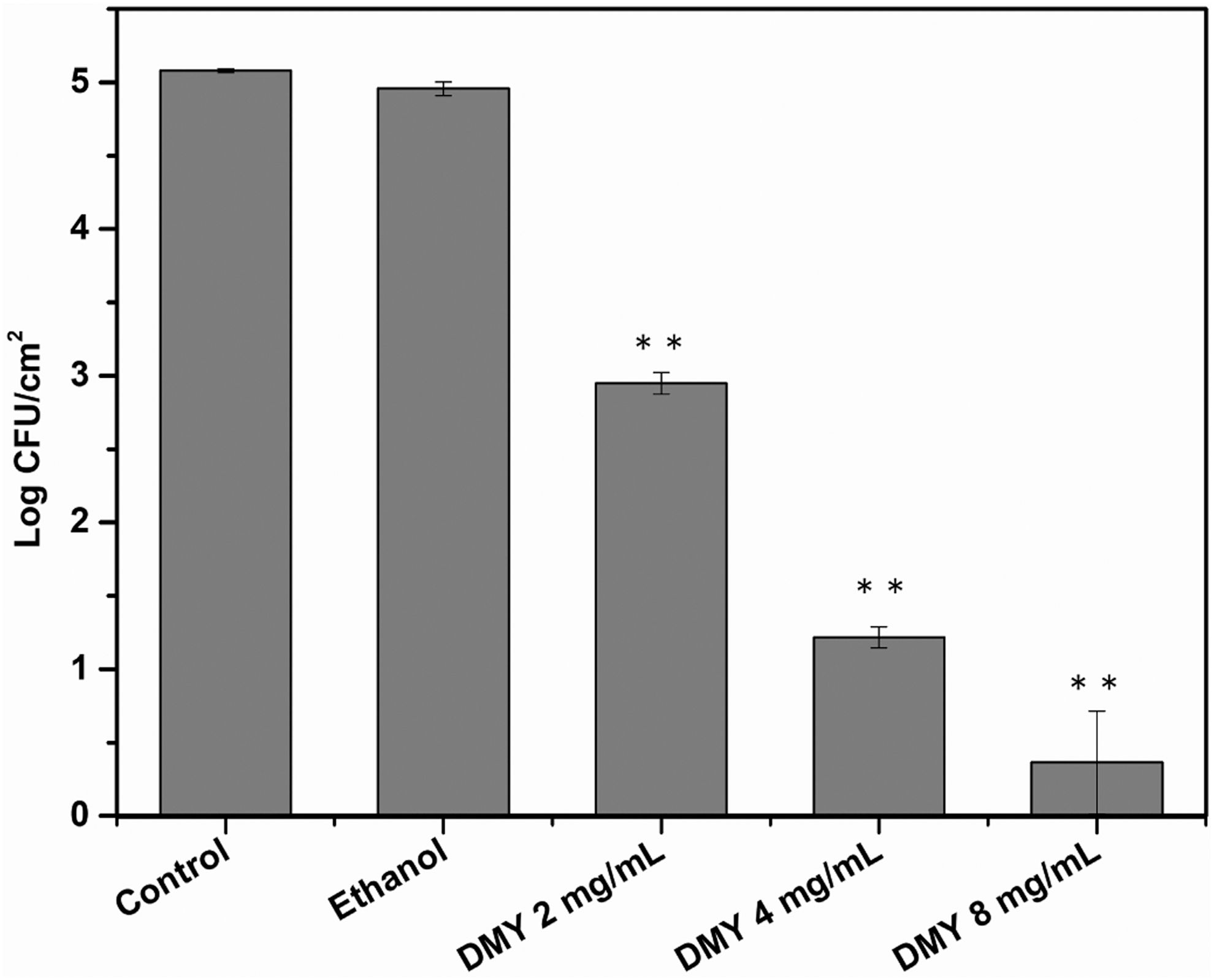
Effect of DMY on the population of Staphylococcus aureus ATCC 29213 biofilm cells adhered to stainless steel. Bars represent standard deviation (n = 3). **p < 0.01. DMY, 2R,3R-dihydromyricetin.
Discussion
Biofilm formation by undesirable pathogens on food contact surfaces is common because the presence of nutrients combined with appropriate environmental conditions facilitates the growth and attachment of bacteria to surfaces. Most of the microbial contamination of food products is biofilm related (Brooks and Flint, 2008), which causes significant problems and still remains challenging for the food industry. Thus, it is imperative to search for new alternative strategies, and polyphenols with antimicrobial activity have attracted much attention due to their abundance in plants, benefits for human health, and potential inhibitory effect on biofilm formation (Slobodníková et al., 2016). This study investigated the antibacterial and antibiofilm activities of DMY, a flavonoid found in C. deodara, against S. aureus and mainly focused on evaluating the effect of DMY on biofilm formation.
The results of MIC determination revealed that DMY completely inhibited the visible bacterial growth of S. aureus ATCC 29213 at 0.125 mg/mL. However, DMY showed lower inhibitory activity in the biofilm assay with inhibition of 27% even at 2 mg/mL (Fig. 1). Biofilm cells are known to be more resistant to antimicrobial agents than the planktonic counterparts with 10–1000 times less susceptible (Davies, 2003). The protective effect is multifactorial and principally conferred by the decreased penetration of antimicrobials, alteration of physiological state of biofilm cells, presence of persistent subpopulations, expression of specific resistance genes, and upregulation of efflux pumps (Høiby et al., 2010; Borges et al., 2017). In contrast, the biofilm biomass measurement demonstrated that DMY significantly inhibited the biofilm formation of S. aureus at concentrations ranging from 0.25 to 1 mg/mL, although the planktonic growth was slightly inhibited (Fig. 1). It is implied that the reduction in biofilm biomass was predominantly attributed to the antibiofilm activity of DMY rather than the decreased total number of S. aureus cells due to the antibacterial effect.
Furthermore, the results of XTT reduction assay showed that DMY markedly decreased the cellular viability of S. aureus biofilm in a dose-dependent manner, which confirmed the antibiofilm activity of DMY obtained from the biofilm biomass quantification. Our findings are consistent with a previous study that demonstrated that gallic, caffeic, and chlorogenic acids showed strong inhibitory effect on the metabolic activity of adherent cells in S. aureus biofilm (Luís et al., 2014). It was recognized that such decrease in metabolic activity as a physiological change was responsible for the resistance of biofilms to antimicrobial agents (Bazargani and Rohloff, 2016). Numerous studies have revealed that most of the antibiofilm agents inhibited the biofilm formation of microorganisms coupled with decrease in the cellular viability of biofilm (Luís et al., 2014; Sivaranjani et al., 2016; Tan et al., 2016; Khan et al., 2017). Interestingly, a recent study suggested that inversely proportional correlations were present between the biomass and metabolic inhibition of chitosan against S. aureus biofilm in some cases (Costa et al., 2017). As a consequence, metabolic activity probably indirectly interfered with the ability of bacteria to adhere and form biofilms.
Formation of characteristic biofilm architecture is a crucial stage in the biofilm development (You et al., 2007). The micrographs of light microscope and SEM further validated the remarkable inhibitory effect of DMY on the biofilm formation by S. aureus, which revealed that DMY caused a huge collapse on the complex biofilm architecture, resulting in loosely arranged bacterial cell that can be easily disrupted. Similar observations have also been reported for many antibiofilm compounds that exhibited disruption effect on the recalcitrant three dimensional structure of S. aureus biofilm (Luís et al., 2014; Martínez et al., 2016; Kannappan et al., 2017). The disintegration efficacy of DMY on S. aureus biofilm could lead to a decrease in the resistance of sessile bacteria to antibacterial agents.
Considering the promising antibiofilm activity of DMY against S. aureus biofilm formed in food processing, an attempt was made to assess the prevention of S. aureus biofilm formation on stainless steel. Stainless steel is extensively used for the manufacture of equipment in food industry because of its excellent physicochemical properties and relative low cost. However, the hydrophilicity of stainless steel provides favorable conditions for the attachment and biofilm formation of bacteria (Engel et al., 2017). The results indicated that S. aureus were able to adhere in high numbers to stainless steel after a contact of 2 h, which was distinctly reduced by a short time treatment with DMY. However, higher concentrations of DMY were required to achieve comparable inhibitory efficacy on the biofilm formation compared with that used in the biofilm biomass measurement. One possible reason is that adherent cells are generally more resistant to antimicrobial agents, and removal of adherent cells is more difficult than prevention of bacterial adhesion. Consequently, preventing bacterial adhesion to food contact surfaces is vital for the control of biofilm formation by pathogens in a food processing environment.
Conclusions
This study revealed that DMY exhibited antibacterial and antibiofilm activities against S. aureus. Significant reduction in the biofilm biomass and metabolic activity demonstrated the inhibitory efficacy of DMY against the biofilm formation by S. aureus. Light microscopic observations and SEM analysis further validated that DMY caused a huge collapse on the complex biofilm architecture, leading to a decrease in the biofilm formation. Furthermore, DMY reduced the population of sessile S. aureus cells adhered to stainless steel. Hence, these findings indicated that DMY might be a promising candidate as a natural antibiofilm agent in the food industry.
Footnotes
Acknowledgment
This work was financially supported by the National Natural Science Foundation of China (No. 31571936).
Disclosure Statement
No competing financial interests exist.